
Magnesium in skin allergy
Joanna Błach 1 , Wojciech Nowacki 1 , Andrzej Mazur 1
Streszczenie
Magnez pełni wiele funkcji prawie we wszystkich procesach zachodzących w ustroju. Jego niedobór powoduje wiele zaburzeń w organizmie, m.in. upośledza odporność immunologiczną. Przedstawiona praca jest przeglądem literatury dotyczącym związku między magnezem i reakcjami alergicznymi skóry, ze szczególnym uwzględnieniem reakcji nadwrażliwości typu I i IV. Najlepiej znane są zależności między magnezem i nadwrażliwością typu I. Deficyt magnezu u zwierząt doświadczalnych, głównie u szczurów, ujawnia się charakterystycznym rozszerzeniem naczyń obwodowych, wzrostem stężeń swoistych dla alergii immunoglobulin klasy IgE, wzrostem liczby neutrofilów i eozynofilów, podniesieniem stężeń cytokin prozapalnych, degranulacją komórek tucznych i histaminemią, oraz splenomegalią. Istnieje podobieństwo w objawach klinicznych obserwowanych u szczurów z hipomagnezemią a pacjentami z atopią skórną. Znikome są informacje o wpływie magnezu na pozostałe typy reakcji nadwrażliwości, z wyjątkiem fragmentarycznych danych na temat związku magnezu z nadwrażliwością typu IV Obserwacje kliniczne wskazują na korzystny wpływ zastosowania magnezu zarówno stosowanego miejscowo, jak i doustnego na przebieg chorób alergicznych skóry. Wszystkie dane literaturowe wskazują na ważność związku między magnezem i reakcjami nadwrażliwości. Potrzebne są dalsze badania zarówno nad mechanizmami działania magnezu jak i kontrolowane badania kliniczne nad skutecznością suplementacji w magnez u pacjentów z alergią skóry.
Słowa kluczowe:magnez • deficyt magnezu • alergia • nadwrażliwość • reakcja zapalna • skóra
Summary
Magnesium is involved in many biological processes within the body. Magnesium deficiency causes many disorders, including impairment of immunity. This review summarizes present knowledge on the relationship between magnesium and skin allergy reactions. Special focus is on allergy types I and IV. At present the best knowledge is on allergy I. Magnesium deficiency in experimental animals, mainly rats, leads to characteristic hyperemia, an increase in IgE, neutrophilia and eosinophilia, an increase in the level of proinflammatory cytokines, mastocyte degranulation, histaminemia, and splenomegaly. These symptoms observed in hypomagnesemic rats are similar to those in atopic patients. Data on the relationship between magnesium and other types of allergy are scarce. Clinical observations show the beneficial effect of topical and oral administration of magnesium salts in patients with skin allergy. All the presented data point to an important role of magnesium in allergy reactions. Other studies are needed to better understand the mechanism of magnesium’s action. Well-controlled clinical protocols should also be conducted to assess the efficiency of magnesium supplementation in patients with skin allergy.
Key words:magnesium • magnesium deficiency • allergy • hypersensitivity • inflammation • skin
WSTĘP
Magnez jest drugim – po potasie – kationem wewnątrzkomórkowym występującym u ssaków, pod względem ogólnej ilości. Głównym jego rezerwuarem w organizmie są kości i mięśnie [15]. Pełni on wiele istotnych funkcji niemal we wszystkich procesach zachodzących w ustroju. Magnez uczestniczy w wielu szlakach metabolicznych zachodzących w komórce. Jest on aktywatorem prawie 300 różnych enzymów, przez co uczestniczy w przemianie węglowodanów, kwasów nukleinowych oraz białek [13,57]. Stabilizuje strukturę DNA, wpływa na transkrypcje RNA, składanie jednostek rybosomalnych [73]. We wszystkich procesach przebiegających z udziałem ATP wymagana jest obecność jonów Mg2+ [58]. Poprzez aktywny kompleks Mg2+-ATP bierze udział w reakcjach fosforylacji [16,73]. Jedną z najważniejszych właściwości jest jego rola stabilizująca błony komórkowe. Wiązanie magnezu w związki kompleksowe z fosfolipidami zmniejsza ich płynność i przepuszczalność [13]. Magnez jest również konieczny do zainicjowania kaskady dopełniacza zarówno w sposób klasyczny jak i alternatywny [40].
Ważne jest utrzymanie homeostazy magnezu zarówno w przestrzeniach komórkowych, jak i pozakomórkowych organizmu. Za utrzymanie stałego poziomu magnezu w organizmie są odpowiedzialne głównie nerki, w których zachodzi odzyskiwanie (reabsorpcja) i jelita, gdzie odbywa się wchłanianie (absorpcja) [58]. Za utrzymanie homeostazy Mg2+ na poziomie komórkowym odpowiedzialne są różnego rodzaju systemy transportu Mg2+ m.in. antyporty Na+/Mg2+ i kanały TRPM6 i TRPM7 (transient receptor potential melastin) [42,50,62,68]. Pierwotny niedobór magnezu wynika z niedostatecznej jego podaży w diecie. U ludzi ma zwykle przebieg przewlekły lecz łagodny. Niedobór wtórny magnezu jest zwykle spowodowany zaburzeniami jego wchłaniania lub wydalania, np. niewydolność nerek, a także przez stosowane terapie (np. diuretyki) czy stres [13]. Najlepiej znane i najwcześniej rozpoznawane spośród pierwotnych niedoborów magnezu są postacie nerwowo-mięśniowe. Nawet niewielki niedobór Mg2+ w ustroju może spowodować wzmożoną pobudliwość nerwowo-mięśniową, stany rozdrażnienia, lęku, bezsenność, zmęczenie, bóle i zawroty głowy, kołatanie serca, drgania powiek, drętwienie kończyn, w miarę pogłębiania się jego niedoboru rozwija się obraz tężyczki. Przykładem klinicznego niedoboru magnezu u zwierząt jest tężyczka pastwiskowa (Hypomagnesemic tetany), występująca u bydła z zaburzoną gospodarka mineralną (niedobór magnezu, nadmiar potasu w paszy) i objawiająca się napadami drgawek [64]. Niedobór magnezu wpływa więc na upośledzenie wielu elementów odporności [39] i może sprzyjać występowaniu alergii typu I wg klasyfikacji Gella i Coombsa. Wywołany eksperymentalnie deficyt Mg2+ u szczurów powoduje utratę masy ciała [9,41] i splenomegalię [4,7,10,46]. Powiększenie śledziony jest spowodowane naciekiem przez leukocyty wielojądrzaste (PMN) i makrofagi [35]. Niedobór tego pierwiastka przyczynia się także do przyspieszonej inwolucji grasicy [10,37]. Zwiększa się ilość komórek ulegających apoptozie, następuje zubożenie istoty korowej w tymocyty już w trzecim tygodniu obserwacji, a w kolejnych tygodniach prawie całkowity ich zanik [65]. W niedoborze magnezu dochodzi do obniżenia stężenia immunoglobulin, spada stężenie IgM do 70%, IgG 35–60%, IgA 25–30% [39], wzrasta natomiast stężenie IgE [10,39], rozwija się rekcja ostrej fazy.
Magnez może odgrywać rolę w przebiegu reakcji alergicznych. Dla przypomnienia wyróżniamy 4 typy reakcji nadwrażliwości (podsumowanie w tabeli 1)
Tabela 1. Cztery typy reakcji nadwrażliwości
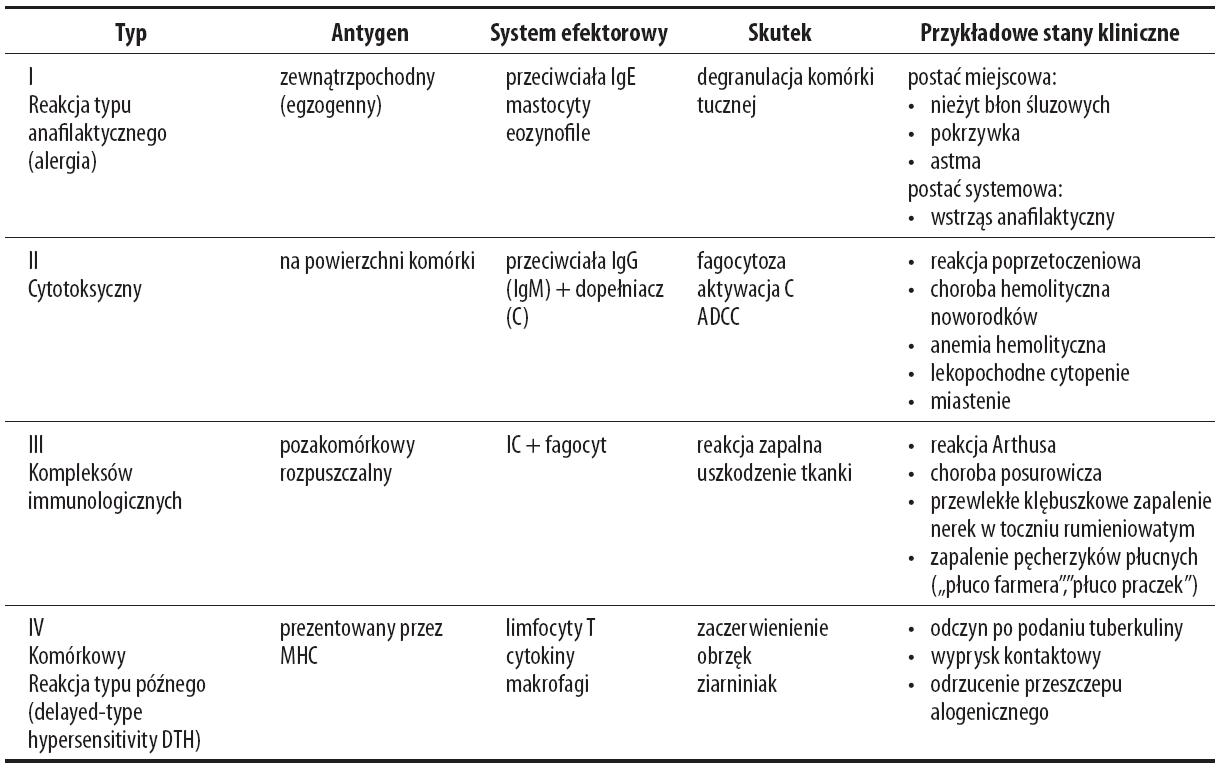
Celem tego artykułu jest przedstawienie wpływu magnezu, głównie jego niedoboru, na przebieg reakcji nadwrażliwości objawiających się na skórze, głównie typu I i IV, gdyż tylko w tych przypadkach znane są zależności z jonami Mg2+.
MAGNEZ I ALERGIA
Mg2+ i I typ alergii
Wiadomo, że w tym typie nadwrażliwości magnez ma wpływ na występowanie zmian skórnych i na astmę [2,13,17,24,30,59,67].
Łatwo wykazać niedobory magnezu na modelach doświadczalnych, które można wywołać przez karmienie dietą niedoborową [14,25,60]. Pierwszym klinicznym objawem u szczurów jest charakterystyczne rozszerzenie naczyń obwodowych, zauważalne na uszach [7,27,29,36,37,41] następnie zaczerwienienie obejmuje podstawę ogona, a później głowę, tułów i łapy [27,45]. Zmiany skórne utrzymują się przez 4–5 dni, następnie słabną, ale nie zanikają całkowicie [27,60]. U myszy nie zauważono takich symptomów [41]. Doskonałym modelem zwierzęcym w badaniach nad zapaleniami skóry są nagie szczury, u których łatwiej zauważyć zaczerwienienia [25] i u których widoczne symptomy pojawiają się szybciej, niż u szczurów owłosionych. Ogólnie u szczurów z niedoborem magnezu, oprócz widocznych na skórze zmian obejmujących zaczerwienienie i dermatozy, odnotowano również inne objawy występujące także przy alergii. Należą do nich wzrost liczby neutrofilów i eozynofilów, histaminemia oraz splenomegalia [10].
Ponieważ podczas niedoboru magnezu dochodzi do wzrostu poziomu IgE w surowicy potwierdza to założenie, że jego deficyt łączony jest z reakcją nadwrażliwości typu I, w której biorą udział immunoglobuliny tej klasy. Wiążą się one do receptorów znajdujących się na komórkach tucznych i bazofilach zwiększając gotowość komórek do degranulacji [10,40,41]. Uwolnione zostają aktywne mediatory obejmujące histaminę, serotoninę, ECF-A (czynnik chemotaktyczny eozynofilów), heparynę, leukotrieny (LTB4, LTC4, LTD4), tromboksany, prostaglandyny, PAF (czynnik aktywujący płytki krwi) [40]. Uwolniona histamina łączy się z odpowiednimi receptorami komórek określonych tkanek, powodując rozszerzenie naczyń i zwiększenie ich przepuszczalności, czego klinicznym objawem jest miejscowe przekrwienie i obrzęk spowodowany przenikaniem osocza do przestrzeni międzykomórkowej. U myszy i szczurów z deficytem magnezu odnotowano zwiększony poziom histaminy w tkankach grasicy, śledziony, nerek, płuc, w dnie żołądka i mięśniach szkieletowych [51], przy czym większy wzrost stwierdzono u szczurów [41]. Do wzrostu stężenia histaminy w różnych tkankach przyczynia się wzrost aktywności dekarboksylazy histydyny a spadek aktywności oksydazy tiaminy [48], jednak w skórze szczurów z obniżonym stężeniem magnezu, pomimo podwyższenia aktywności dekarboksylazy histydyny, zawartość histaminy nie wzrasta [46,47].
Jony magnezu biorą udział w aktywacji cyklazy adenylowej, syntetyzującej powstanie cyklicznego adenozynomonofosforanu (cAMP). Ponieważ duża wewnątrzkomórkowa koncentracja cAMP i cGMP (cyklicznego monofosforanu guanozyny) pomaga spowolnić lub zatrzymać degranulację komórek, aktywacja cyklazy adenylowej jest decydująca w kontroli reakcji anafilaktycznych. Dostępność magnezu dla enzymu może więc zmieniać metabolizm cyklicznego nukleotydu w komórce i dlatego sądzi się, że Mg2+ łagodzi reakcje nadwrażliwości [40]. Deficyt magnezu natomiast może stymulować uwolnienie histaminy z komórek tucznych, przez zahamowanie wytwarzania zależnego od Mg2+ cAMP [14].
Kolejnym charakterystycznym dla alergii, i ostrej fazy, objawem jest wzrost ilości leukocytów [19,28,39,44], zauważalne są również zmiany w ich subpopulacjach [28,37]. Znacznie wzrasta liczba krążących wielojądrzastych komórek (PMN), co prowadzi do neutrofilii [4,36], która sprzyja fagocytozie, wytwarzaniu reaktywnych form tlenu i cytokin [8]. Eozynofilia, która pojawia się bardzo wyraźnie od 4 tygodnia doświadczenia, jest zwykle reakcją na wytwarzane przez komórki tuczne substancje chemotaktyczne [28]. Procesom tym towarzyszy toczący się proces zapalny, objawiający się m.in. podwyższeniem stężenia cytokin prozapalnych (IL-1, IL-6, TNF) po 3 tygodniach magnezowo zubożonej diety [72]. Badania Rayssiguiera i wsp. [55] wykazują, że podniesienie poziomu krążącej IL-6 następuje już od 4 dnia zastosowania tej diety, ale nie towarzyszył temu wzrost TNF. IL-6 jest wytwarzana przez wiele komórek, obejmujących makrofagi, monocyty i komórki śródbłonka. Ten mediator zapalenia stymuluje syntezę białek ostrej fazy w wątrobie [1]. W osoczu stwierdza się zmiany stężeń pozytywnych i negatywnych białek ostrej fazy (BOF) [54]. Po tygodniu diety deficytowej odnotowano wzrost poziomu a
2-makroglobuliny, a
1-kwaśnej glikoproteiny i fibrynogenu, oraz znaczny spadek poziomu BOF negatywnych – albuminy, RBP (retinol-binding protein) i apolipoproteiny E w osoczu [36]. W osoczu szczurów z deficytem Mg2+ wzrasta poziom składnika C3 dopełniacza. Wzrost ten jest spowodowany toczącą się reakcją zapalną, ponieważ synteza C3 dopełniacza jest indukowana w odpowiedzi na cytokiny prozapalne (IL-1, IL-6 i TNF) [18]. U zwierząt z niedoborem podniesienie poziomu dopełniacza może być odpowiedzialne za wczesną degranulację komórek tucznych i aktywację fagocytów. Mimo podwyższenia poziomu składowej C3 dopełniacza w osoczu u szczurów niedoborowych nie dochodzi u tych zwierząt do rozdziału tego składnika (metodą Western-blotting) [7]. Wyniki te poddają w wątpliwość rolę dopełniacza i jego aktywacji w reakcji zapalnej spowodowanej niedoborem magnezu. We krwi szczurów z obniżonym stężeniem Mg2+ stwierdza się także wzrost poziomu trójglicerydów i spadek poziomu lipoprotein o dużej gęstości (HDL), co jest również charakterystyczne dla ostrej fazy [38,43].
Podstawy reakcji zapalnej w deficycie magnezu są ciągle niepewne. Markerem, który pojawia się najszybciej jest substancja P [70,71,72]. Neuropeptyd ten wpływa na uwalnianie cytokin przez limfocyty T, inicjuje wytwarzanie wolnych rodników i powoduje uwolnienie histaminy z komórek tucznych [70,72]. Przypuszcza się więc, że to ona inicjuje proces zapalny wywołany deficytem magnezu. Jednak badania Malpuech-Brugčre i wsp. [36] wykazują, że wysoki poziom IL-6 wykryto bez znacznego wzrostu poziomu substancji P.
Deficyt magnezu powoduje wzrost wytwarzania wolnych rodników. Na szczególną uwagę zasługuje również tlenek azotu (NO), ponieważ bierze udział w funkcjach fizjologicznych, takich jak regulacja napięcia naczyń czy cytotoksyczność aktywowanych makrofagów [56]. Jest on syntetyzowany z pochodnej L-argininy, przez trzy różne syntetazy (nitric oxide synthase NOS) [3]. Przypuszcza się, że iNOS odgrywa decydującą rolę przy wzroście poziomu NO w osoczu podczas deficytu Mg2+. Koncentracja tlenku azotu w osoczu wzrasta od drugiego dnia deficytu, a najwyższe wartości osiąga dnia ósmego, w porównaniu ze szczurami z grupy kontrolnej [4,56]. Stwierdzono również, że spadek koncentracji Mg2+ w pożywce hodowlanej powoduje uwolnienie NO z makrofagów już po 2 godzinach [74]. Wyższy poziom NO może być spowodowany jego wzmożonym wytwarzaniem przez zaktywowane komórki podczas odpowiedzi zapalnej i może się przyczyniać do wzrostu stresu oksydacyjnego i uszkodzenia oksydacyjnego tkanek [32,33,56], czego dowodem może być spadek poziomu glutationu (GSH) w czerwonych krwinkach szczurów po 2–3 tygodniach diety ubogiej w magnez [70].
Istnieją różnice między wpływem deficytu magnezu a wiekiem, płcią zwierząt i składnikami diety. Bardziej wrażliwe na deficyt magnezu są samce w okresie wzrostu. U młodych szczurów już niewielki deficyt magnezu w pożywieniu powoduje po 4 dniach wystąpienie wysypki, podczas, gdy u szczurów starszych (6 miesięcznych), była ona obserwowana dopiero przy przedłużonym i znacznym deficycie. Ta odmienność w odpowiedzi jest spowodowana różnicami w zasobach tkankowych magnezu (głównie w kościach i mięśniach) między zwierzętami młodymi i dorosłymi. Ogólnie zmiany pojawiają się wcześniej u samców [60]. Wystąpienie przekrwienia na uszach u dorosłych szczurów z deficytem magnezu jest zależne częściowo od hormonów płciowych, głównie estradiolu wpływającego na metabolizm histaminy [49]. U samic stwierdzono również mniej zaznaczoną leukocytozę z neutrofilią i niższy poziom trójglicerydów w osoczu, w porównaniu z samcami. Samce szczurów karmione paszą bez Mg2+, którym podano estrogen (100 µg/100 g masy ciała/dzień), były chronione przed następstwami niedostatku Mg2+, odnotowano bowiem u nich mniejszą liczba leukocytów i trójglicerydów, w porównaniu z grupą, której nie podano hormonu [5]. W związku z tym, że fosfor w diecie jest odpowiedzialny za spadek absorpcji magnezu, przypuszczano, że wysoki poziom tego pierwiastka może nasilać objawy niedoboru magnezu [69]. Badania tych autorów dowodzą, że u młodych szczurów dieta z dużą zawartością fosforu nasila reakcję zapalną wywołaną ostrym deficytem magnezu, ujawniającą się powiększeniem śledziony i leukocytozą. Tymczasem badania tych samych autorów pokazują, że również niski poziom fosforu nasila tę reakcję zapalną i to w większym stopniu niż jego nadmiar. Mechanizm odpowiedzialny za interakcje między niskim poziomem magnezu i fosforu w diecie jest nieznany. Natomiast deficyt wapnia w diecie wpływa hamująco na przebieg reakcji zapalnej wywołanej niedoborem Mg2+ [6]. Mechanizm tej interakcji zostanie omówiony poniżej.
Z innych interakcji pokarmowych znany jest nasilający wpływ diety wysokosacharozowej lub wysokofruktozowej na objawy zapalenia u zwierząt z niedoborem magnezu [53]. Prawdopodobnie interakcja może być wytłumaczona zwiększonym wytwarzaniem wolnych rodników poprzez dietę wysokocukrową.
Karmiąc nagie myszy HR-1 paszą ze zredukowaną zawartością magnezu i cynku otrzymano mysi model, z charakterystycznymi objawami dla atopowego zapalenia skóry u ludzi. Po kilku tygodniach od rozpoczęcia eksperymentu u myszy obserwowano pomarszczenia, suchość i znaczny spadek ilości wody w skórze. Badania immunohistologiczne wykazują znaczny wzrost liczby komórek T CD4+, IgE, IL-4, IL-5 w skórze w porównaniu z myszami karmionymi standardową dietą. Wysypka, która towarzyszy deficytom Zn2+ i Mg2+, jest hamowana przez leki antyhistaminowe. Jednak mysi model zapalenia skóry jest swoisty tylko dla myszy szczepu HR-1, nie uzyskano symptomów atopowego zapalenia skóry u myszy Balb/c, C57BL i ICR, które także karmiono dietą ze zredukowaną ilością Zn2+ i Mg2+ [34]. Istnieje podobieństwo w objawach klinicznych między szczurami z hipomagnezemią a pacjentami z atopią skórną [20]. Skóra szczurów z niedoborem magnezu może więc być modelem do oszacowania skuteczności środków przeciwko stanom zapalnym skóry.
Mg2+ i IV typ reakcji nadwrażliwości
Mało jest danych na temat zależności między magnezem a przebiegiem nadwrażliwości typu komórkowego (typ IV wg Coombsa i Gella). Ogólnie uważa się, ze deficyt magnezu ogranicza objawy tego typu nadwrażliwości [14]. Na przykład niedobór tego jonu hamuje nadwrażliwość kontaktową na dinitrochlorobenzen (DNCB). Stwierdzono bowiem, że w przeciwieństwie do grupy kontrolnej, szczury z niedostatkiem magnezu nie odpowiadały na DNCB. Tymczasem szczury, które poddano niedoborowi po uwrażliwieniu, odpowiadały pozytywnie na wywołującą dawkę DNCB [25]. Wskazuje to na decydujący wpływ statusu magnezowego w chwili uwrażliwienia na przebieg reakcji nadwrażliwości typu IV.
HIPOTEZA MECHANIZMÓW DZIAŁANIA
Magnez jest antagonistą wapnia, obniżenie zewnątrzkomórkowego poziomu Mg2+ powoduje wzrost wewnątrzkomórkowego Ca2+, natomiast wzrost zewnątrzkomórkowego Mg2+ daje odwrotny efekt. Wiadomo, że wzrost w osoczu szczurów koncentracji Ca2+ jest odpowiedzią na deficyt Mg2+. Zawartość wapnia w surowicy znacznie wzrasta u szczurów z małą zawartością magnezu w diecie, zauważalna jest odwrotna korelacja liniowa między ilością Mg2+ w diecie a zawartością Ca2+ w surowicy [75]. Napływ Ca2+ do komórki (po stymulacji antygenem) jest głównym czynnikiem do syntezy histaminy i uwolnienia IL-6 z komórek tucznych [66]. Deficyt Ca2+ zapewnia znaczną ochronę przeciwko prozapalnemu działaniu spowodowanemu przez deficyt Mg2+. U szczurów karmionych paszą z deficytem Mg2+, ale z odpowiednią ilością Ca2+ przekrwienie na uszach pojawiało się już od 4 dnia i utrzymywało się do końca eksperymentu. Natomiast w grupie szczurów karmionych paszą zarówno deficytową w Mg2+, jak i Ca2+ hipermia pojawiała się wolniej i była słabsza [6].
W skórze odpowiedni stosunek Ca2+/Mg2+ jest niezbędny do utrzymania lokalnej jonowej homeostazy [11,12]. Uszkodzenia skóry skutkują zmianami we wzajemnej pozakomórkowej koncentracji Mg2+ i Ca2+, dając pierwszeństwo magnezowi, który ułatwia i poprawia zdolności migracyjne komórek, takich jak fibroblasty, leukocyty, komórki nabłonka, które odgrywają rolę w udanym procesie leczenia ran [22,23].
WŁAŚCIWOŚCI LECZNICZE MG2+ W CHOROBACH SKÓRY
Liczne obserwacje wskazują na korzystne działanie soli magnezu w zapaleniach skóry u ludzi [52,61]. Badano skuteczność kąpieli u chorych z atopowym zapaleniem skóry, w wodzie z Morza Martwego, która jest szczególnie bogata w MgCl2. Kąpiele te znacznie poprawiały funkcję bariery skóry, a chropowatość (szorstkość) i zaczerwienienie, jako znaczniki zapalenia, były znacznie zredukowane [52]. Stwierdzono również, że jony Mg2+ hamują zdolność prezentowania antygenów przez obecne w skórze komórki Langerhansa, co może się przyczyniać do skuteczności kąpieli w wodach z Morza Martwego w leczeniu zapaleń skóry [61].
Istnieją doniesienia, że wyselekcjonowane z Morza Martwego sole bogate w Mg2+ mają zdolności hamowania proliferacji fibroblastów, co może się przyczyniać do poprawy kondycji skóry u chorych z łuszczycą. Badania Levi-Schaffer i wsp. [31] wskazują, że sole te in vitro hamują podziały ludzkich fibroblastów z łuszczycą, jak i fibroblastów od ludzi zdrowych, przy czym najbardziej czynne okazały się sole magnezu: MgCl2 i MgBr2, które w największym stopniu hamowały proliferację fibroblastów. U chorych na łuszczycę stwierdza się w naskórku znacznie niższe stężenie cAMP niż w skórze zdrowej, a po krótkotrwałym zastosowaniu leków powodujących wzrost wewnątrzkomórkowego stężenia tego nukleotydu, obserwowano znaczną poprawę na skórze. Ponieważ Mg2+ jest aktywatorem cAMP istnieją sugestie, że duże stężenie Mg2+ ma właściwości terapeutyczne w łuszczycy [31]. Stosowanie naskórne przez dwa tygodnie wybranych soli z Morza Martwego na skórę zdrowych królików, jest wystarczające do uzyskania wysokiego poziomu Mg2+ i K+ w ich osoczu [63]. Ponadto Mg2+ i K+ mają zdolność hamowania syntezy poliamin, które są związane z patogenezą łuszczycy.
Opisany jest również wpływ doustnej suplementacji magnezu na choroby skóry. Picie oczyszczonej głębinowej wody morskiej, która zawierała Mg2+ jako główny kation, poprawia symptomy skórne u pacjentów z zespołem atopowego wyprysku skóry/zapalenia skóry (atopic egzema/dermatitis syndrome AEDS) [26]. U chorych, którzy przez rok pili dziennie 500 ml wody morskiej, odnotowano poprawienie objawów skórnych, podczas gdy w grupie pacjentów, którzy pili wodę destylowaną polepszenia nie stwierdzono. Również w surowicy chorych otrzymujących wodę z dużą zawartością Mg2+, spada poziom IgE i cytokin: IL-4, IL-13 i IL-18, czego nie zaobserwowano w grupie kontrolnej. Dane te wskazują, że woda morska zawierająca duże ilości Mg2+ wpływa korzystnie na skórę chorych z AEDS.
Istnieją także dane na temat korzystnego wpływu Mg2+ w alergicznym kontaktowym zapaleniu skóry, wywoływanym przez 1-chloro-2,4-dinitrobenzol. Reakcję wywoływano u myszy (Balb/c), stosując ten związek powierzchniowo na ucho w obecności chlorku magnezu (28 i 14%). Stwierdzono, że opuchlizna na uchu w tym przypadku była znacznie mniejsza, niż jeśli zastosowano go samodzielnie lub w połączeniu z chlorkiem sodu. Znajduje to potwierdzenie u ludzi uczulonych na nikiel, u których chlorek magnezu hamował kontaktowe zapalenie skóry powodowane siarczanem niklu, podczas gdy chlorek sodu nie zmieniał przebiegu reakcji [21].
PODSUMOWANIE
Podsumowując przedstawiony powyżej przegląd literatury można stwierdzić, że dieta uboga w magnez i hipomagnezemia nasilają reakcję nadwrażliwości typu I. Znikome są natomiast dane na temat wpływu tego pierwiastka na inne typy reakcji nadwrażliwości. Piśmiennictwo wskazuje na korzystny wpływ zastosowania magnezu zarówno lokalnego, jak i doustnego na przebieg chorób alergicznych skóry.
Potrzebne są dalsze badania nad mechanizmami zależności między magnezem a reakcjami nadwrażliwości oraz kontrolowane badania kliniczne nad skutecznością suplementacji w magnez u pacjentów z alergią skóry.
PIŚMIENNICTWO
[1] Bauman H., Gauldie J.: Regulation of hepatic acute phase plasma protein genes by hepatocyte stimulating factors and other mediators of inflammation. Mol. Biol. Med., 1990; 7: 147-159
[PubMed]
[2] Blitz M., Blitz S., Hughes R., Diner B., Beasley R., Knopp J., Rowe B.H.: Aerosolized magnesium sulfate for acute asthma: a systematic review. Chest, 2005; 128: 337-344
[PubMed] [Full Text HTML] [Full Text PDF]
[3] Bogdan C.: Nitric oxide and the immune response. Nat. Immunol., 2001; 10: 907-916
[PubMed]
[4] Bussiere F., Gueux E., Rock E., Girardeau J.P., Tridon A., Mazur A., Rayssiguier Y.: Increased phagocytosis and production of reactive oxygen species by neutrophils during magnesium deficiency in rats and inhibition by high magnesium concentration. Br. J. Nutr., 2002; 87: 107-113
[PubMed]
[5] Bussiere F., Gueux E., Rock E., Mazur A., Rayssiguier Y.: Female rats are less susceptible to inflammation induced by magnesium deficiency than males: the influence of estrogen. W: Advances in Magnesium Research: Nutrition and Health, red. Rayssiguier Y., Mazur A, Durlach J., John Libbey & Company Ltd., 2001, 313-314
[6] Bussiere F.I., Gueux E., Rock E., Mazur A., Rayssiguier Y.: Protective effect of calcium deficiency on the inflammatory response in magnesium-deficient rats. Eur. J. Nutr., 2002; 41: 197-202
[PubMed]
[7] Bussiere F.I., Tridon A., Zimowska W., Mazur A., Rayssiguier Y.: Increase in complement component C3 is an early response to experimental magnesium deficiency in rats. Life Sci., 2003; 73: 499-507
[PubMed]
[8] Cassatella M.A.: The production of cytokines by polymorphonuclear neutrophils. Immunol. Today, 1995; 16: 21-26
[PubMed]
[9] Classen C.U., Abele C., Schimatschek H.F., Friedberg K.D., Classen H.G., Haubold W.: Erythema formation in magnesium-deficient albino rats. A non-invasine model for the screening of anti-inflammatory agents and oral mineral supplements. Arzneimittelforschung, 1993; 43: 672-675
[PubMed]
[10] Claverie-Benureau S., Lebel B., Gaudin-Harding F.: Magnesium deficiency allergy-like crisis in hairless rats. A suggested model for inflammation studies. J. Physiol., 1980; 76: 173-175
[PubMed]
[11] Denda M.: New strategies to improve skin barrier homeostasis. Adv. Drug Deliv. Rev., 2002; 54, Suppl.1: 123-130
[PubMed]
[12] Denda M., Hosoi J., Asida Y.: Visual imaging of ion distribution in human epidermis. Biochem. Biophys. Res. Commun., 2000; 272: 134-137
[PubMed]
[13] Durlach J.: Magnez w praktyce klinicznej. PZWL, Warszawa, 1991
[14] Durlach J.: Rapports exorimentaux et cliniques entre magnesium et hypersensibilite. Rev. fr. Allerg. 1975; 15: 133-146
[15] Elin R.J.: Assessment of magnesium status. Clin. Chem.,1987; 33: 1965-1970
[PubMed] [Full Text PDF]
[16] Elin R.J.: The effect of magnesium deficiency in mice on serum immunoglobulin concentrations and antibody plaque-forming cells. Proc. Soc. Exp. Biol. Med., 1975; 148: 620-624
[PubMed]
[17] Frakes M.A., Richardson L.E.2nd: Magnesium sulfate therapy in certain emergency conditions, Am. J. Emerg. Med., 1997; 15: 182-187
[PubMed]
[18] Frank M.M, Fries L.F.: The role of complement in inflammation and phagocytosis. Immunol. Today, 1991; 12: 322-326
[PubMed]
[19] Gaudin-Harding F., Claverie-Benureau S., Armier J., Davy J., Lebel B.: Aromatic amines (serotonin and histamine) and magnesium deficiency in the rat. Int. J. Vitam. Nutr. Res., 1980; 50: 185-192
[PubMed]
[20] Grassberger M., Baumruker T., Enz A., Hiestand P., Hultsch T., Kalthoff F., Schuler W., Schulz M., Werner F.J., Winiski A., Wolff B., Zenke G.: A novel anti-inflammatory drug, SDZ ASM 981, for the treatment of skin diseases: in vitro pharmacology. Br. J. Dermatol., 1999; 141: 264-273
[PubMed]
[21] Greiner J., Diezel W.: Entzündungshemmende wirkung von magnesium-ionen bei der kontaktekzem-reaktion. Hautarzt, 1990; 41: 602-605
[PubMed]
[22] Grzesiak J.J., Davis G.E., Kirchhofer D., Pierschbacher M.D.: Regulation of a
2b
1-mediated fibroblast migration on type I collagen by shifts in the concentrations of extracellular Mg2+ and Ca2+. J. Cell. Biol., 1992; 117: 1109-1117
[PubMed] [Full Text PDF]
[23] Grzesiak J.J., Pierschbacher M.D.: Changes in the concentrations of extracellular Mg++ and Ca++ down-regulate E-cadherin and up-regulate a
2b
1 integrin function, activating keratinocyte migration on type I collagen. J. Invest. Dermatol., 1995; 104: 768-774
[PubMed]
[24] Hughes R., Goldkorn A., Masoli M., Weatherall M., Burgess C., Beasley R.: Use of isotonic nebulised magnesium sulphate as an adjuvant to salbutamol in treatment of severe asthma in adults: randomized placebo-controlled trial. Lancet, 2003; 361: 2114-2117
[PubMed]
[25] Hunziker N.: Magnesium and Its Role in Allergy. W: Metal Ions in Biological System, red. Siegel H., Siegel A., Marcel D., New York, 1990; 26: 531-547
[26] Kimata H., Tai H., Nakagawa K., Yokoyama Y., Nakajima H., Ikegami Y.: Improvement of skin symptoms and mineral imbalance by drinking deep water in patients with atopic eczema/dermatitis syndrome (AEDS). Acta Medica (Hradec Kralove), 2002; 45: 83-84
[PubMed]
[27] Kruse H.D., Orent E.R., McCollum E.: Studies on magnesium deficiency in animals. Symptomatology resulting from magnesium deprivation. J. Biol. Chem., 1932; 96: 519-522
[28] Kurantsin-Mills J., Cassidy M.M., Stafford R.E., Weglicki W.B.: Marked alterations in circulating inflammatory cells during cardiomyopathy development in a magnesium-deficient rat model. Br. J. Nutr., 1997; 78: 845-855
[PubMed]
[29] Laurant P., Dalle M., Berthelot A., Rayssiguier Y.: Time-course of the change in blood pressure level in magnesium-deficient Wistar rats. Br. J. Nutr., 1999; 82: 243-251
[PubMed] [Full Text HTML]
[30] Laurant P., Touyz R.M.: Physiological and pathophysiological role of magnesium in the cardiovascular system: implications in hypertension. J. Hypertens., 2000; 18: 1177-1191
[PubMed]
[31] Levi-Schaffer F., Shani J., Politi Y., Rubinchik E., Brenner S.: Inhibition of proliferation of psoriatic and healthy fibroblasts in cell culture by selected Dead-Sea salts. Pharmacology, 1996; 52: 321-328
[PubMed]
[32] Mak I.T., Dickens B.F., Komarov A.M., Wagner T.L., Phillips T.M., Weglicki W.B.: Activation of the neutrophil and loss plasma glutathione during Mg-deficiency – modulation by nitric oxide synthase inhibition. Mol. Cell Biochem., 1997; 176: 35-39
[PubMed]
[33] Mak I.T., Komarov A.M., Wagner T.L., Stafford R.E., Dickens B.F., Weglicki W.B.: Enhanced NO production during Mg deficiency and its role in mediating red blood cell glutathione loss. Am. J. Physiol., 1996; 271: C385-C390
[PubMed]
[34] Makiura M., Akamatsu H., Akita H., Yagami A., Shimizu Y., Eiro H., Kuramoto M., Suzuki K., Matsunaga K.: Atopic dermatitis-like symptoms in HR-1 hairless mice fed a diet low in magnesium and zinc. J. Int. Med. Res., 2004; 32: 392-399
[PubMed]
[35] Malpuech-Brugere C., Kuryszko J., Nowacki W., Rock E., Rayssiguier Y., Mazur A.: Early morphological and immunological alterations in the spleen during magnesium deficiency in the rat. Magnes. Res., 1998; 11: 161-169
[PubMed]
[36] Malpuech-Brugere C., Nowacki W., Daveau M., Gueux E., Linard C., Rock E., Lebreton J., Mazur A., Rayssiguier Y.: Inflammatory response following acute magnesium deficiency in the rat. Biochim. Biophys. Acta, 2000; 1501: 91-98
[PubMed]
[37] Malpuech-Brugere C., Nowacki W., Gueux E., Kuryszko J., Rock E., Rayssiguier Y., Mazur A.: Accelerated thymus involution in magnesium-deficient rats is related to enhanced apaptosis and sensitivity to oxidative stress. Br. J. Nutr., 1999; 81: 405-411
[PubMed]
[38] Mazur A., Zimowska W., Nowacki W.: Lipoproteiny w reakcji ostrej fazy. W: Białka ostrej fazy u zwierząt, red. Kostro K., Gliński Z., WAR, Lublin, 2003, 121-126
[39] McCoy J. H., Kenney M. A.: Magnesium and Immune Function: A Review. W: Magnesium in Cellular Processes and Medicine, red. Altura B.M., Durlach J., Seeling M.S., Kargel, Basel, 1987, 196-211
[40] McCoy J.H., Kenney M.A.: Magnesium and immune function: recent findings. Magnes. Res., 1992; 5: 281-293
[PubMed]
[41] Miyao N., Ikegaki I., Nishio A., Ishiguro S.: Comparative biochemical studies on the histamine metabolism in acute magnesium-deficient mice and rats. Mem. Fac. Agr. Kagoshima. Univ., 1984; 20: 111-125
[42] Montell C.: Mg2+ Homeostasis: the Mg2+ nificent TRPM chanzymes. Curr. Biol., 2003; 13: R799-R801
[PubMed] [Full Text HTML] [Full Text PDF]
[43] Nassir F. I., Mazur A., Giannoni F., Gueux E., Davidson N.O., Rayssiguier Y.: Magnesium deficiency modulates hepatic lipogenesis and apolipoprotein gene expression in the rat. Biochim. Biophys. Acta, 1995; 1257: 125-132
[PubMed]
[44] Nasulewicz A., Wietrzyk J., Wolf F.I., Dzimira S., Madej J., Maier J.A., Rayssiguier Y., Mazur A., Opolski A.: Magnesium deficiency inhibits primary tumor growth but favors metastasis in mice. Biochim. Biophys. Acta, 2004; 1739: 26-32
[PubMed]
[45] Neckermann G., Bavandi A., Meingassner J.G.: Atopic dermatitis-like symptoms in hypomagnesaemic hairless rats are prevented and inhibited by systemic or topical SDZ ASM 981. Br. J. Dermatol., 2000; 142: 669-697
[PubMed]
[46] Nishio A., Ishiguro S., Ikegaki I., Matsumoto S., Yoshimitsu F., Miyazaki A.: Histamine metabolism and pinnal hyperaemia during magnesium deficiency in rats. Magnes. Res., 1988; 1: 155-161
[PubMed]
[47] Nishio A., Ishiguro S., Matsumoto S., Miyao N.: Histamine content and histidine decarboxylase activity in the spleen of the magnesium-deficient rat: comparison with the skin and peritoneal mast cells. Jpn. J. Pharmacol., 1984; 36: 1-6
[PubMed]
[48] Nishio A., Ishiguro S., Miyao N.: Toxicological and pharmacological studies on magnesium deficiency in rats: histamine-metabolizing enzymes in some tissues of magnesium-deficient rats. Nippon Juigaku Zasshi, 1983; 45: 699-705
[PubMed]
[49] Nishio A., Miyazaki A., Ishiguro S., Miyao N.: Sex difference of pinnal hyperemia in magnesium-deficient rats: effects of castration and administration of sex hormone. Jpn. J. Pharmacol., 1986; 41: 15-22
[PubMed]
[50] Perraud A. L., Knowles H.M., Schmitz C.: Novel aspects of signaling and ion-homeostasis regulation in immunocytes. The TRPM ion channels and their potential role in modulating the immune response. Mol. Immunol., 2004; 41: 657-673
[PubMed]
[51] Ponvert C., Galoppin L., Burtin C., Paupe J., Canu P.: Noninvolvement of histamine and prostaglandins in the dermatosis of magnesium-deficient hairless rats. Pharmacology, 1984; 28: 235-240
[PubMed]
[52] Proksch E., Nissen H.P., Bremgartner M., Urquhart C.: Bathing in magnesium-rich Dead Sea salt solution improves skin barier function, enhances skin hydration, and reduces inflammation in atopic dry skin. Int. J. Dermatol., 2005; 44: 151-157
[PubMed]
[53] Rayssiguier Y., Gueux E., Nowacki W., Rock E., Mazur A.: High fructose consumption combined with low dietary magnesium intake may increase of the metabolic syndrome by inducing inflammation. Magnes. Res., 2006; 19: 237-243
[PubMed] [Full Text HTML] [Full Text PDF]
[54] Rayssiguier Y., Malpuech C., Nowacki W., Daveau M., Lebreton J.P., Rock E., Mazur A., Gueux E.: Evaluation of the inflammatory state during magnesium deficiency in the rat. Magnes. Res., 1994; 7: 51
[55] Rayssiguier Y., Malpuech C., Nowacki W., Rock E., Gueux E., Mazur A.: Inflammatory response in magnesium deficiency. W: Magnesium in cardiology and other topics of clinical, experimental and laboratory research. Fifth European Magnesium Congress, Vienna, 1995, 64: 415-421
[56] Rock E., Astier C., Lab C., Malpuech C., Nowacki W., Gueux E., Mazur A., Rayssiguier Y.: Magnesium deficiency in rats induces a rise in plasma nitric oxide. Magnes. Res.,1995; 8: 237-242
[PubMed]
[57] Rubin H.: The membrane, magnesium, mitosis (MMM) model of cell proliferation control. Magnes. Res., 2005; 18: 268-274
[PubMed]
[58] Rude R.K.: Magnesium Homeostasis. W: Principles of Bone Biology, red. Bilezikian J.P., Raisz L.G., Rodan G.A., Academic Press, 1996, 277-293
[59] Saris N.E.L., Mervaala E., Karppanen H., Khawaja J.A., Lewenstam A.: Magnesium An update on physiological, clinical and analytical aspects. Clin. Chim. Acta, 2000; 294: 1-26
[PubMed]
[60] Saurat J.H., Chavaz P., Barbier A., Faucher F.: Dermatosis in hairless rats fed a low magnesium diet. W: Models in Dermatology, red. Maibach, Lowe, Karger, Basel, 1985; 1: 202-209
[61] Schempp C.M., Dittmar H.C., Hummler D., Simon-Haarhaus B., Schulte-Monting J., Schopf E., Simon J.C.: Magnesium ions inhibit the antigen-presenting function of human epidermal Langerhans cells in vivo and in vitro. Involvement of ATPase, HLA-DR, B7 molecules, and cytokones. J. Invest. Dermatol., 2000; 115: 680-686
[PubMed] [Full Text HTML] [Full Text PDF]
[62] Schmitz C., Deason F., Perraud A.L.: Molecular components of verebrate Mg2+-homeostasis regulation. Magnes. Res., 2007; 20: 6-18
[PubMed] [Full Text HTML] [Full Text PDF]
[63] Shani J., Sulliman A., Katzir I., Brenner S.: Penetration of selected Dead Sea minerals through a healthy rabbit skin, from a sustained-release transparent varnish, as a prospective treatment for psoriasis. J. Eur. Acad. Dermatol. Venereol., 1995; 4: 267-272
[64] Smith B.P.: Large Animal Internal Medicine, Mosby, ST Louis, USA, 1996, 455, 1474-1479
[65] Stachura J.: Badania morfologiczne w niedoborze i nadmiarze magnezu u szczura. Patol. Pol., 1971; 22: 41-53
[PubMed]
[66] Tanaka S., Mikura S., Hashimoto E., Sugimoto Y., Ichikawa A.: Ca2+ influx – mediated histamine synthesis and IL-6 release in mast cells activated by monomeric IgE. Eur. J. Immunol., 2005; 35: 460-468
[PubMed]
[67] Tong G.M., Rude R.K.: Magnesium deficiency in critical illness. J. Intensive Care Med., 2005; 20: 3-17
[PubMed]
[68] Trzeciakiewicz A., Opolski A., Mazur A.: TRPM7: a protein responsible for magnesium homeostasis in a cell. Post. Hig. Med. Dośw., 2005, 59: 496-502
[PubMed] [Full Text HTML] [Full Text PDF]
[69] Van Orden R., Eggett D.L., Franz K.B.: Influence of graded magnesium deficiencies on white blood cell counts and lymphocytes subpopulations in rats. Magnes. Res., 2006; 19: 93-101
[PubMed]
[70] Weglicki W.B., Mak I.T., Kramer J.H., Dickens B.F., Cassidy M.M., Stafford R.E., Phillips T.M.: Role of free radicals and substance P in magnesium deficiency. Cardiovasc. Res., 1996; 31: 677-682
[PubMed]
[71] Weglicki W.B., Phillips T.M.: Pathobiology of magnesium deficiency: a cytokine/neurogenic inflammation hypothesis. Am. J. Physiol., 1992; 263: R734-R737
[PubMed]
[72] Weglicki W.B., Phillips T.M., Freedman A.M., Cassidy M.M., Dickens B.F.: Magnesium-deficiency elevates circulating levels of inflammatory cytokines and endothelin. Mol. Cell. Biochem., 1992; 110: 169-173
[PubMed]
[73] Wolf F.I., Cittadini A.: Magnesium in cell proliferation and differentiation. Front. Biosci., 1999; 4: D607-D617
[PubMed]
[74] Yokoyama T., Oono H., Miyamoto A., Ishiguro S., Nishio A.: Magnesium-deficient medium enhances NO production in alveolar macrophages isolated from rats. Life Sci., 2003; 72: 1247-1257
[PubMed]
[75] Zimmermann P., Weiss U., Classen H. G., Wendt B., Epple A., Zollner H., Temmel W., Weger M., Porta S.: The impact of diets with different magnesium contents on magnesium and calcium in serum and tissues of the rat. Life Sci., 2000; 67: 949-958
[PubMed]