
The structural diversity of lipid A from gram-negative bacteria
Jolanta Lodowska 1 , Daniel Wolny 2 , Ludmiła Węglarz 1 , Zofia Dzierżewicz 2
Streszczenie
Większość bakterii Gram-ujemnych to gatunki chorobotwórcze dla ludzi i zwierząt. Najbardziej aktywnym biologicznie związkiem uczestniczącym w infekcjach jest lipopolisacharyd (LPS). Dla podkreślenia jego działań negatywnych nazwano go endotoksyną bakteryjną. Za owe niepożądane działania odpowiada lipid A – jeden z trzech podstawowych komponentów strukturalnych LPS. Choć jest on uważany za stosunkowo konserwatywny strukturalnie składnik endotoksyn, to międzygatunkowa, a nawet międzyszczepowa heterogenność może obejmować wiele elementów jego budowy chemicznej, np. typ reszt aminocukrowych, stopień podstawienia rdzenia disacharydowego kwasami tłuszczowymi i ugrupowaniami obdarzonymi ładunkiem, np. grupami fosforanowymi i/lub etanoloaminą, a także rodzaj, liczbę i rozmieszczenie kwasów tłuszczowych występujących w lipidzie A. Szkielet cukrowy lipidu A u większości bakterii Gram-ujemnych tworzą dwie cząsteczki glukozaminy połączone wiązaniem b
(1–>6) glikozydowym, których grupy aminowe (przy atomach węgla 2, 2’) oraz hydroksylowe w pozycjach 3, 3’ podstawione są zazwyczaj 3-hydroksykwasami tłuszczowymi, najczęściej kwasem 3-hydroksytetradekanowym. Przez grupę hydroksylową tych związków są połączone estrowo inne (zazwyczaj nasycone, nierozgałęzione) kwasy tłuszczowe. W lipidach A poszczególnych mikroorganizmów występuje stosunkowo duże zróżnicowanie kwasów tłuszczowych, począwszy od kwasu mirystynowego (kwasu tetradekanowego; 14:0) i kwasu laurynowego (kwasu dodekanowego; 12:0) oraz ich hydroksylowych pochodnych, a skończywszy na tak unikatowych strukturach jak kwas cis-11-oktadecenowy (Rhodospirillum salinarum 40), kwas 3-hydroksy-5-dodecenowy (Phenylobacterium immobile) i kwas izo-2,3-dihydroksytetradekanowy (Legionella pneumophila). Rdzeń cukrowy lipidów A niektórych bakterii może budować także inny niż glukozamina cukier, tj. 2,3-diamino- 2,3-dideoksy-D-glukoza. Wśród podstawników występujących w tym regionie LPS oprócz reszt fosforanowych i etanoloaminy można także wymienić b
-mannopiranozę, 4-aminoarabinozę, kwas galaktouronowy i glicynę. Tak więc lipid A, choć uznawany za względnie zachowawczy strukturalnie komponent endotoksyn, cechuje się stosunkowo dużą zmiennością strukturalną, co przekłada się na zróżnicowanie aktywności biologicznej tych makrocząsteczek.
Słowa kluczowe:lipopolisacharyd • endotoksyna • bakterie Gram-ujemne • lipid A
Summary
The majority of Gram-negative bacteria are pathogenic to humans and animals. Lipopolysaccharide (LPS) is the most biologically active component of these microorganisms. This compound is also called endotoxin to emphasize its negative impact on a macroorganism. Lipid A, one of the three structural components of the LPS molecule, is responsible for the pathophysiological effects associated with Gram-negative bacteria infections. Although lipid A is considered the conservative component of endotoxin, differences in its structure among species and even strains may occur. These differences concern the type of aminosugars, the degree of substitution of the disaccharide core by fatty acids, phosphate, and/or ethanolamine, and also the type, quantity, and distribution of fatty acids. The lipid A saccharide backbone of the majority of Gram-negative bacteria consists of two glucosamine units in b
(1–>6) glycosidic linkage. Amino groups (at positions 2 and 2’) and hydroxy groups (at positions 3 and 3’) of glucosamines are commonly substituted by 3-hydroxyfatty acids, most often by 3-hydroxytetradecanoic acid. Other fatty acids (usually saturated, unbranched) are ester-linked to hydroxyacids by their hydroxy group. In lipid A of different microorganisms there is a high diversity of fatty acids, from mirystic (tetradecanoic, 14:0) and lauric (dodecanoic, 12:0) acids and their hydroxylated derivatives to such unique structures as cis-11-octadecenoic acid (Rhodospirillum salinarum 40), 3-hydroxy-5-dodecenoic acid (Phenylobacterium immobile), and iso-2,3-dihydroxytetradecanoic acid (Legionella pneumophila). The saccharide core of some bacterial lipid A may consist of sugars different from glucosamine, e.g., 2,3-diamino- 2,3-dideoxy-D-glucose. Other substituents of this part of LPS, besides phosphate groups and ethanolamine, are b
-mannopyranose, 4-aminoarabinose, galacturonic acid, and glycine. Therefore, lipid A, though considered the relatively conservative component of endotoxin, reveals relatively large structural diversity, which influences the variety of LPS biological activity.
Key words:lipopolysaccharide • endotoxin • Gram-negative bacteria • lipid A
WSTĘP
Lipid A jest komponentem strukturalnym bakteryjnych endotoksyn, czyli lipopolisacharydów (LPS) uczestniczących w patogenezie wielu chorób wywoływanych przez bakterie Gram-ujemne, lecz mogących także wpływać korzystnie na zainfekowany makroorganizm. Ten integralny składnik błony zewnętrznej bakterii Gram-ujemnych zajmujący aż 70% powierzchni komórki bakteryjnej jest niezbędny do funkcjonowania drobnoustroju. LPS chroni go przed obronnymi mechanizmami makroorganizmu oraz działaniem kwasów żółciowych i hydrofobowych antybiotyków [34]. W przebiegu infekcji uwolniony z powierzchni komórek bakteryjnych LPS może być przyczyną złożonych reakcji patofizjologicznych. Stymulacja endotoksyną leukocytów, a szczególnie monocytów i makrofagów oraz komórek śródbłonka skutkuje uwolnieniem licznych cytokin i czynników zapalnych, m.in. interleukin (IL-1, -6, -8, -10), czynnika martwicy nowotworu (TNF-a
), produktów metabolizmu kwasu arachidonowego (prostaglandyn, leukotrienów), czynnika aktywującego płytki krwi, wolnych rodników tlenowych, nadtlenku wodoru i tlenku azotu. Substancje te wzmacniają zdolności obronne organizmu, jednak uwolnione gwałtownie w stanach uogólnionego zakażenia bakteriami Gram-ujemnymi mogą powodować uszkodzenie tkanek nieobjętych infekcją, a ich zwiększona sekrecja powoduje wysoką gorączkę, hipotensję, tachykardię, przyspieszony oddech, wewnątrznaczyniowe wykrzepianie krwi oraz wstrząs septyczny. Korzystne działanie endotoksyn ujawnia się natomiast przy niskim poziomie mediatorów, co skutkuje stanem podgorączkowym, zwiększeniem odporności na infekcje, aktywnościami adiuwantowymi [53,66,76,81].
Pojedyncza cząsteczka LPS składa się z trzech odrębnych regionów różniących się budową chemiczną, właściwościami biologicznymi oraz zmiennością strukturalną. Jest to łańcuch O-swoisty, rdzeń oligosacharydowy oraz lipid A. Schemat budowy LPS przedstawia ryc. 1.

Ryc. 1. Ogólny schemat budowy LPS [52]
ZMIENNOŚĆ STRUKTURALNA ANTYGENU O
Łańcuch O-swoisty jest najbardziej zewnętrzną częścią LPS, tworzącą charakterystyczny i unikatowy dla danego szczepu antygen somatyczny. Określa swoistość serologiczną, pełni rolę antygenu powierzchniowego bakterii Gram-ujemnych (antygen O). Zbudowany jest z kilkunastu do kilkudziesięciu powtarzających się podjednostek oligosacharydowych, zawierających od jednej do ośmiu reszt węglowodanowych. Oprócz cukrów obojętnych, tj. heksoz (galaktoza, glukoza, mannoza) i heksozamin (glukozamina, galaktozamina) w antygenie O stwierdza się obecność kwasów uronowych, 6-deoksyheksoz (ramnoza, fukoza, chinowoza, 6-deoksyaltroza, 6-deoksytaloza) oraz 3,6-dideoksyheksoz (abekwoza, kolitoza, paratoza, tyweloza). U wielu gatunków bakterii w antygenie O zidentyfikowano składniki niecukrowe (aminokwasy, fosforan, fosfoetanoloaminę, glicerol, rybitol) oraz inne rzadko spotykane w przyrodzie związki chemiczne, m.in. 3,6-dideoksy- 3-[(R)-3-hydroksybutyroamido]-D-glukozę u Hafnia alvei 1216 [42], N-hydroksyetylo-D-alaninę u Proteus mirabilis 03 [105], kwas N-acetylomuraminowy u P. penneri 62 [45], kwas N-acetyloizomuraminowy u P. penneri 35 [44], kwas 7-acetamido-3,4,7,9-tetradeoksy-5-(4-hydroksybutyroamido)- D-glicero-L-galakto-nonulozonowy u Yersinia ruckerii 01 [4], kwas 3-hydroksy-2,3-dimetylo-5-oksopirolidyno-2- karboksylowy u Plesiomonas shigelloides [64]. Struktury powtarzających się podjednostek, determinowane przez stereokonfigurację, ich sekwencje i pozycje wiązań oraz lokalizację niecukrowych podstawników, warunkują zróżnicowanie międzyszczepowe w obrębie gatunku, stanowiąc o zmienności strukturalnej tej części LPS [79]. Jednak endotoksyna wyizolowana z zewnętrznej błony osłony komórkowej niektórych mikroorganizmów nie ma łańcucha O-swoistego. Tego komponentu endotoksyny nie zidentyfikowano w LPS bakterii gatunków Chlamydia trachomatis, C. psittaci, Haemophilus influenzae, Campylobacter jejuni, Bordetella pertussis czy Neisseria meningitidis [79], oraz w formach szorstkich (R) drobnoustrojów z rodziny Enterobacteriaceae. Tylko jedną jednostkę antygenu O zawierają natomiast formy półszorstkie (SR) z rodziny Enterobacteriaceae [31]. Schemat budowy różnych form LPS bakterii Gram-ujemnych przedstawiono na ryc. 2.

Ryc. 2. Schemat budowy R-LPS, SR-LPS, S-LPS bakterii Gram-ujemnych
ZRÓŻNICOWANIE STRUKTURALNE OLIGOSACHARYDU RDZENIOWEGO LPS
Region rdzeniowy łączący łańcuch O-swoisty z lipidem A wykazuje mniejsze zróżnicowanie strukturalne niż antygen O. W rdzeniu można wyróżnić część dystalną względem lipidu A (region zewnętrzny) oraz proksymalną (region wewnętrzny). Pierwszy z nich ze względu na zawartość heksoz (np. glukozy, galaktozy, glukozaminy, galaktozaminy) nazywany jest również regionem heksozowym. Rdzeń wewnętrzny zbudowany z heptopiranoz (głównie w konfiguracji L-glicero-D-manno-) i kwasu 3-deoksyoktulozonowego (Kdo) stanowi natomiast region heptozowy. Obecnie wiadomo, że istnieją lipopolisacharydy, których budowa odbiega od tego schematu, bowiem w regionie heksozowym identyfikuje się heptozy, a w części heptozowej – heksozy. W regionie wewnętrznym rdzenia występują również kwasy uronowe, grupy fosforanowe, pirofosforanowe, etanoloamina lub aminokwasy, np. glicyna [31,115]. Związek ten zidentyfikowano w regionie rdzeniowym wielu bakterii, np. Shigella, Escherichia, Salmonella, Hafnia czy Citrobacter [22,23]. U Providencia rettgeri [3] i Shigella sonnei oprócz glicyny w regionie rdzeniowym zidentyfikowano również alaninę, a u Rhodopseudomonas spherides [83] stwierdzono obecność treoniny. W regionie rdzeniowym LPS oprócz Kdo mogą również występować inne pochodne kwasowe cukrów, np. kwas N-acetyloneuraminowy u Rhodobacter sp. [48], Campylobacter jejuni, C. fetus [1,2], kwas 2-keto-3-deoksy-1,7-dikarboksyheptonowy u Acinetobacter calcoaceticus [10] oraz kwas D-glicero-D-talo-oktulozonowy (Ko) u A. calcoaceticus [24] i Burkholderia cepacia [37]. W lipopolisacharydach Vibrio cholerae stwierdzono obecność fruktozy [38], a w V. parahaemolyticus kwasu 3-deoksy-D-treo-heksulozonowego [46]. Różnice w strukturze rdzenia mogą występować, nie tylko między gatunkami, lecz również w ich obrębie. Po kilka rodzajów oligosacharydu rdzeniowego stwierdzono, m.in. u Escherichia coli, Neisseria meningitidis, Campylobacter, Citrobacter oraz w szczepach Haemophilus i Bacteroides [1,2,33]. Znane są także drobnoustroje mające defektywną (skróconą) strukturę tego komponentu LPS, np. Salmonella minnesota. Całkowicie pozbawione są rdzenia zewnętrznego bakterie Chlamydia trachomatis, C. psittaci i Haemophilus influenzae [79].
MIĘDZYGATUNKOWE ZRÓŻNICOWANIE STRUKTURALNE LIPIDU A
Lipid A jest powszechnie uważany za najbardziej konserwatywny strukturalnie region endotoksyn bakterii Gramujemnych. Jednakże i tę komponentę cechuje międzygatunkowe, a nawet międzyszczepowe zróżnicowanie strukturalne. Jedną z najwcześniej poznanych była struktura lipidu A bakterii E. coli [36]. Jego rdzeniem cukrowym są dwie cząsteczki D-glukozaminy (D-GlcN) połączone wiązaniem b
(1–>6) glikozydowym. Ten disacharyd podstawiony jest dwiema grupami fosforanowymi. Jedna jest związana estrowo w pozycji 4’ GlcN II (koniec nieredukujący, dystalny), natomiast druga a
-glikozydowo w pozycji 1 GlcN I (koniec redukujący, proksymalny). Grupa hydroksylowa przy 4 atomie węgla GlcN I jest wolna, natomiast przy 6’ atomie węgla GlcN II jest podstawiona oligocukrem rdzenia przez ketozydowo połączoną cząsteczkę Kdo. Do glukozaminowego rdzenia w pozycjach 3, 3’ przyłączone są estrowo dwie reszty kwasu b
-hydroksymirystynowego, a kolejne są związane amidowo w pozycjach 2, 2’. Natomiast grupa hydroksylowa amidowo związanej reszty kwasu tłuszczowego przy nieredukującej cząsteczce GlcN podstawiona jest kwasem laurynowym, tworząc pentaacylową strukturę. W lipidzie A bakterii E. coli grupa hydroksylowa acylu przy 3’ jest estryfikowana dodatkową resztą kwasu mirystynowego (ryc. 3). Rozmieszczenie kwasów tłuszczowych w lipidzie A tych mikroorganizmów jest asymetryczne.

Ryc. 3. Schemat budowy lipidu A E. coli [86]
W lipidzie A można zatem wyróżnić trzy podregiony: najczęściej disacharydowy komponent cukrowy, kwasy tłuszczowe oraz hydrofilowe podstawniki, np. reszty fosforanowe.
Najmniejszą zmienność strukturalną wykazuje fragment cukrowy lipidu A. Najczęściej jest nim disacharyd zbudowany z dwóch cząsteczek glukozaminy połączonych wiązaniem b(1–>6) glikozydowym, który zidentyfikowano nie tylko u E. coli, lecz także na przykład w lipidzie A bakterii Pseudomonas aeruginosa [50], Vibrio cholerae [13], Chromobacterium violaceum [26], Bordetella pertussis [14].
W lipidach A niektórych mikroorganizmów, np. Bartonella henselae, Legionella pneumophila, Leptospira interrogans, Mesorhizobium huakuii, Aquifex pyrophilus, Pseudomonas diminuta (ryc. 4), P. vesicularis czy Bdellovibrio bacteriovorus zamiast disacharydu glukozaminylowego występują dwie cząsteczki 2,3-diamino-2,3-dideoksyglukozy (GlcpN3N, DAG) połączone wiązaniem b
(1–>6) glikozydowym (lipid ADAG) [15,39,67,70,87,107,112,113]. U niektórych bakterii stwierdzono występowanie „mieszanego lipidu A”, w którym GlcN3N połączona jest z GlcN. Tego typu struktura chemiczna występuje w lipopolisacharydach bakterii z rodzaju Thiobacillus, Brucella oraz w gatunkach Campylobacter jejuni (ryc. 5) [60,63] i Chlorobium vibrioforme [57].

Ryc. 4. Struktura chemiczna lipidu A P. diminuta (? – niezidentyfikowany podstawnik, R – reszty acylowe) [107]

Ryc. 5. Schemat budowy lipidu A C. jejuni [60,63]
Badając endotoksyny bakterii Campylobacter jejuni Moran i wsp. [63] stwierdzili obecność w izolacie aż trzech postaci lipidu A, różniących się budową komponenty węglowodanowej: disacharyd glukozaminylowo-glukozaminylowy (12%), strukturę złożoną z 2 cząsteczek diaminoglukozy (15%) oraz ilościowo dominującą (73%) formę mieszaną składającą się z diaminoglukozy i glukozaminy (tab. 1).
Tabela 1. Stopień ufosforylowania i rodzaj podstawienia rdzenia węglowodanowego lipidu A bakterii Gram-ujemnych
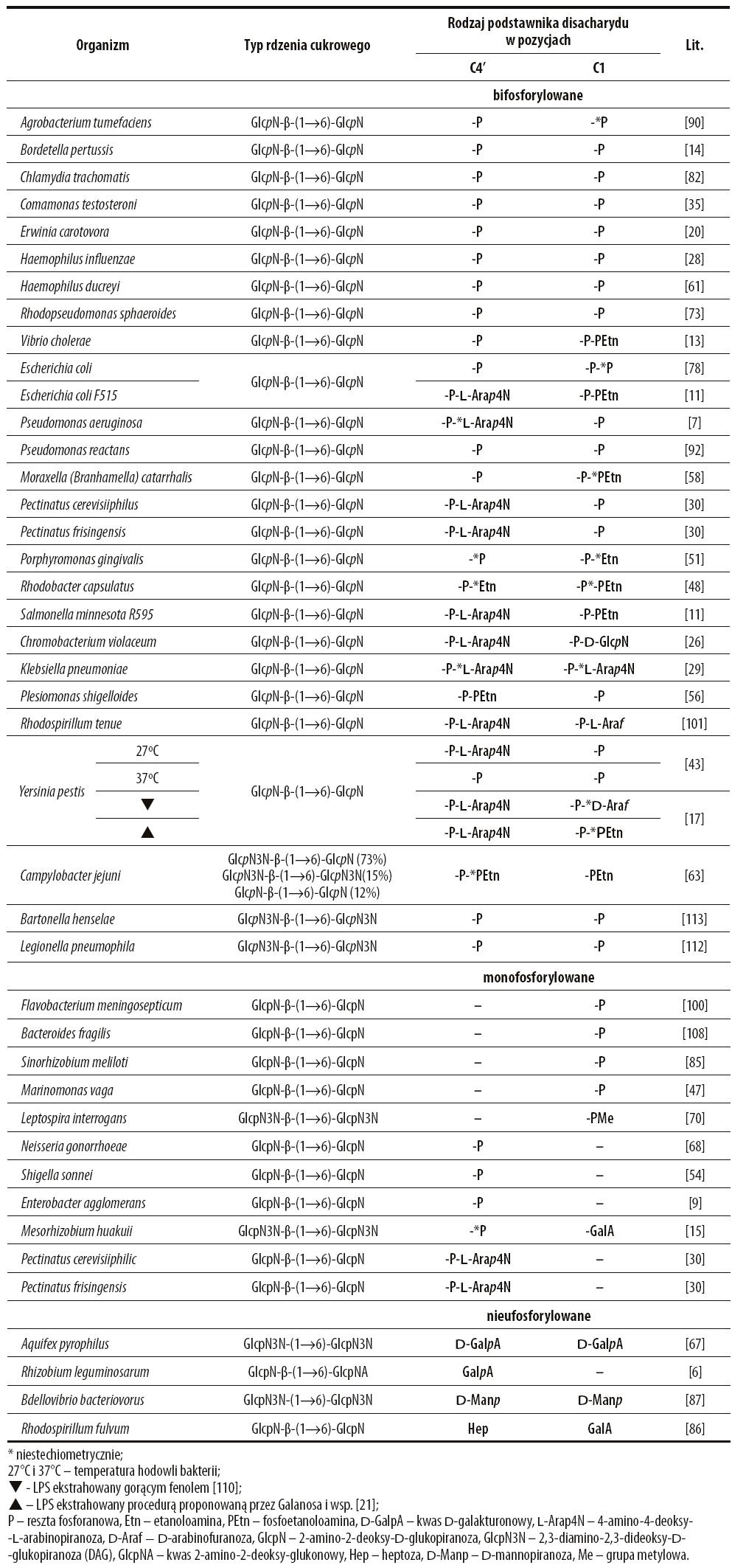
Unikalną strukturę cukrową ma lipid A Rhizobium leguminosarum, bowiem redukująca reszta glukozaminy zastąpiona jest kwasem 2-amino-2-deoksyglukonowym [6].
Disacharyd lipidu A jest połączony z hydrofilowymi podstawnikami, które w znacznym stopniu determinują heterogenność tego komponentu LPS, tym bardziej że mogą one występować w ilościach niestechiometrycznych. U wielu bakterii Gram-ujemnych rdzeń cukrowy lipidu A w pozycjach 1 i 4′ jest ufosforylowany. Te grupy funkcyjne mogą być podstawione etanoloaminą (Porphyromonas gingivalis [51,98], Campylobacter jejuni [63]), fosfoetanoloaminą (Rhodobacter capsulatus [48], Moraxella catarrhalis [58], Neiseria meningitidis i Salmonella typhimurium (ryc. 6) [103]), grupą metylową (Leptospira interrogans [70]), 4-amino-4-deoksy-L-arabinozą (Salmonella typhimurium (ryc. 6), Pseudomonas aeruginosa (ryc. 7) [103], D-arabinofuranozą (Yersinia pestis [17], Rhodospirillum tenue [101]), lub dodatkową – trzecią resztą glukozaminy (Chromobacterium violaceum [26]).

Ryc. 6. Schemat struktury lipidu A N. meningitidis (A) oraz S. typhimurium (B) [103]

Ryc. 7. Schemat budowy lipidu A P. aeruginosa [103]
W lipidach A niektórych bakterii (Aquifex pyrophilus [67], Rhizobium leguminosarum [6], Bdellovibrio bacteriovorus [87]) w pozycjach 1 i 4’ disacharyd nie jest ufosforylowany, lecz bezpośrednio podstawiony dodatkowymi resztami węglowodanowymi, np. kwasem galakturonowym, 4-aminoarabinozą lub mannozą (tab. 1). W lipidzie A Rhodomicrobium vannielii [32], Rhodopseudomonas acidophila, Chromatium vinosum, Thiocapsa roseopersicina występuje mannoza [60], a u fotosyntetyzującej bakterii Rhodocyclus tenuis dodatkowa cząsteczka GlcN [101].
Badania Kawahary i wsp. [43] nad strukturą lipidu A bakterii Yersinia pestis wykazały, że rodzaj hydrofilowych podstawników zależy od temperatury, w której hodowano mikroorganizmy. W 37°C szkielet cukrowy lipidu A tych drobnoustrojów przy C4’ jest podstawiony resztą fosforanową, a w 27°C fosfo-4-amino-4-deoksy-L-arabinozą (tab. 1). Dalla Venezia i wsp. [17] analizując skład chemiczny endotoksyn wspomnianych mikroorganizmów zidentyfikowali różne podstawniki komponentu cukrowego lipidu A przy zastosowaniu odmiennych metod ekstrakcji LPS. Badane heteropolimery izolowane metodą Westphala i wsp. [110] przy atomie C1 redukującej GlcN były podstawione fosfo- D-arabinofuranozą, natomiast ekstrahowane procedurą wg Galanosa i wsp. [21] – pirofosfoetanoloaminą.
Międzygatunkowe zróżnicowanie strukturalne lipidu A dotyczy także stopnia podstawienia komponenty disacharydowej kwasami tłuszczowymi, ich rozmieszczenia oraz rodzaju reszt acylowych. W tabeli 2 zestawiono te informacje w odniesieniu do glukozaminylo-glukozaminowych lipidów A wybranych bakterii Gram-ujemnych.
Tabela 2. Rodzaj reszt acylowych i sposób ich rozmieszczenia w rdzeniu glukozaminylo-glukozaminowym lipidu A bakterii Gram-ujemnych

Ze względu na stopień podstawienia kwasami tłuszczowymi disacharydu lipidu A wyróżnia się triacylowany lipid A (Porphyromonas (Bacteroides) gingivalis) [65], tetraacylowany lipid A (Francisella tularensis) [106], pentaacylowany lipid A (Rhodobacter capsulatus [48], Rhodopseudomonas sphaeroides (ryc. 8) [73,95], Pseudomonas aeruginosa [50], Bacteroides fragilis [108]), heksaacylowany lipid A (Escherichia coli [88], Salmonella typhimurium [74], Haemophilus influenzae [28], Providencia rettgeri [3], Neisseria gonorrhoeae [96], N. meningitidis [48]), heptaacylowany lipid A (Proteus mirabilis) [89]. Lipidy A zawierające parzystą liczbę kwasów może cechować symetryczne (Comamonas testosteroni: 3+3 [35]) lub asymetryczne (Haemophilus influenzae: 4+2 [28]) ich rozmieszczenie. Zazwyczaj kwasami tłuszczowymi związanymi bezpośrednio ze składnikiem cukrowym są 3-hydroksy- lub 3-oksokwasy (Rhodopseudomonas sphareoides (ryc. 8), Rhodobacter capsulatus) [48,71,73,84], choć disacharyd w lipidach A niektórych bakterii może być podstawiony przy C3, 3’ kwasami 16:0 i 15:0 (Coxiella burnetii) [102] lub 14:0 (Chlamydia trachomatis) [82]. Dla struktury chemicznej lipidu A mikroorganizmów z rodzaju Chlamydia jest charakterystyczna obecność związanych estrowo ze składnikiem cukrowym kwasów tłuszczowych niemających grup hydroksylowych [27,114]. W lipidzie A identyfikowane są także kwasy 2- hydroksy, takie jak np. u Chromobacterium violaceum (12:0(2OH)) [111], Salmonella minnesota (14:0(2OH)) [72], kwasy w formie izo- u Flavobacterium meningosepticum (i15:0) [41,100], Bacteroides fragilis (i15:0) [108] i kwasy nienasycone, np. u Rhodobacter sphareoides (14:1) [73,84], R. capsulatus (12:1) [48]. W grupie tej można także znaleźć długołańcuchowe hydroksykwasy, takie jak 30:0(29OH) lub 28:0(27OH) u Sinorizobium meliloti [18,85]. W lipidzie A bakterii Agrobacterium tumefaciens występuje kwas 28:0(27OH), którego grupa hydroksylowa podstawiona jest kwasem 4:0(3OH). Oprócz międzygatunkowego zróżnicowania profilu kwasów tłuszczowych lipidu A, u bakterii Marinomonas vaga [47], Pectinatus cerevisiiphilus i P. frisingensis [30], Francisiella tularensis [106] czy Helicobacter pylorii [62] stwierdzono również heterogenność międzyszczepową. Kawahara i wsp. [43] badając strukturę lipidu A bakterii Yersinia pestis wykazali, że ilość kwasów tłuszczowych w tej strukturze może zależeć od temperatury, w której namnażano mikroorganizmy (tab. 2).

Ryc. 8. Struktura chemiczna lipidu A R. sphaeroides [73]
Lipid A Rhizobium etli cechuje wyjątkowa struktura – glukozamina jest połączona z kwasem 2-amino-2-deoksyglukonowym, a reszta hydroksyacylowa przy grupie aminowej tego cukru podstawiona jest kwasem 27-hydroksyoktakozanowy (28:0(27OH)) [69]. Ten długołańcuchowy hydroksykwas zidentyfikowano także w lipidzie A Brucella sp. i Sinorhizobium [8,18] oraz Agrobacterium tumefaciens [90]. W lipopolisacharydach stwierdzono również obecność innych długołańcuchowych kwasów tłuszczowych, np. u Pseudomonas carboxydovorans i Bradyrhizobium lupini 26:0(25OH) [5], u Rhodopseudomonas palustris i Rhizobium tropici 29:0(28OH) [5,25]. Reszty acylowe zawierające ponad 20 atomów węgla zidentyfikowano także u bakterii Legionella bozemanii, L. longbeachae, L. feeleii, L. hackeliae, L. jordanis [93,94] oraz Chlamydia trachomatis i Chlamydophila psittaci [27]. W lipidach A niektórych bakterii obecne są kwasy tłuszczowe zawierające więcej niż jedną grupę hydroksylową, np. i14: 0(2,3diOH) i 14: 0(2,3diOH) u Legionella pneumophila [112]. W lipidzie A mogą także występować kwasy nienasycone, bowiem u Phenylobacterium immobile zidentyfikowano 12:15(3OH) [109], a u Rhodospirillum salinarum 40 – kwas cis18:111 [75].
STRUKTURA CHEMICZNA LIPIDU A A JEGO FUNKCJA BIOLOGICZNA
Unikatowa struktura lipidu A odzwierciedla jego swoistą rolę w aktywności biologicznej LPS, stanowiąc centrum toksyczności tej makrocząsteczki [40]. Zależność aktywności biologicznej lipopolisacharydu od jego struktury chemicznej przedstawiono na ryc. 9 i 10.

Ryc. 9. Modyfikacje hydrofilowej komponenty lipidu A a aktywność biologiczna bakteryjnych endotoksyn, na przykładzie zmian strukturalnych w lipidzie A E. coli wraz ze współczynnikami określającymi stopień zmniejszenia aktywności zmodyfikowanego lipidu A w odniesieniu do kompletnej struktury tego komponentu LPS [77]

Ryc. 10. Wpływ modyfikacji strukturalnych hydrofobowej składowej lipidu A na aktywność biologiczną bakteryjnych endotoksyn na przykładzie zmian strukturalnych w lipidzie A E. coli wraz ze współczynnikami określającymi stopień zmniejszenia aktywności zmodyfikowanego lipidu A w odniesieniu do kompletnej struktury tej części LPS [77]
Pełną aktywność endotoksyczną wykazuje lipid A bakterii E. coli lub jego syntetyczne analogi o zbliżonej strukturze. Jest to uwarunkowane podstawieniem glukozaminylo-glukozaminy sześcioma resztami acylowymi, rozmieszczonymi tak jak w lipidzie A E. coli (ryc. 3) oraz połączeniem tego komponentu LPS z jedną resztą Kdo. Jakiekolwiek odstępstwa od tej struktury skutkują zmniejszeniem endotoksyczności cząsteczki. Monosacharydowe pochodne (z jedną resztą heksozaminy), będące syntetycznymi analogami lipidu A, nie wykazują aktywności biologicznych lub mają tylko niektóre z nich. Struktury odpowiadające tylko nieredukującej części lipidu A mogą pobudzać komórki do wytwarzania TNF-a
i IL-1, ale nie są pirogenne [81].
Na obniżenie aktywności biologicznej LPS ma wpływ zmniejszenie ilości podstawników acylowych [55,77]. Lipidy A o pięciu lub siedmiu podstawnikach acylowych są stukrotnie mniej aktywne niż struktura o sześciu takich resztach, a tetraacylowany lipid A jest prawie nieaktywny biologicznie. Lipidy A o dwóch lub czterech resztach kwasów tłuszczowych są słabszymi inhibitorami wiązania LPS przez makrofagi niż struktury heksaacylowane, takie jak lipid A E. coli. Deacylowany lipid A jest pod tym względem nieaktywny. Obecność kwasów tłuszczowych połączonych amidowo z disacharydem powoduje antygenowe, lecz nie pirogenne właściwości tego komponentu LPS. Podstawienie grup hydroksylowych resztami acetylowymi lub bursztynylowymi nie ma wpływu na aktywność biologiczną lipidu A [97]. Liczba kwasów tłuszczowych prawdopodobnie pełni istotną rolę w aktywacji komórek do wydzielania cytokin. Acylowany 4 resztami lipid A nie stymuluje ludzkich monocytów do wytwarzania IL-1, IL-6 i TNF-a
, a z 7 kwasami tłuszczowymi (S. minnesota, C. violaceum) jest mniej aktywny niż lipid A E. coli.
Aktywność biologiczną lipidu A warunkuje nie tylko liczba i rodzaj kwasów tłuszczowych, lecz także ich rozmieszczenie w tej strukturze [80,86]. Reszty acylowe mogą być rozmieszczone symetrycznie, np. w lipidach A bakterii Chromobacterium violaceum [26], Neisseria meningitidis, N. gonorrhoeae [111], lub w bardziej aktywnej postaci asymetrycznej, syntetyzowanej przez bakterie Escherichia coli, Salmonella sp., Haemophilus influenzae, Providencia retgerii, Moraxella catarrhalis [58], Bacteroides fragilis [108].
Na aktywność endotoksyny ma także wpływ obecność reszt fosforanowych oraz ich liczba. Rietschel i wsp. [77] twierdzą, że im mniej reszt fosforanowych zawiera cząsteczka lipidu A, tym mniejszą wykazuje on aktywność. Tezę tę potwierdza niewielka aktywność biologiczna endotoksyny Francisella tularensis, której lipid A pozbawiony jest grup fosforanowych [106].
Ponadto, konformacja lipidu A ma wpływ na właściwości biologiczne lipopolisacharydu. Największą aktywność biologiczną wykazuje lipid A o stożkowatym kształcie, występujący w podwójnie ufosforylowanych heksaacylowych cząsteczkach lipidu A z asymetrycznym ułożeniem kwasów tłuszczowych na resztach glukozaminy. Tym samym struktura ta ma większą część hydrofobową niż hydrofilową, a więc większy kąt odchylenia szkieletu glukozamin od powierzchni tworzonej przez kwasy tłuszczowe (>45°). Monofosforan heksaacylowego lipidu A ma mniejszy kąt nachylenia, dlatego kształt jego struktury zmienia się w bardziej cylindryczny, zmniejszając tym samym aktywność biologiczną cząsteczki. Natomiast całkowicie pozbawione właściwości toksycznych są lipidy A o kształcie cylindrycznym mające cztery, pięć lub symetrycznie rozmieszczonych sześć reszt acylowych o kącie nachylenia mniejszym niż 25° [55].
PIŚMIENNICTWO
[1] Aspinall G.O., McDonald A.G., Raju T.S., Pang H., Kurjanczyk L.A., Penner J.L., Moran A.P.: Chemical structure of the core region of Campylobacter jejuni serotype O:2 lipopolysaccharide. Eur. J. Biochem., 1993; 213: 1029-1037
[PubMed]
[2] Aspinall G.O., McDonald A.G., Raju T.S., Pang H., Moran A.P., Penner J.L.: Chemical structures of the core regions of Campylobacter jejuni serotypes O:1, O:4, O:23 and O:36 lipopolysaccharides. Eur. J. Biochem., 1993; 213: 1017-1027
[PubMed]
[3] Basu S., Radziejewska-Lebrecht J., Mayer H.: Lipopolysaccharide of Providencia rettgeri. Chemical studies and taxonomical implications. Arch. Microbiol., 1986; 144: 213-218
[PubMed]
[4] Beynon L.M., Richards J.C., Perry M.B.: The structure of the lipopolysaccharide O-antigen from Yersinia ruckeri serotype 01. Carbohydr. Res., 1994; 256: 303-317
[PubMed]
[5] Bhat U.R., Carlson R.W., Busch M., Mayer H.: Distribution and phylogenetic significance of 27-hydroxyoctacosanoic acid in lipopolysaccharides from bacteria belonging to the alpha-2 subgroup of Proteobacteria. Int. J. Syst. Bacteriol., 1991; 41: 213-217
[PubMed]
[6] Bhat U.R., Forsberg L.S., Carlson R.W.: Structure of lipid A component of Rhizobium leguminosarum bv. phaseoli lipopolysaccharide. Unique nonphosphorylated lipid A containing 2-amino-2-deoxygluconate, galacturonate, and glucosamine. J. Biol. Chem., 1994; 269: 14402-14410
[PubMed] [Full Text PDF]
[7] Bhat R., Marx A., Galanos C., Conrad R.S.: Structural studies of lipid A from Pseudomonas aeruginosa PAO1: occurrence of 4-amino-4-deoxyarabinose. J. Bacteriol., 1990; 172: 6631-6636
[PubMed] [Full Text HTML] [Full Text PDF]
[8] Bhat U.R., Mayer H., Yokota A., Hollingsworth R.I., Carlson R.W.: Occurrence of lipid A variants with 27-hydroxyoctacosanoic acid in lipopolysaccharides from members of the family Rhizobiaceae. J. Bacteriol., 1991; 173: 2155-2159
[PubMed] [Full Text HTML] [Full Text PDF]
[9] Boue S.M., Cole R.B.: Confirmation of the structure of lipid A from Enterobacter agglomerans by electrospray ionization tandem mass spectrometry. J. Mass Spectrom., 2000; 35: 361-368
[PubMed]
[10] Brade H., Rietschel E.T.: Identification of 2-keto-3-deoxy-1,7-dicarboxyheptonic acid as constituent of the lipopolysaccharide of Acinetobacter calcoaceticus NCTC 10305. Eur. J. Biochem., 1985; 153: 249-254
[PubMed]
[11] Brandenburg K.: Fourier transform infrared spectroscopy characterization of the lamellar and nonlamellar structures of free lipid A and Re lipopolysaccharides from Salmonella minnesota and Escherichia coli, J. Biophys., 1993; 64: 1215-1231
[PubMed] [Full Text HTML] [Full Text PDF]
[12] Brandenburg K., Mayer H., Koch M.H., Weckesser J., Rietschel E.T., Seydel U.: Influence of the supramolecular structure of free lipid A on its biological activity. Eur. J. Biochem., 1993; 218: 555-563
[PubMed]
[13] Broady K.W., Rietschel E.T., Lüderitz O.: The chemical structure of the lipid A component of lipopolysaccharides from Vibrio cholerae. Eur. J. Biochem., 1981; 115: 463-468
[PubMed]
[14] Caroff M., Deprun C., Richards J.C., Karibian D.: Structural characterization of the lipid A of Bordetella pertussis 1414 endotoxin. J. Bacteriol., 1994; 176: 5156-5159
[PubMed] [Full Text HTML] [Full Text PDF]
[15] Choma A., Sowinski P.: Characterization of Mesorhizobium huakuii lipid A containing both D-galacturonic acid and phosphate residues, Eur. J. Biochem., 2004; 271: 1310-1322
[PubMed] [Full Text HTML] [Full Text PDF]
[16] Corsaro M.M., Piaz F.D., Lanzetta R., Parrilli M.: Lipid A structure of Pseudoalteromonas haloplanktis TAC 125: use of electrospray ionization tandem mass spectrometry for the determination of fatty acid distribution. J. Mass Spectrom., 2002; 37: 481-488
[PubMed]
[17] Dalla Venezia N., Minka S., Bruneteau M., Mayer H., Michel G.: Lipopolysaccharides from Yersinia pestis. Studies on lipid A of lipopolysaccharides I and II. Eur. J. Biochem., 1985; 151: 399-404
[PubMed]
[18] Ferguson G.P., Datta A., Baumgartner J., Roop R.M., Carlson R.W., Walker G.C.: Similarity to peroxisomal-membrane protein family reveals that Sinorhizobium and Brucella BacA affect lipid-A fatty acids. Proc. Natl. Acad. Sci. USA., 2004; 101: 5012-5017
[PubMed] [Full Text HTML] [Full Text PDF]
[19] Fukuoka S., Kamishima H., Nagawa Y., Nakanishi H., Ishikawa K., Niwa Y., Tamiya E., Karube I.: Structural characterization of lipid A component of Erwinia carotovora lipopolysaccharide. Arch. Microbiol., 1992; 157: 311-318
[20] Fukuoka S., Knirel Y.A., Lindner B., Moll H., Seydel U., Zähringer U.: Elucidation of the structure of the core region and the complete structure of the R-type lipopolysaccharide of Erwinia carotovora FERM P-7576. Eur. J. Biochem., 1997; 250: 55-62
[PubMed]
[21] Galanos C., Lüderitz O., Westphal O.: A new method for the extraction of R lipopolysaccharides. Eur. J. Biochem., 1969; 9: 245-249
[PubMed]
[22] Gamian A., Katzenellenbogen E., Mieszała M., Romanowska E., Czarny A.: Studies on glycine as a component of bacterial lipopolysaccharides. 12th International Symposium on Glycoconjugates, Glyco XII, Kraków, Poland. Glycoconjugate J., 1993; 10: Abstr. S19.7
[23] Gamian A., Mieszała M., Katzenellenbogen E., Czarny A., Żal T., Romanowska E.: The occurrence of glycine in bacterial lipopolysaccharides. FEMS Immunol. Med. Microbiol., 1996; 13: 261-268
[PubMed]
[24] Gass J., Strobl M., Loibner A., Kosma P., Zähringer U.: Synthesis of allyl O-[sodium(alpha-D-glycero-D-talo-2-octulopyranosyl)onate]-(2–>6)-2-acetamido-2-deoxy-beta-D-glucopyranoside, a core constituent of the lipopolysaccharide from Acinetobacter calcoaceticus NCTC 10305. Carbohydr. Res., 1993; 244: 69-84
[PubMed]
[25] Gil-Serrano A.M., Gonzalez-Jimenez I., Tejero-Mateo P., Megias M., Romeo-Vazquez M.J.: Analysis of the lipid moiety of lipopolysaccharide from Rhizobium tropici CIAT899: identification of 29-hydroxytriacontanoic acid. J. Bacteriol., 1994; 176: 2454-2457
[PubMed] [Full Text HTML] [Full Text PDF]
[26] Hase S., Reitschel E.T.: The chemical structure of the lipid A component of lipopolysaccharides from Chromobacterium violaceum NCTC 9694. Eur. J. Biochem., 1977; 75: 23-34
[PubMed]
[27] Heine H., Müller-Loennies S., Brade L., Lindner B., Brade H.: Endotoxic activity and chemical structure of lipopolysaccharides from Chlamydia trachomatis serotypes E and L2 and Chlamydophila psittaci 6BC. Eur. J. Biochem., 2003; 270: 440-450
[PubMed] [Full Text HTML] [Full Text PDF]
[28] Helander I.M., Lindner B., Brade H., Altmann K., Lindberg A.A., Rietschel E.T., Zähringer U.: Chemical structure of the lipopolysaccharide of Haemophilus influenzae strain I-69 Rd-/b+. Description of a novel deep-rough chemotype. Eur. J. Biochem., 1988; 177: 483-492
[PubMed]
[29] Helander I.M., Kato Y., Kilpelainen I., Kostiainen R., Lindner B., Nummila K., Sugiyama T., Yokochi T.: Characterization of lipopolysaccharides of polymyxin-resistant and polymyxin-sensitive Klebsiella pneumoniae O3. Eur. J. Biochem., 1996; 237: 272-278
[PubMed]
[30] Helander I.M., Kilpelainen I., Vaara M., Moran A.P., Lindner B., Seydel U.: Chemical structure of the lipid A component of lipopolysaccharides of the genus Pectinatus. Eur. J. Biochem., 1994; 224: 63-70
[PubMed]
[31] Holst O.: Chemical structure of the core region of lipopolysaccharides. W: Endotoxin in health and disease. red.: H. Brade, S.M. Opal, S.N. Vogel, D.C. Morrison, Marcel Dekker Inc., New York – Basel 1999, 115-154
[32] Holst O., Borowiak D., Weckesser J., Mayer H.: Structural studies on the phosphate-free lipid A of Rhodomicrobium vannielii ATCC 17100. Eur. J. Biochem., 1983; 137: 325-332
[PubMed]
[33] Holst O., Brade H.: Chemical structure of the core region of lipopolysaccharides. W: Bacterial endotoxic lipopolysaccharides. red.: D. C. Morrison, J. L. Ryan. CRC Press, Boca Raton 1992, 134-170
[34] Holst O., Ulmer A., Brade H., Flad H.D., Rietschel E.T.: Biochemistry and cell biology of bacterial endotoxins. EMS Immunol. Med. Microbiol., 1996; 16: 83-104
[PubMed]
[35] Iida T., Haishima Y., Tanaka A., Nishiyama K., Saito S., Tanamoto K.: Chemical structure of lipid A isolated from Comamonas testosteroni lipopolysaccharide. Eur. J. Biochem., 1996; 237: 468-475
[PubMed]
[36] Imoto M., Kusumoto S., Shiba T., Naoki H., Iwashita T., Rietschel E. Th., Wollenweber H. W., Galanos C., Lüderitz O.: Chemical structure of E. coli lipid A: linkage site of acyl groups in the disaccharide backbone. Tetrahedron Lett., 1983; 24: 4017-4020
[37] Isshiki Y., Zähringer U., Kawahara K.: Structure of the core-oligosaccharide with a characteristic D-glycero-alpha-D-talo-oct-2-ulosylonate-(2–>4)-3-deoxy-D-manno-oct-2-ulosonate [alpha-Ko-(2–>4)-Kdo] disaccharide in the lipopolysaccharide from Burkholderia cepacia. Carbohydr. Res., 2003; 338: 2659-2666
[PubMed]
[38] Kaca W., Brade L., Rietschel E.T., Brade H.: The effect of removal of D-fructose on the antigenicity of the lipopolysaccharide from a rough mutant of Vibrio cholerae. Carbohydr. Res., 1986; 149: 293-298
[PubMed]
[39] Kasai N., Arata S., Mashimo J., Ogmori M., Mizutanii T., Egawa K.: Structure-activity relationships of endotoxic lipid A containing 2,3-diamino-2,3-dideoksy-D-glucose. Endotoxin Research Series, Elsevier, Amsterdam 1990, 1, 121
[40] Kaszowska M.: Budowa chemiczna i biosynteza lipopolisacharydu – ważnego składnika osłony komórkowej bakterii Gram-ujemnych. Post. Hig. Med. Dośw., 2004; 58: 333-334
[PubMed] [Full Text PDF]
[41] Kato H., Haishima Y., Iida T., Tanaka A., Tanamoto K.: Chemical structure of lipid A isolated from Flavobacterium meningosepticum lipopolysaccharide. J. Bacteriol., 1998; 180: 3891-3899
[PubMed] [Full Text HTML] [Full Text PDF]
[42] Katzenellenbogen E., Romanowska E., Shashkov A., Kocharova N., Knirel Y., Kochetkov N.: The structure of the O-specific polysaccharide of Hafnia alvei strain 1216. Carbohydr. Res., 1994; 259: 67-76
[PubMed]
[43] Kawahara K., Tsukano H., Watanabe H., Lindner B., Matsuura M.: Modification of the structure and activity of lipid A in Yersinia pestis lipopolysaccharide by growth temperature. Infect. Immun., 2002; 70: 4092-4098
[PubMed] [Full Text HTML] [Full Text PDF]
[44] Knirel Y., Paramonov N., Vinogradov E., Kochetkov N., Sidorczyk Z., Zych P.: 2-Acetamido-4-O-[(S)-1-carboxyethyl]-2-deoxy-D-glucose: a new natural isomer of N-acetylmuramic acid from the O-specific polysaccharyde of Proteus penneri 35. Carbohydr. Res., 1994; 259: C1-C3
[PubMed]
[45] Knirel Y.A., Paramonov N.A., Vinogradov E.V., Shashkov A.S., Kochetkov N.K., Sidorczyk Z., Świerzko A.: Structure of the O-specific polysaccharide of Proteus penneri 62 containing 2-acetamid-3-O-[(S)-1-carboxyethyl]-2-deoxy-D-glucose (N-acetylisomuramic acid). Carbohydr. Res., 1992; 235: C19-C23
[PubMed]
[46] Kondo S., Zähringer U., Seydel U., Sinwell V., Hisatsune K., Rietschel E.T.: Chemical structure of the carbohydrate backbone of Vibrio parahaemoliticus serotype 012 lipopolisaccharide. Eur. J. Biochem., 1991; 200: 689-698
[PubMed]
[47] Krasikova I.N., Kapustina N.V., Isakov V.V., Dmitrenok A.S., Dmitrenok P.S., Gorshkova N.M., Solov’eva T.F.: Detailed structure of lipid A isolated from lipopolysaccharide from the marine proteobacterium Marinomonas vaga ATCC 27119. Eur. J. Biochem., 2004; 271: 2895-2904
[PubMed] [Full Text HTML] [Full Text PDF]
[48] Krauss J.H., Seydel U., Weckesser J., Mayer H.: Structural analysis of the nontoxic lipid A of Rhodobacter capsulatus 37b4. Eur. J. Biochem., 1989; 180: 519-526
[PubMed]
[49] Kulshin V.A., Zahringer U., Lindner B., Frasch C.E., Tsai C.M., Dmitriev B.A., Rietschel E.T.: Structural characterization of the lipid A component of pathogenic Neisseria meningitidis. J. Bacteriol.,1992; 174: 1793-1800
[PubMed] [Full Text HTML] [Full Text PDF]
[50] Kulshin V.A., Zähringer U., Lindner B., Jäger K.E., Dmitriev B.A., Rietschel E.T.: Structural characterization of the lipid A component of Pseudomonas aeruginosa wild-type and rough mutant lipopolysaccharides. Eur. J. Biochem., 1991; 198: 697-704
[PubMed]
[51] Kumada H., Haishima Y., Umemoto T., Tanamoto K.: Structural study on the free lipid A isolated from lipopolysaccharide of Porphyromonas gingivalis. J. Bacteriol., 1995; 177: 2098-2106
[PubMed] [Full Text HTML] [Full Text PDF]
[52] Kunicki-Goldfinger W.: Życie bakterii. Mikroskopijne zakłady energetyczne. Wydawnictwo Naukowe PWN, Warszawa 2001
[53] Ługowski C., Niedziela T., Jachymek W.: Endotoksyny bakteryjne: struktura, aktywności biologiczne, szczepionki koniugatowe. Mikrobiol. Med., 1996; 4: 28-39
[54] Ługowski C., Romanowska E.: Chemical studies on Shigella sonnei lipid A. Eur. J. Biochem., 1974; 48: 319-323
[PubMed]
[55] Łukasiewicz J., Ługowski C.: Biologiczna aktywność lipopolisacharydu. Post. Hig. Med. Dośw., 2003; 57: 33-36
[PubMed]
[56] Łukasiewicz J., Niedziela T., Jachymek W., Kenne L., Ługowski C.: Structure of the lipid A-inner core region and biological activity of Plesiomonas shigelloides O54 (strain CNCTC 113/92) lipopolysaccharide. Glycobiology, 2006; 16: 538-550
[PubMed]
[57] Markiewicz Z.: Struktura i funkcje osłon bakteryjnych. Wydawnictwo Naukowe PWN, Warszawa 1993
[58] Masoud H., Perry M., Richards J.: Characterization of the lipopolysaccharide of Moraxella catarrhalis. Structural analysis of the lipid A from M. catarrhalis serotype A lipopolysaccharide. Eur. J. Biochem., 1994; 220: 209-216
[PubMed]
[59] Masoud H., Weintraub S.T., Wang R., Cotter R., Holt S.C.: Investigation of the structure of lipid A from Acinobacillus actinomycetemcomitians strain Y4 and human clinical isolate PO 1021-7. Eur. J. Biochem., 1991; 200: 775-781
[PubMed]
[60] Mayer H., Salimath P.V., Holst O., Weckesser J.: Unusual lipid A types in phototrophic bacteria and related species. Rev. Infect. Dis., 1984; 6: 542-545
[PubMed]
[61] Melaugh W., Phillips N.J., Campagnari A.A., Karalus R., Gibson B.W.: Partial characterization of the major lipooligosaccharide from a strain of Haemophilus ducreyi, the causative agent of chancroid, a genital ulcer disease. J. Biol. Chem., 1992; 267: 13434-13439
[PubMed] [Full Text PDF]
[62] Moran A.P., Lindner B., Walsh E.J.: Structural characterization of the lipid A component of Helicobacter pylori rough- and smooth-form lipopolysaccharides. J. Bacteriol., 1997; 179: 6453-6463
[PubMed] [Full Text PDF]
[63] Moran A.P., Zähringer U., Seydel U., Scholz D., Stutz P., Rietschel E.T.: Structural analysis of the lipid A component of Campylobacter jejuni CCUC 10936 (serotype O:2) lipopolysaccharide. Description of a lipid A containing a hybrid backbone of 2-amino-2-deoxy-D-glucose and 2,3-diamino-2,3-dideoxy-D-glucose. Eur. J. Biochem., 1991; 198: 459-469
[PubMed]
[64] Niedziela T., Dag S., Łukasiewicz J., Dzieciatkowska M., Jachymek W., Ługowski C., Kenne L.: Complete lipopolysaccharide of Plesiomonas shigelloides O74: H5 (strain CNCTC 144/92). 1. Structural analysis of the highly hydrophobic lipopolysaccharide, including the O-antigen, its biological repeating unit, the core oligosaccharide and the linkage between them. Biochemistry, 2006; 45: 10422-10433
[PubMed]
[65] Ogawa T.: Chemical structure of lipid A Porphyromonas (Bacteroides) gingivalis lipopolysaccharide. FEBS Lett., 1993; 332: 197-201
[PubMed]
[66] Pietras T., Mazerat P.: Udział endotoksyny bakterii Gram-ujemnych w patogenezie chorób człowieka. Mikrobiol. Med., 1998; 1: 7-11
[67] Plötz B.M., Lindner B., Stetter K.O., Holst O.: Characterization of a novel lipid A containing D-galacturonic acid that replaces phosphate residues. The structure of the lipid A of the lipopolysaccharide from the hyperthermophilic bacterium Aquifex pyrophilus. J. Biol. Chem., 2000; 275: 11222-11228
[PubMed] [Full Text HTML] [Full Text PDF]
[68] Post D.M., Phillips N.J., Shao J.Q., Entz D.D., Gibson B.W., Apicella M.A.: Intracellular survival of Neisseria gonorrhoeae in male urethral epithelial cells: importance of a hexaacyl lipid A. Infect. Immun., 2002; 70: 909-920
[PubMed] [Full Text HTML] [Full Text PDF]
[69] Que N.L., Lin S., Cotter R.J., Raetz C.R.: Purification and mass spectrometry of six lipid A species from the bacterial endosymbiont Rhizobium etli. Demonstration of a conserved distal unit and a variable proximal portion. J. Biol. Chem., 2000; 275: 28006-28016
[PubMed] [Full Text HTML] [Full Text PDF]
[70] Que-Gewirth N.L., Ribeiro A.A., Kalb S.R., Cotter R.J., Bulach D.M., Adler B., Girons I.S., Werts C., Raetz C.R.: A methylated phosphate group and four amide-linked acyl chains in Leptospira interrogans lipid A. The membrane anchor of an unusual lipopolysaccharide that activates TLR2. J. Biol. Chem., 2004; 279: 25420-25429
[PubMed] [Full Text HTML] [Full Text PDF]
[71] Qureshi N., Honovich J.P., Hara H., Cotter R.J., Takayama K.: Location of fatty acids in lipid A obtained from lipopolysaccharide of Rhodopseudomonas sphaeroides ATCC 17023. J. Biol. Chem., 1988; 236: 5502-5504
[PubMed] [Full Text PDF]
[72] Qureshi N., Mascagni P., Ribi E., Takayama K.: Monophosphoryl lipid A obtained from lipopolysaccharides of Salmonella minnesota R595. Purification of the dimethyl derivative by high performance liquid chromatography and complete structural determination. J. Biol. Chem., 1985; 260: 5271-5278
[PubMed] [Full Text PDF]
[73] Qureshi N., Takayama K., Meyer K.C., Kirkland T.N., Bush C.A., Chen L., Wang R., Cotter R.J.: Chemical reduction of 3-oxo and unsaturated groups in fatty acid of diphosphoryl lipid A from the lipopolysaccharide of Rhodopseudomonas sphaeroides. Comparison of biological properties before and after reduction. J. Biol. Chem., 1991; 266: 6532-6538
[PubMed] [Full Text PDF]
[74] Qureshi N., Takayama K., Ribi E.: Purification and structural determination of nontoxic lipid A obtained from the lipopolysaccharide of Salmonella typhimurium. J. Biol. Chem., 1982; 257: 11808-11815
[PubMed] [Full Text PDF]
[75] Rau H., Seydel U., Freudenberg M., Weckesser J., Mayer H.: Lipopolysaccharide of Rhodospirillum salinarum 40: structural studies on the core and lipid A region. Arch. Microbiol., 1995; 164: 280-289
[PubMed]
[76] Rietschel E.T., Brade H.: Endotoksyny bakteryjne. Świat Nauki, 1992; 10: 64-73
[PubMed]
[77] Rietschel E.T., Krikae T., Schade F.U., Mamat U., Schmidt G., Loppnow H., Ulmer A.J., Zähringer U., Seydel U., Di Padova F., Schreier M., Brade H.: Bacterial endotoxin: molecular relationships of structure to activity and function. FASEB J., 1994; 8: 217-225
[PubMed] [Full Text PDF]
[78] Rosner M.R., Tang J., Barzilay I., Khorana H.G.: Structure of the lipopolysaccharide from an Escherichia coli heptose-less mutant. I. Chemical degradations and identification of products. J. Biol. Chem., 1979; 254: 5906-5917
[PubMed] [Full Text PDF]
[79] Różalski A.: Lipopolisacharyd (LPS) bakterii Gram-ujemnych – struktura chemiczna, aktywność biologiczna i znaczenie w chorobotwórczości. [I]: Struktura chemiczna i właściwości fizyko-chemiczne lipopolisacharydów. Post. Mikrobiol., 1995; 34: 289-313
[80] Różalski A.: Lipopolisacharyd (LPS) bakterii Gram-ujemnych – struktura chemiczna, aktywność biologiczna i znaczenie w chorobotwórczości. [II]: Budowa chemiczna a funkcja biologiczna LPS. Post. Mikrobiol., 1995; 34: 317-337
[81] Różalski A.: Lipopolisacharyd (LPS) bakterii Gram-ujemnych – struktura chemiczna, aktywność biologiczna i znaczenie w chorobotwórczości. [III]: Lipopolisacharyd jako czynnik chorobotwórczości. Post. Mikrobiol., 1995; 34: 339-364
[82] Rund S., Lindner B., Brade H., Holst O.: Structural analysis of the lipopolysaccharide from Chlamydia trachomatis serotype L2. J. Biol. Chem., 1999; 274: 16819-16824
[PubMed] [Full Text HTML] [Full Text PDF]
[83] Salimath P.V., Tharanathan R.N., Weckesser J., Mayer H.: The structure of the polysaccharide moiety of Rhodopseudomonas sphaeroides ATCC 17023 lipopolysaccharide. Eur. J. Biochem., 1984; 144: 227-232
[PubMed]
[84] Salimath P.V., Weckesser J, Strittmatter W., Mayer H.: Structural studies on the non-toxic lipid A from Rhodopseudomonas sphaeroides ATCC 17023. Eur. J. Biochem., 1983; 136: 195-200
[PubMed]
[85] Scheidle H., Gross A., Niehaus K.: The Lipid A substructure of the Sinorhizobium meliloti lipopolysaccharides is sufficient to suppress the oxidative burst in host plants. New Phytol., 2005; 165: 559-565
[PubMed]
[86] Schromm A.B., Brandenburg K., Loppnow H., Moran A.P., Koch M.H., Rietschel E.T., Seydel U.: Biological activities of lipopolysaccharides are determined by the shape of their lipid A portion. Eur. J. Biochem., 2000; 267: 2008-2013
[PubMed] [Full Text HTML] [Full Text PDF]
[87] Schwudke D., Linscheid M., Strauch E., Appel B., Zähringer U., Moll H., Muller M., Brecker L., Gronow S., Lindner B.: The obligate predatory Bdellovibrio bacteriovorus possesses a neutral lipid A containing a
-D-mannoses that replace phosphate residues: similarities and differences between the lipid As and the lipopolysaccharides of the wild type strain B. bacteriovorus HD100 and its host-independent derivative HI100. J. Biol. Chem., 2003; 278: 27502-27512
[PubMed] [Full Text HTML] [Full Text PDF]
[88] Seydel U., Lindner B., Wollenweber H.W., Rietschel E.T.: Structural studies on the lipid A component of enterobacterial lipopolysaccharides by laser desorption mass spectrometry. Location of acyl groups at the lipid A backbone. Eur. J. Biochem., 1984; 145: 505-509
[PubMed]
[89] Sidorczyk Z., Zähringer U., Rietschel E.T.: Chemical structure of the lipid A component of the lipopolysaccharide from a Proteus mirabilis Re-mutant. Eur. J. Biochem., 1983; 137: 15-22
[PubMed]
[90] Silipo A., De Castro C., Lanzetta R., Molinaro A., Parrilli M.: Full structural characterization of the lipid A components from the Agrobacterium tumefaciens strain C58 lipopolysaccharide fraction. Glycobiology, 2004; 14: 805-815
[PubMed] [Full Text HTML] [Full Text PDF]
[91] Silipo A., Lanzetta R., Amoresano A., Parrilli M., Molinaro A.: Ammonium hydroxide hydrolysis: a valuable support in the MALDI-TOF mass spectrometry analysis of Lipid A fatty acid distribution. J. Lipid Res., 2002; 43: 2188-2195
[PubMed] [Full Text HTML] [Full Text PDF]
[92] Silipo A., Lanzetta R., Garozzo D., Lo Cantore P., Iacobellis N.S., Molinaro A., Parrilli M., Evidente A.: Structural determination of lipid A of the lipopolysaccharide from Pseudomonas reactans. A pathogen of cultivated mushrooms. Eur. J. Biochem., 2002; 269: 2498-2505
[PubMed] [Full Text HTML] [Full Text PDF]
[93] Sonesson A., Jantzen E., Tangen T., Zähringer U.: Chemical characterization of lipopolysaccharides from Legionella feeleii, Legionella hackeliae and Legionella jordanis. Microbiol., 1994; 140: 2663-2671
[PubMed]
[94] Sonesson A., Jantzen E., Tangen T., Zähringer U.: Chemical composition of lipopolysaccharides from Legionella bozemanii and Legionella longbeachae. Arch. Microbiol., 1994; 162: 215-221
[PubMed]
[95] Strittmatter W., Weckesser J., Salimath P.V., Galanos C.: Nontoxic lipopolysaccharide from Rhodopseudomonas sphaeroides ATCC 17023. J. Bacteriol., 1983; 155: 153-158
[PubMed] [Full Text HTML] [Full Text PDF]
[96] Takayama K., Qureshi N., Hyver K., Honovich J., Cotter R.J., Mascagni P., Schneider H.: Characterization of a structural series of lipid A obtained from the lipopolysaccharides of Neisseria gonorrhoeae. Combined laser desorption and fast atom bombardment mass spectral analysis of high performance liquid chromatography-purified dimethyl derivatives. J. Biol. Chem., 1986; 261: 10624-10631
[PubMed] [Full Text PDF]
[97] Tanamoto K.: Free hydroxyl groups are not required for endotoxic activity of lipid A. Infect. Immun., 1994; 62: 1705-1709
[PubMed] [Full Text HTML] [Full Text PDF]
[98] Tanamoto K., Azumi S., Haishima Y., Kumada H., Umemoto T.: Endotoxic properties of free lipid A from Porphyromonas gingivalis. Microbiology, 1997; 143: 63-71
[PubMed] [Full Text PDF]
[99] Tanamoto K., Iida T., Haishima Y., Azumi S.: Endotoxic properties of lipid A from Comamonas testosteroni. Microbiology, 2001; 147: 1087-1094
[PubMed] [Full Text HTML] [Full Text PDF]
[100] Tanamoto K., Kato H., Haishima Y., Azumi S.: Biological properties of lipid A isolated from Flavobacterium meningosepticum. Clin. Diagn. Lab. Immunol., 2001; 8: 522-527
[PubMed] [Full Text HTML] [Full Text PDF]
[101] Tharanathan R.N., Weckesser J., Mayer H.: Structural studies on the D-arabinose-containing lipid A from Rhodospirillum tenue 2761. Eur. J. Biochem., 1978; 84: 385-394
[PubMed]
[102] Toman R., Garidel P., Andra J., Slaba K., Hussein A., Koch M.H., Brandenburg K.: Physicochemical characterization of the endotoxins from Coxiella burnetii strain Priscilla in relation to their bioactivities. BMC Biochem., 2004; 5: 1-11
[PubMed] [Full Text HTML] [Full Text PDF]
[103] Trent M.S.: Biosynthesis, transport, and modification of lipid A. Biochem. Cell Biol., 2004; 82: 71-86
[PubMed]
[104] Tsukioka D., Nishizawa T., Miyase T., Achiwa K., Suda T., Soma G., Mizuno D.: Structural characterization of lipid A obtained from Pantoea agglomerans lipopolysaccharide. FEMS Microbiol Lett., 1997; 149: 239-244
[PubMed]
[105] Vinogradov E.V., Kaca W., Shashkov A.S., Krajewska-Pietrasik D., Rozalski A., Knirel Y.A., Kochetkov N.K.: The structure of Proteus mirabilis O3 O-specific polysaccharide containing N-(2-hydroxyethyl)-D-alanine. Eur. J. Biochem., 1990; 188: 645-651
[PubMed]
[106] Vinogradov E., Perry M.B., Conlan J.W.: Structural analysis of Francisella tularensis lipopolysaccharide. Eur. J. Biochem., 2002; 269: 6112-6118
[PubMed] [Full Text HTML] [Full Text PDF]
[107] Weckesser J., Mayer H.: Different lipid A types in lipopolysaccharides of phototrophic and related non-phototrophic bacteria. FEMS Microbiol. Rev., 1988; 54: 143-153
[PubMed]
[108] Weintraub A., Zähringer U., Wollenweber H.W., Seydel U., Rietschel E.T.: Structural characterization of the lipid A component of Bacteroides fragilis strain NCTC 9343. Eur. J. Biochem., 1989; 183: 425-431
[PubMed]
[109] Weisshaar R., Lingens F.: The lipopolysaccharide of a chloridazon-degrading bacterium. Eur. J. Biochem., 1983; 137: 155-161
[PubMed]
[110] Westphal O., Lüderitz O. Bister F.: Über die extraction von bacterien mit phenol-wasser. Z. Naturforsch., 1952; 78: 148-155
[111] Wollenweber H.W., Seydel U., Lindner B., Lüderitz O., Rietschel E.T.: Nature and location of amide-bound (R)-3-acyloxyacyl groups in lipid A of lipopolysaccharides from various Gram-negative bacteria. Eur. J. Biochem., 1984; 145: 265-272
[PubMed]
[112] Zähringer U., Knirel Y.A., Lindner B., Helbig J.H., Sonesson A., Marre R., Rietschel E.T.: The lipopolysaccharide of Legionella pneumophila serogroup 1 (strain Philadelphia 1): chemical structure and biological significance. Prog. Clin. Biol. Res., 1995; 392: 113-139
[PubMed]
[113] Zähringer U., Lindner B., Knirel Y.A., van den Akker W.M., Hiestand R., Heine H., Dehio C.: Structure and biological activity of the short-chain lipopolysaccharide from Bartonella henselae ATCC 49882 T. J. Biol. Chem., 2004; 279: 21046-21054
[PubMed] [Full Text HTML] [Full Text PDF]
[114] Zähringer U., Lindner B., Rietschel E.T.: Chemical structure of lipid A: recent advances in structural analysis of biologically active molecules. W: Endotoxin in Health and Disease, red.: D. Morrison, H. Brade, S. Opal, S. Vogel. Marcel Dekker, New York 1998, 93
[115] Zielińska-Kuźniarz K., Mieszała M., Lipiński T., Gamian A.: Glicyna jako integralny składnik bakteryjnych lipopolisacharydów. Post. Hig. Med. Dośw., 2003; 57: 473-483
[PubMed]