
Pathogenesis and prophylaxis of AMD: focus on oxidative stress and antioxidants
Anna Wiktorowska-Owczarek 1 , Jerzy Nowak 1Streszczenie
Zwyrodnienie plamki związane z wiekiem (AMD) jest przyczyną poważnych ubytków widzenia centralnego ze ślepotą włącznie u osób powyżej 55–60 r.ż. Patogeneza schorzenia, wciąż słabo poznana, wynika z nałożenia interakcji metaboliczno-funkcjonalno-genetycznych oraz czynników środowiskowych. Ważnym ogniwem w patogenezie AMD jest stres oksydacyjny w siatkówce – strukturze, która jest szczególnie wrażliwa na działanie reaktywnych form tlenu (RFT), ponieważ błona zewnętrznych segmentów fotoreceptorów zawiera wielonienasycone kwasy tłuszczowe, które łatwo ulegają utlenianiu, co inicjuje cytotoksyczną reakcję łańcuchową. Występujące w sąsiedztwie fotoreceptorów komórki nabłonka barwnikowego siatkówki (RPE) biorą czynny udział zarówno w cyklu retinoidów, jak i metabolizmie nieustannie zużywających się w procesie widzenia segmentów zewnętrznych fotoreceptorów. Fagocytowane przez RPE fragmenty fotoreceptorów nie są w pełni degradowane, co przyczynia się do odkładania w lizosomach złogów nierozpuszczalnego materiału określanego jako lipofuscyna; jej składowymi są m.in. związki zawierające retinoidy o silnych właściwościach fotouwrażliwiających, przyczyniających się do nasilania i propagacji stresu oksydacyjnego. Siatkówka zawiera antyoksydacyjne systemy obronne – enzymatyczne i nieenzymatyczne, które w warunkach fizjologii skutecznie neutralizują skutki stresu oksydacyjnego. W obrębie systemu nieenzymatycznego szczególną rolę pełnią ksantofile (luteina i zeaksantyna), witaminy C i E oraz inne, których ilość w tkankach oka obniża się z wiekiem, stwarzając zagrożenie rozwoju patologii AMD. W pracy przedstawiono najnowsze koncepcje na temat roli stresu oksydacyjnego w patogenezie AMD, a także omówiono najważniejsze elementy antyoksydacyjnego systemu obronnego. Przedstawiono również przegląd preparatów handlowych zawierających barwniki plamkowe, które znajdują zastosowanie w profilaktyce i leczeniu AMD.
Słowa kluczowe:AMD • stres oksydacyjny • lipofuscyna • karotenoidy • luteina • zeaksantyna • siatkówkowe systemy antyoksydacyjne • profilaktyka
Summary
Age-related macular degeneration (AMD) is the leading cause of severe visual loss and blindness in people over 55. Its pathogenesis – likely multifactorial, involving a complex interaction of metabolic, functional, genetic and environmental factors – remains poorly understood. Among molecular links in pathogenesis of AMD is the oxidative stress in the retina, a structure that is particularly susceptible to damage by reactive oxygen species (ROS) since photoreceptor outer segment (POS) membranes are rich in polyunsaturated fatty acids which can be readily oxidized and can initiate a cytotoxic chain reaction. Occurring in the neighborhood of photoreceptors, the retinal pigment epithelial cells (RPE) actively contribute to both the retinoid cycle and catabolism of constantly shed and phagocytized parts of photoreceptor outer segments. Enzymatic degradation of photoreceptor fragments occurring in RPE phagolysosomes is not complete and undigested material in the form of insoluble aggregates, called lipofuscin, is deposited in lysosomes of RPE cells. Lipofuscin contains a mixture of diverse molecular components including retinoid-derived compounds, some of which displaying potent photoinducible properties, contributing to an enhancement and propagation of the oxidative stress. The retina possesses defense mechanisms against the oxidative stress that effectively neutralize the consequences of reactive oxygen species actions under normal conditions. A key role in the antioxidant defense plays an array of substances, including: xanthophylls (lutein and zeaxanthin), vitamin C and E, and glutathione. This paper surveys the current concepts on the role of the oxidative stress in pathophysiology of AMD, and describes major components of the antioxidant defense system, including their use in AMD prophylaxis and therapy.
Key words:AMD • oxidative stress • lipofuscin • carotenoids • vitamin C • vitamin E • glutathione • retinal antooxidative defense systems • prophylaxis
Wykaz skrótów:
AMD – zwyrodnienie plamki związane z wiekiem; ARA – kwas arachidonowy; CEP – karboksyetylopirol; DHA – kwas dokozaheksaenowy; DPA – kwas dokozapentaenowy; EPA – kwas eikozapentaenowy; HHE – 4-hydroksy-2-heksenal; HOHA – kwas 4-hydroksy-7-oksohepta-5-enowy; HNE – 4-hydroksy-2-nonenal; MDA – dialdehyd malonowy; PEDF – czynnik pochodzący z nabłonka barwnikowego; POS – zewnętrzne segmenty fotoreceptorów; RFT – reaktywne formy tlenu; RPE – nabłonek barwnikowy siatkówki; SOD – dysmutaza ponadtlenkowa; VEGF – czynnik wzrostu śródbłonka naczyń; WNKT – wielonienasycone kwasy tłuszczowe; WRT – wolne rodniki tlenowe.
1. Wstęp
Zwyrodnienie plamki związane z wiekiem jest postępującą chorobą plamki żółtej, której uszkodzenie doprowadza – w zależności od zaawansowania patologii – do pogorszenia lub różnego stopnia ubytków widzenia centralnego, z całkowitą utratą widzenia włącznie. W publikacjach anglojęzycznych, a także coraz częściej w języku obiegowym ta choroba jest określana jako AMD od age-related macular degeneration. Jak nazwa oraz badania epidemiologiczne przeprowadzone w krajach całego świata wskazują, że patologia ta koreluje z wiekiem i ujawnia się u ludzi zwykle po 55–60 roku życia. AMD częściej dotyka kobiety, rasę białą oraz osoby o niebieskiej barwie tęczówki. Proces chorobotwórczy rozwija się podstępnie, jest złożony i nie w pełni poznany. Podkreśla się rolę czynników środowiskowych i genetycznych, a także wiele anomalii metabolicznych i funkcjonalnych przebiegających w obrębie kompleksu anatomiczno- funkcjonalnego obejmującego fotoreceptory, nabłonek barwnikowy siatkówki (RPE), błonę Brucha i choriokapilary. Pełniejszy opis symptomatologii AMD zawarty jest w innych publikacjach obecnych autorów [33,37,39].
Wyróżnia się przynajmniej cztery główne czynniki/procesy leżące u podstaw zwyrodnienia plamki związanego z wiekiem:
• Lipofuscyna – powstająca w procesie lipofuscynogenezy w komórkach nabłonka barwnikowego siatkówki (retinal pigment epithelium – RPE). Tworzenie lipofuscyny ocznej ma związek z aktywnością wzrokową fotoreceptorów i ich funkcjonalnie niezbędnego partnera – komórek RPE. W procesie widzenia nieustannie zużywają się szczytowe fragmenty zewnętrznych segmentów fotoreceptorów (photoreceptor outer segment – POS), które – po oderwaniu się – są fagocytowane przez sąsiadujące z fotoreceptorami komórki RPE. Do fagolizosomów RPE dostają się zatem wszystkie składniki budulcowe i funkcjonalne obecne w POS, wśród nich pigmenty wzrokowe występujące w błonach dysków, lipidy błonowe, zawierające m.in. fragmenty i metabolity wielonienasyconych kwasów tłuszczowych (głównie kwasu dokozaheksaenowego, DHA), a także ich połączenia z białkami (lipopropeidy), reaktywne związki izolewuglandynowe (powstające z fosfolipidów błonowych zawierających kwas arachidonowy) oraz różne związki związane z transkomórkowym cyklem retinoidów kaskady wzbudzenia wzrokowego, w tym ich produkty uboczne o potencjale cytotoksycznym. W wyniku nieustannej podaży wspomnianego materiału i ograniczonej aktywności lizosomalnych enzymów degradujących, dochodzi do gromadzenia się we wnętrzu fagolizosomów nierozpuszczalnych złogów określanych jako lipofuscyna. Nasilający się wraz z wiekiem proces lipofuscynogenezy nie jest cechą charakterystyczną dla komórek RPE, zachodzi on bowiem w wielu typach komórek postmitotycznych (np. kardiomiocytach czy neuronach), dlatego też powstająca i kumulująca się lipofuscyna często nazywana jest barwnikiem/ pigmentem „starości”. Jednak lipofuscyna „oczna” różni się od innych pigmentów starości unikatowym składem, w których dominują fotowrażliwe i cytotoksyczne składniki, np. wysoce fotocytotoksyczny bis-retinoid pirymidoniowy, zawierający dwie czasteczki trans-retinalu i znany pod skrótową nazwą A2E, czy immunogenne addukty fragmentów DHA z albuminami, łatwo indukujące reakcje z pogranicza patologii bądź wręcz reakcje patologiczne [37,39]. Należy dodać, że przeładowane lipofuscyną fagolizosomy tracą z czasem szczelność i zawarty w nich materiał może wyciekać do cytoplazmy i dalej – z funkcjonalnie „niedomagających” komórek RPE – na zewnątrz w kierunku błony Brucha, stanowiąc zarzewie druzów [33,34,36,37,39].
• Druzy czyli nierozpuszczalne złogi, które powstają między warstwą komórek nabłonka barwnikowego (RPE) a błoną Brucha w procesie określanym jako druzogeneza. Badania proteomiczne materiału druzowego wykazały obecność mieszaniny różnorodnych białek, w tym białek o typowej lokalizacji zarówno wewnątrz- jak i zewnątrzkomórkowej, białek osocza oraz białek związanych z aktywnością układu odpornościowego (głównie układu dopełniacza, włącznie z kompleksem atakującym błonę – MAC), a także immunogennych adduktów lipoproteinowych [33,34,36,37,39].
• Zapalenie mające związek z przewlekłym procesem zapalnym o zasięgu lokalnym bez objawów (a w wywiadzie bez epizodów wskazujących na przebycie) klasycznego zapalenia, ale przypominające w pewnych aspektach zapalenie. Proces taki obecnie określa się terminem parainflammation w rozumieniu tkankowej odpowiedzi adaptacyjnej na czynniki stresowe (szkodliwe) lub funkcjonalne dysfunkcje (mulfunctions), które mogą być przyczyną rozwinięcia i/lub podtrzymania („tlenia się”) procesu patologicznego1. W przypadku AMD takimi przewlekłymi i nasilającymi się z wiekiem „stresorami” mogą być: lipofuscyna i druzy (bądź ich składniki) – produkty dwóch procesów: lipofuscynogenezy i druzogenezy, które zachodzą u wszystkich ludzi, jednakże tylko u niektórych doprowadzają do rozwinięcia się jawnej patologii. Wystąpienie tej ostatniej może się wiązać z tym, że stan zagrożenia bądź „tlącej” się dysfunkcji może być rozpoznany przez mechanizmy (sensory) nieswoiste (wrodzone) układu odpornościowego, którego celem jest uruchomienie działań naprawczych, poprzez m.in. uruchomienie „programu zapalenie”, zmierzających do przywrócenia homeostazy [33,34,36,37,39].
• Neowaskularyzacja (w przypadku postaci wysiękowej AMD) jest skutkiem lokalnej czynnościowej przewagi czynników proangiogennych (głównie VEGF – vascular endothelial growth factor – czynnik wzrostu śródbłonka naczyń) nad czynnikami antyangiogennymi (np. PEDF – pigment epithelium-derived factor – czynnik pochodzący z nabłonka barwnikowego). Nowo tworzone naczynia pochodzą z włośniczek naczyniówki [33,37,39,40].
Wyróżnia się dwie postacie kliniczne AMD: suchą (lub zanikową), w której obserwuje się postępujący zanik komórek nabłonka barwnikowego siatkówki (RPE) i wtórnie fotoreceptorów, jako konsekwencję (w dużej mierze) procesu druzogenezy oraz postać mokrą (inaczej określaną jako wysiękową lub neowaskularną), a związaną z tworzeniem naczyń tuż pod siatkówką w obszarze plamki, gdzie naturalnie one nie występują. Ponadto nowo powstałe naczynia są zazwyczaj kruche, nieszczelne, co prowadzi do wysięków i krwotoków [40].
Oprócz wyżej wymienionych czynników, wymienia się również rolę stresu oksydacyjnego w patomechanizmie AMD, głównie w kontekście lipofuscynogenezy. Stres oksydacyjny wynika z zaburzenia równowagi między wytwarzanymi reaktywnymi rodnikami a zdolnością obronną organizmu, a więc zdolnością do ich neutralizacji. Rozchwianie homeostazy jest pochodną nadmiernego wytwarzania wolnych rodników lub dysfunkcji układu antyoksydacyjnego w warunkach ich naturalnego wytwarzania.
W niniejszym artykule zostaną przedstawione przyczyny nadmiernego powstawania wolnych rodników właśnie w siatkówce, a tym samym ich udział w patogenezie AMD.
Omówimy ponadto systemy obrony antyoksydacyjnej, a także przedstawimy zawierające barwniki plamkowe preparaty handlowe rekomendowane w profilaktyce i terapii AMD.
2. Powstawanie reaktywnych form tlenu (RFTRFTRFT) w siatkówce
Powstawanie i neutralizacja wolnych rodników, w tym tlenowych, są procesami powszechnie występującymi w komórkach. Procesy te są nieodłącznymi elementami metabolizmu komórkowego i nie stanowią zagrożenia w sytuacji ich szybkiej inaktywacji przez obecne w komórkach systemy obrony o charakterze antyoksydacyjnym. Wśród wolnych rodników, charakteryzujących się jednym niesparowanym elektronem na ostatniej orbicie, ważną grupę stanowią wolne rodniki tlenowe (WRT). Wolne rodniki tlenowe, do których zalicza się anionorodnik ponadtlenkowy (O2–•) i rodnik hydroksylowy (OH•), należą do szerszej grupy wysoce reaktywnych związków, tzw. reaktywnych form tlenu (RFT; reactive oxygen species – ROS). Przykładem takiej reaktywnej cząsteczki jest nadtlenek wodoru (H2O2), łatwo ulegający rozpadowi np. do rodnika hydroksylowego [4,56]. W komórce takie formy tlenu powstają przede wszystkim w mitochondrium, co ma znaczenie w procesie starzenia, a także w siateczce endoplazmatycznej, błonie komórkowej, peroksysomach i innych miejscach, określanych jako pozamitochondrialne źródła RFT, które z kolei odgrywają rolę w chorobach związanych z wiekiem [41]. Anionorodnik ponadtlenkowy powstaje głównie na łańcuchu oddechowym w reakcji fosforylacji oksydacyjnej, na skutek jednoelektronowej reakcji nieenzymatycznej z tlenem i jest prekursorem dla kolejnych reaktywnych form tlenu. Podczas jego inaktywacji z udziałem dysmutazy ponadtlenkowej (SOD) powstaje nadtlenek wodoru, który z kolei w reakcjach Fentona:

i Habera-Weissa (katalizatorem jest miedź):

zostaje przekształcony do najbardziej reaktywnego wolnego rodnika hydroksylowego (OH•); wyżej opisane tworzenie RFT i ich dezaktywacja przez enzymatyczny układ antyoksydacyjny przedstawiono na ryc. 1. Powstające w komórkach np. nabłonka barwnikowego siatkówki (RPE), opisane wyżej reaktywne formy tlenu dają początek następnym rodnikom, co może prowadzić do dysfunkcji, a w dalszej konsekwencji degeneracji i/lub atrofii komórek [19].
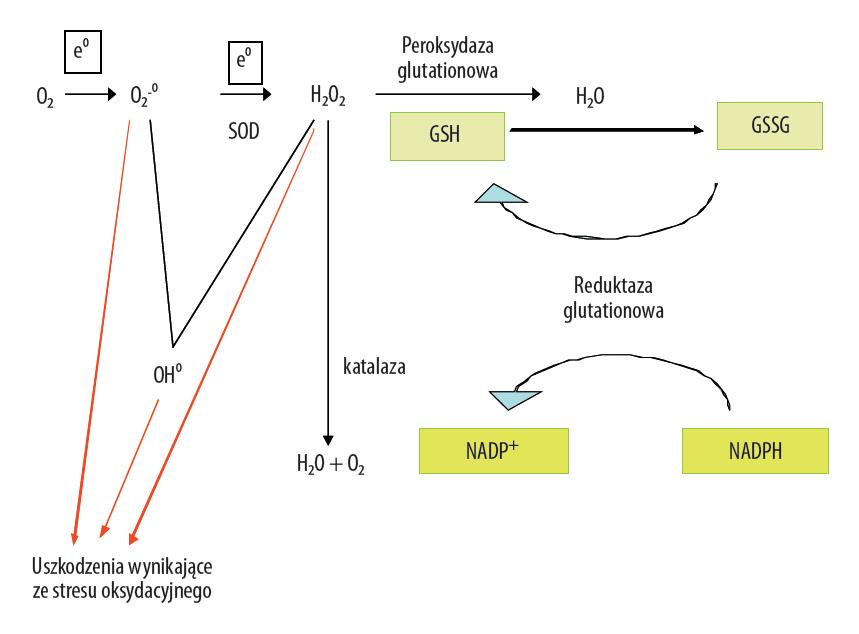
Ryc. 1. Wytwarzanie wolnych rodników tlenowych i aktywność endogennego układu antyoksydacyjnego
Należy zaznaczyć, że proces tworzenia potencjalnie szkodliwych wolnych rodników jest wszechobecnym zjawiskiem w organizmie żywym, zachodzącym w różnych typach komórek i tkanek w ciągu całego życia. Jednakże właśnie siatkówka stanowi szczególnie dogodne miejsce do powstawania RFT z kilku powodów. Wśród nich jednym z ważniejszych jest wysokie ciśnienie parcjalne tlenu, który jest substratem dla rodników tlenowych. Siatkówka, a zwłaszcza jej fotoreceptory – te niezwykle intensywnie pracujące i charakteryzujące się intensywnym metabolizmem komórki, wymagają dużej ilości tlenu i składników odżywczych. Wszystkie te elementy są dostarczane w niezbędnych ilościach z krwią drogą systemu choroidalnego, który tworzy naczyniówka wraz z rozbudowaną siecią mikronaczyń graniczących z błoną Brucha i dalej z warstwą komórek RPE. Ze względu na duży popyt tlenu organizm zwiększa jego podaż i stąd, w porównaniu z innymi tkankami, wysoki poziom substratu dla reaktywnych form tlenu sprzyja ich generowaniu w siatkówce [4,15,56,59]. Tworzeniu w siatkówce wolnych rodników sprzyjają i inne czynniki, takie jak: promieniowanie, obecność wielonienasyconych kwasów tłuszczowych (WNKT) podatnych na działanie RFT oraz obecność różnorodnych chromoforów – fotouwrażliwiaczy. Wśród nich wymienia się określone składniki lipofuscyny, np. A2E, czy związki karboksyalkilopirolowe, np. karboksyetylopirol (CEP), pochodne WNKT, a zwłaszcza najbardziej nienasyconego kwasu tłuszczowego – DHA, przybierające postać m.in. immunogennych adduktów/koniugatów typu CEP-białko [4,37,39,56].
2.1. Promieniowanie
Oko narażone jest na szeroki zakres promieniowania, od nadfioletowego (UV-C, 220–290 nm; UV-B, 290–320 nm; UV-A, 320–400 nm) do widzialnego (400–700 nm). Promieniowanie krótkofalowe jest wysokoenergetyczne, natomiast długofalowe, niosące kwanty niskoenergetyczne (>600 nm) wykazują zdolność do przenikania w głąb oka. Promieniowania krótko- i długofalowe są niebezpieczne i mogą powodować biologiczne uszkodzenia. Pierwszym „filtrem” docierającego do oka światła jest rogówka, która może absorbować promieniowanie poniżej 295 nm; dalej, soczewka absorbuje promieniowanie do 400 nm, a siatkówka w zakresie 400–760 nm (zakres światła widzialnego). Zatem zarówno fotoreceptory, jak i nabłonek barwnikowy siatkówki mogą ulegać uszkodzeniom wywołanym przez promieniowanie, ale rodzaj uszkodzeń jest uzależniony od intensywności, czasu trwania promieniowania oraz przerw w ekspozycji na światło, a także rodzaju dystrybucji promieniowania [3,42].
Oko, którego funkcja polega na odbieraniu i przetwarzaniu bodźców świetlnych, narażone jest na działania niepożądane promieniowania, dlatego ważne jest ograniczanie jego wpływu (np. okulary przeciwsłoneczne), bowiem obecność tlenu i fotonu jest wystarczającym warunkiem do powstawania rodników. Rozwijająca się wraz z wiekiem zaćma w pewnym stopniu zmniejsza narażenie siatkówki na promieniowanie, dlatego ryzyko rozwoju AMD po operacji usunięcia zaćmy na ogół zwiększa się w ciągu 5 lat [3,14].
2.2. Chromofory
Chromofory lub inaczej fotouwrażliwiacze są cząsteczkami, które absorbują światło i inicjują reakcje chemiczne. Siatkówka zawiera chromofory, takie jak chromoproteiny typu rodopsyny (i innych pigmentów wzrokowych), melanina, lipofuscyna i enzym łańcucha oddechowego mitochondriów – oksydazę cytochromu c [4].
Lipofuscyna (jako jeden z ważniejszych elementów stanowiących potencjalne podłoże dla rozwoju AMD) jest tworzona i odkładana progresywnie wraz z wiekiem w komórkach nabłonka barwnikowego siatkówki (RPE) [11,36,37,39]. Zmagazynowany, a jednocześnie wystawiony na promieniowanie światła widzialnego, A2E sprzyja generowaniu dużych ilości RFT. Badania Sparrow i wsp. [51,52] wykazały, że A2E przy jednoczesnej ekspozycji na światło niebieskie, uczestniczy w dalszym promowaniu tworzenia wolnych rodników tlenowych, a nawet prowadzi do apoptozy komórek ARPE-19 (linii komórkowej wywodzącej się z ludzkiego nabłonka barwnikowego siatkówki); natomiast przy nieobecności A2E w zawiesinie/hodowli komórek ARPE-19 nie dochodziło do ich śmierci, nawet przy ekspozycji na światło niebieskie.
Dodatkową przyczyną generowania dużej ilości rodników, a wywołanych obecnością lipofuscyny, jest inaktywacja przez nią enzymów układu antyoksydacyjnego, np. katalazy w cytoplazmie i lizosomach. W obecności lipofuscyny i jednoczesnej ekspozycji na światło zmniejsza się aktywność dysmutazy ponadtlenkowej (SOD), peroksydazy i reduktazy glutationowej w komórkach RPE [11,45].
2.3. Fagocytarna funkcja komórek nabłonka barwnikowego
Komórki nabłonka barwnikowego siatkówki (RPE), które swoimi wypustkami otulają zewnętrzne segmenty fotoreceptorów (POS), pełnią wiele funkcji niezbędnych do przeżycia fotoreceptorów [37,39], wśród nich fagocytują szczytowe fragmenty POS. Fotoreceptory zawierają pigmenty wzrokowe, np. barwnik „pręcikowy” – rodopsynę, odpowiedzialną za pochłanianie fotonów i detekcję obrazów. W procesie odbioru wrażeń świetlnych dochodzi do zużywania szczytowych fragmentów POS, które są złuszczane, a następnie pochłaniane przez komórki RPE, czemu może towarzyszyć tzw. „wybuch tlenowy”, albowiem w pochłanianym materiale pochodzącym z POS znajdują się zmodyfikowane, utlenione postaci wielu związków lipidowych i białek, które znacznie trudniej podlegają metabolizmowi i inaktywacji, dlatego odkładają się w lizosomach w postaci ziaren lipofuscyny. W wyniku fagocytozy i metabolizmu złuszczonych w procesie widzenia fragmentów POS są generowane rodniki (utlenione białka i lipidy) oraz kolejne substancje fotouwrażliwiające, a to przyczynia się do nasilenia i propagacji stresu oksydacyjnego [33,34,36,37,39].
2.4. Wielonienasycone kwasy tłuszczowe
Błona komórkowa fotoreceptorów (czopków i pręcików) zawiera wielonienasycone kwasy tłuszczowe (WNKT), które stanowią 50% podwójnej warstwy białkowo-lipidowej, a pozostałe 50% to białka. Natomiast kwas dokozaheksaenowy (docosahexaenoic acid – DHA) stanowi >60% wśród nienasyconych kwasów tłuszczowych obecnych w błonie komórkowej fotoreceptorów. W przeliczeniu na błonowe fosfolipidy, zawartość DHA może być relatywnie jeszcze większa [35,38]. DHA (C22:6-ω3), zawierający łańcuch węglowodorowy zbudowany z 22 atomów węgla (C22) z 6 podwójnymi wiązaniami C=C, należy do długołańcuchowych wielonienasyconych kwasów tłuszczowych szeregu omega 3 (WNKT-ω3). Przynależność do rodziny omega 3 oznacza, że w strukturze takich kwasów pierwsze podwójne wiązanie C=C występuje przy 3 atomie węgla licząc od końca cząsteczki, tj. od grupy metylowej zawierającej atom węgla C22 (czyli węgiel omega – ω). Przemiany w szeregu kwasów tłuszczowych omega 3 inicjuje kwas α-linolenowy (ALA; C18:3-ω3), a innymi przedstawicielami w tej rodzinie są takie związki, jak: kwas eikozapentaenowy (EPA; C20:5-ω3) i kwas dokozapentaenowy (DPA; C22:5-ω3) – wszystkie występują w dużych ilościach w błonach plazmatycznych fotoreceptorów [35,38]. Poza kwasami szeregu omega 3, drugą grupę związków lipidowych występujących powszechnie w błonach plazmatycznych wszystkich komórek w organizmie, w tym komórek siatkówki, stanowią kwasy szeregu omega 6 (ω6), do których zalicza się m.in. kwas arachidonowy (ARA; 20:4-ω6).
Większość naturalnie występujących nienasyconych kwasów tłuszczowych ma podwójne wiązania o konformacji cis, dzięki czemu łańcuchy węglowodorowe cząsteczek są zgięte w miejscu podwójnego wiązania o 120°. Nienasycone kwasy tłuszczowe zawierające kilka podwójnych wiązań o konformacji cis nadają błonie komórkowej określone możliwości (w przeciwieństwie do nasyconych kwasów tłuszczowych niemających wiązań podwójnych C=C), takie jak: płynność, elastyczność oraz predysponują białka-receptory do zmian konformacji, a tym samym odbierania i przekazywania informacji między neuronami. Z tego powodu, nienasycone kwasy tłuszczowe są obecne przede wszystkim w błonie komórkowej neuronów bogatych w różnego rodzaju receptory oraz fotoreceptorów zawierających barwnik wzrokowy – np. rodopsynę (receptora dla fotonów światła) [37,38,56].
Reasumując, przesyłanie informacji między neuronami oraz odbieranie bodźców świetlnych przez pigmenty wzrokowe (np. rodopsynę) wymaga zmiany konformacji receptorów, co jest uzależnione od składu błony komórkowej, a przede wszystkim od obecności nienasyconych kwasów tłuszczowych z wieloma podwójnymi wiązaniami o konfiguracji cis. Jednakże peroksydacja kwasów tłuszczowych (a jest tym większa, im więcej dany związek zawiera podwójnych wiązań C=C w łańcuchu węglowodorowym) prowadzi do kaskady reakcji wolnorodnikowych, których efektem jest powstawanie RFT2. Produkty utleniania WNKT zmieniają właściwości fizyczne błony komórkowej (zmniejszenie różnic potencjałów elektrycznych po obu stronach błony), czego następstwem jest utrata funkcjonalności oraz integralności strukturalnej błony komórkowej. Metabolity, powstające na skutek utleniania i defragmentacji tychże kwasów, takie jak np. kwas 4-hydroksy-7-oksyhept-5-enowy (HOHA), stanowią substrat dla 2-(ω-karboksyetylo)pirolu (CEP). Ten ostatni związek może się przyczyniać do generacji wolnych rodników, a także może przyłączać cząsteczki białek, tworząc wysoce reaktywne addukty CEP-białko, wcześniej wspomniane immunogenne struktury zdolne do aktywacji układu dopełniacza [37,39]. Ponadto produkty peroksydacji lipidów, takie jak HNE (4-hydroksy- 2-nonenal) oraz MDA (dialdehyd malonowy), mogą hamować aktywność proteazy lizosomalnej RPE, a w dalszej kolejności prowadzić do dysfunkcji lizosomalnej i nasilenia lipofuscynogenezy [27].
Prawie 30 lat temu pojawiły się doniesienia mówiące o powstawaniu z kwasu arachidonowego (20:4-ω6), w wyniku jego nieenzymatycznej peroksydacji z udziałem wolnych rodników, izoprostanów – związków przypominających prostaglandyny [30]. Dalsze badania wykazały, że w wyniku przemian peroksydacyjnych, izoprostany różnych typów mogą powstawać zarówno w warunkach in vitro jak i in vivo z WNKT szeregu omega 3, takich jak kwas eikozapentaenowy (EPA) i kwas dokozaheksaenowy (DHA) [29,49]. Izoprostany pochodzące z DHA w obrębie tkanek ośrodkowego układu nerwowego nazywane są neuroprostanami; co ciekawe, powstaje ich więcej niż innych izoprostanów [43]. W warunkach in vivo i zwiększonego ciśnienia parcjalnego tlenu z kwasu arachidonowego powstają dodatkowe związki o strukturze izofuranu, a z DHA strukturalnie podobne związki nazywane neurofuranami [48]. Ich zwiększone stężenie wykazano w korze mózgowej u zwierząt służących za model choroby Alzheimera [48,49]. Już pierwsze obserwacje dotyczące wolnorodnikowego tworzenia izoprostanów z kwasu arachidonowego sugerowały, iż związki te są mediatorami stresu oksydacyjnego. Sugestia taka znalazła potwierdzenie w wynikach dalszych badań dotyczących wolnorodnikowego tworzenia izofuranów i neurofuranów w płynach biologicznych, takich jak mocz, krew czy płyn mózgowo-rdzeniowy, może być miarodajnym wskaźnikiem (biomarkerem) intensywności stresu oksydacyjnego i peroksydacji lipidów w komórce/ tkance/organizmie, a w tkance nerwowej – zaawansowania procesu neurodegeneracji [48,49]. Jak do tej pory nie ma opublikowanych danych o tworzeniu izoprostanów, izofuranów i neurofuranów w siatkówce, wydaje się jednak bardzo prawdopodobne, że takie tworzenie zachodzi, bowiem przekonujące dowody przemawiają za istotną rolą stresu oksydacyjnego w obrębie anatomiczno-funkcjonalnego kompleksu RPE-fotoreceptory i w patogenezie AMD [37,39].
Podsumowując warunki istniejące w siatkówce można wnioskować, że utlenianie lipidów oraz białek przy jednoczesnym osłabieniu aktywności enzymów o charakterze antyoksydacyjnym na skutek obecności lipofuscyny ocznej szczególnie wrażliwej na wszechobecne światło widzialne, prowadzi do intensywnego wytwarzania reaktywnych form tlenu. Rozwijający się stres oksydacyjny napędza dalsze tworzenie wolnych rodników, a także aktywuje układ dopełniacza (zwłaszcza jego ścieżkę alternatywną), czego konsekwencją może być rozwinięcie się procesu przypominającego zapalenie, tj. parainflammation [35,37]. Istniejąca druzogeneza wraz z długotrwałą lipofuscynogenezą przy jednoczesnym braku buforów stresu oksydacyjnego (tj. lokalnych czynników obrony antyoksydacyjnej) może prowadzić do wystąpienia reakcji immunologicznej oraz uaktywnienia się procesu parainflammation, co znacznie przyspiesza progresję choroby i utratę widzenia centralnego (ryc. 2) [37,39].

Ryc. 2. Udział stresu oksydacyjnego w patogenezie AMD
Wrażliwość siatkówki ludzkiej, a przede wszystkim składników błony plazmatycznej/komórkowej fotoreceptorów, jest regionalna, zależna od wieku i może się zmieniać na skutek modyfikacji diety, zwłaszcza zawierającej związki o właściwościach przeciwutleniających, zdolnych do neutralizacji wolnych rodników tlenowych. W dalszej części artykułu zostaną przedstawione i poddane dyskusji związki systemu ochrony antyoksydacyjnej, stanowiące często elementy suplementów diety zalecanych w profilaktyce AMD, obecnych na rynku polskim.
3. Układ antyoksydacyjny
W organizmie znajduje się układ antyoksydacyjny dwojakiego rodzaju: układ enzymatyczny i nieenzymatyczny. Inny podział uwzględnia jego naturę endogenną i egzogenną. Układ endogenny–enzymatyczny składa się z 3 zasadniczych enzymów: dysmutazy ponadtlenkowej (SOD), katalazy i peroksydazy glutationowej, które są uzależnione jednak od elementów egzogennych, jakimi są jony metali: cynku, miedzi, manganu i selenu, bez których enzymatyczny układ antyoksydacyjny nie może funkcjonować [4,56]. Część nieenzymatyczną układu antyoksydacyjnego stanowią takie związki jak: karotenoidy, witaminy (witamina E i C) oraz glutation. Poniżej omówimy najważniejsze z punktu widzenia AMD czynniki egzogenne, które są przedmiotem badań i dyskusji dotyczącej ich roli i skuteczności jako środków prewencyjnych.
3.1. Karotenoidy: luteina, zeaksantyna i mezozeaksantyna
Karotenoidy należą do tetraterpenów (grupa terpeny, inaczej izoprenoidy), zbudowanych z 8 jednostek izoprenowych oraz stanowią obszerną grupę barwników występujących (a także pełniących określone funkcje) w królestwie roślin i zwierząt. Wśród karotenoidów wyróżnia się dwie grupy: karoteny i ksantofile; te ostatnie są tlenowymi pochodnymi, określanymi jako oksykarotenoidy. Najbardziej rozpowszechnionym karotenem jest α- i β-karoten (powszechnie znany ze względu na obecność np. w marchwi). Do ksantofilów należą: luteina i zeaksantyna, które obecnie przeżywają renesans zainteresowania ze względu na obecność i ochronne funkcje w siatkówce, a także rolę w zapobieganiu AMD.
Luteina jest żółtym barwnikiem o wzorze sumarycznym C40H56O2 i masie cząsteczkowej 568,88 Da; jej pełna nazwa chemiczna brzmi (3R, 3’R, 6’R)beta, epsilon-karoten- 3,3’-diol. Luteina może występować w 8 stereoizomerycznych formach, gdyż ma 3 asymetryczne atomy węgla w pozycjach 3, 3’ i 6’. Luteina w łańcuchu polienowym (łączącym dwa pierścienie) zawiera 9 sprzężonych podwójnych wiązań.
Wzór sumaryczny i masa cząsteczkowa zeaksantyny, a także mezozeaksantyny, jest taki sam jak luteiny. Zeaksantyna różni się od luteiny położeniem podwójnego wiązania w cyklicznym b pierścieniu a jej nazwa chemiczna to: (3R, 3’R)-beta, beta-karoten-3,3’-diol. Ze względu na obecność tylko dwóch stereocentrycznych atomów węgla istnieje w 3 izoformach. Jedną z nich jest mezozeaksantyna – prawdopodobnie powstająca z luteiny, a jej nazwa chemiczna to: (3R, 3’S)beta, beta-karoten-3,3’-diol [24,26,28,56,58]. Na ryc. 3 przedstawiono struktury chemiczne ksantofilów.

Ryc. 3. Struktury ksantofilów obecnych w siatkówce
Aktywność biochemiczna luteiny, zeaksantyny i mezozeaksantyny
George Wald w 1945 roku zanalizował barwnik plamki ocznej (nazywanej dawniej plamką żółtą), odkrywając, że za barwę żółtą odpowiedzialne są naturalne antyoksydanty, dihydroksykarotenoidy – ksantofile. Tlenowe pochodne karotenoidowe znajdują się w siatkówce, przy czym obie formy zeaksantyny (zeaksantyna i mezozeaksantyna) są dominujące w plamce ocznej, a luteina jest dominującym składnikiem peryferyjnej części siatkówki. Luteina, zeaksantyna oraz mezozeaksantyna stanowią barwnik plamki ocznej (macular pigment – MP) i obecne są w siatkówce w stosunku 2:1:1. Porównując poszczególne ksantofile obecne w siatkówce należy podkreślić, że luteina i zeaksantyna to składniki pochodzące z pożywienia (obecne i syntetyzowane w roślinach), natomiast mezozeaksantyna nie pochodzi z diety, chociaż są nieliczne doniesienia o jej obecności w naturze (u skorupiaków i rozgwiazdy), jednak te składniki nie mogą stanowić źródła dla człowieka [1,53,55]. Istnieje kilka elementów składających się na hipotezę, że mezozeaksantyna powstaje w siatkówce z luteiny. Bone i wsp. [8,9] wykazali, że w osoczu poziom luteiny jest wyższy niż zeaksantyny i stanowi 3:1, natomiast w siatkówce jest 2:1. Ponadto mezozeaksantyny nie stwierdza się w osoczu ani w wątrobie, ale występuje w plamce ocznej. Omówiony rozkład ksantofilów w poszczególnych kompartmentach pozwala wnioskować, że luteina ulega konwersji enzymatycznej lub fotochemicznej („migracja” podwójnego wiązania) do mezozeaksantyny. W oczach płodu, noworodków i dzieci do 2 r.ż. obserwuje się znacznie mniejszą ilość mezozeaksantyny w porównaniu z zeaksantyną, a dominującym karotenoidem jest luteina, co tłumaczy się niedorozwojem odpowiednich mechanizmów przekształcających luteinę w mezozeaksantynę. Również badania na małpach potwierdzają przemianę luteiny do mezozeaksantyny w siatkówce [9,23].
Jedną z hipotez dotyczącą etiologii AMD jest osłabienie układu antyoksydacyjnego w siatkówce, która ma wyjątkowo korzystne warunki do tworzenia wolnych rodników tlenowych, dlatego związki o charakterze antyoksydantów mogą odgrywać rolę w zapobieganiu tego schorzenia. W przypadku ksantofilów znajdujących się w siatkówce rozważa się dwa mechanizmy działania składające się na pełnioną funkcję ochronną, to jest absorpcja światła niebieskiego (funkcja filtra) oraz funkcja „zmiataczy” wolnych rodników. Te właściwości ksantofilów wynikają również z ich obecności i ułożenia w błonie komórkowej, a także ochrony m.in. wielonienasyconych kwasów tłuszczowych. Luteina może się układać zarówno pionowo, jak i poziomo (równolegle) w błonie komórkowej, natomiast zeaksantyna pionowo, tj. w poprzek błony [20].
Junghans i wsp. [24] wykazali zdolność karotenoidów do absorpcji światła niebieskiego w następującej kolejności: luteina > zeaksantyna > β-karoten > likopen. A2E i jego pochodne (składniki lipofuscyny ocznej) pochłaniają promieniowanie w zakresie 450 nm i tym samym generują RFT. Ksantofile obecne w siatkówce (fotoreceptorach i nabłonku barwnikowym siatkówki) pochłaniają promieniowanie w tym samym zakresie, dlatego stanowią filtr dla światła niebieskiego i zapobiegają fotooksydacji m.in. A2E. Właściwości antyoksydacyjne ksantofile zawdzięczają liczbie sprzężonych podwójnych wiązań między atomami węgla, z tego względu zeaksantyna wykazuje wiekszą aktywność antyoksydacyjną od luteiny (zeaksantyna ma 11 podwójnych wiązań sprzężonych: 9 w łańcuchu polienowym i po jednym w cyklicznych β pierścieniach, natomiast luteina – 10 podwójnych wiązań sprzężonych: 9 w łańcuchu polienowym i jedno w cyklicznym β pierścieniu – ryc. 3) [7,8,9,10,26,28,58].
Na podstawie funkcji i rozłożenia ksantofilów, niektórzy autorzy uważają [1,26,28,47,58], że luteina stanowi ochronę dla pręcików, które koncentrują się w peryferyjnej części siatkówki, a zeaksantyna oraz mezozeaksantyna chronią czopki, obecne z kolei w centralnej części siatkówki i stąd takie ich rozłożenie. Ponadto wymieniane ksantofile chronią fotoreceptory przed apoptozą indukowaną stresem oksydacyjnym [13].
3.2. β-Karoten
Suplementy diety zalecane w profilaktyce AMD zawierają niekiedy w swoim składzie witaminę A lub β-karoten. Jednak stosowanie obu tych składników w profilaktyce zwyrodnienia plamki związanego z wiekiem niesie ze sobą wiele kontrowersji i zastrzeżeń. Beta-karoten w przeciwieństwie do trzech omawianych wcześniej ksantofilów nie jest składnikiem barwnika plamki ocznej [21,55]. Ponadto stosowanie jego u palaczy sprzyja rozwojowi raka płuc, co dowiodły badania ATBC (alpha-tocopherol, beta-caroten cancer prevention study) [2]. Beta-karoten jest prekursorem witaminy A (retinolu), która odgrywa główną rolę w cyklu wzrokowym, a jednocześnie stanowi podstawowy element budujący toksyczny składnik lipofuscyny ocznej – A2E (o czym wspomniano wcześniej), dlatego nadmiar witaminy A sprzyja tworzeniu i odkładaniu w lipofuscynie ocznej bis-retinoidu – związku A2E [37,50], natomiast zmniejszenie jej podaży hamuje jego powstawanie. Z tych powodów witamina A oraz β-karoten nie powinny być rekomendowane w zapobieganiu czy leczeniu AMD.
3.3. Witaminy
Oprócz obecnie szczególnie zalecanych ksantofilów, w skład niektórych preparatów farmaceutycznych wchodzą typowe związki o charakterze przeciwutleniaczy, takie jak glutation oraz witamina E i C (tab. 1).
Tabela 1. Preparaty farmaceutyczne zawierające antyoksydanty bez witaminy A i jej prekursora

Witamina C (kwas askorbinowy) jest rozpuszczalnym w wodzie antyoksydantem zewnątrz- i wewnątrzkomórkowym. Może wchodzić w reakcje z RFT, takimi jak: anionorodnik ponadtlenkowy, rodnik hydroksylowy, rodniki nadtlenkowe oraz nadtlenek wodoru, tlen singletowy i unieszkodliwiać je. Witamina C pośrednio chroni również lipidy błony komórkowej przez reaktywację witaminy E [4,46,56,59]. Natomiast witamina E jako cząsteczka rozpuszczalna w tłuszczach, jest umiejscowiona głównie w błonie komórkowej, również fotoreceptorów, gdzie chroni kwasy tłuszczowe przed ich utlenianiem. Tokoferole i tokotrienole zapobiegają peroksydacji lipidów, a także wygaszają RFT, tj. tlen singletowy, anionorodnik ponadtlenkowy oraz rodnik hydroksylowy. W wyniku tych reakcji witamina E staje się rodnikiem, który jest regenerowany przez witaminę C. Na tej podstawie opisuje się synergizm między tymi dwoma związkami. W dalszym etapie w tej interakcji uczestniczy glutation oraz NADPH z reduktazą glutationową [4,46,56,59]. Synergistyczne działanie w zapobieganiu fotoutleniania A2E obserwowano po połączeniu ksantofilów z antyoksydantami rozpuszczalnymi w lipidach (witamina E) oraz rozpuszczalnymi w wodzie (witamina C) [25,62].
3.4. Glutation
Glutation jest tripeptydem (γ-glutamylo-cysteinylo-glicyna) rozpuszczalnym w wodzie. Jest on endogennym antyoksydantem, obecnym i syntetyzowanym w komórkach eukariotycznych, w których może występować w postaci zredukowanej (GSH) oraz utlenionej (GSSG). Synteza GSH odbywa się w cytosolu komórki w dwustopniowej reakcji przebiegającej z udziałem zależnych od ATP enzymów: syntetazy g-glutamylocysteinowej oraz syntetazy glutationowej. Synteza GSH jest regulowana poprzez ujemne sprzężenie zwrotne, co oznacza, że produkt końcowy, a więc glutation, hamuje aktywność syntetazy γ-glutamylocysteinowej. Ważnym elementem budowy glutationu jest wiązanie izopeptydowe między grupą karboksylową glutaminianu a grupą aminową cysteiny, które chroni cząsteczkę peptydu przed działaniem peptydaz [6,56,57,60].
Dostarczanie glutationu w postaci suplementów diety jest kontrowersyjne ze względu na jego katabolizm, bowiem zachodzi on w przestrzeni zewnątrzkomórkowej z udziałem enzymów: γ-glutamylotranspeptydazy (γ-GT) oraz dipeptydazy cysteinylo-glicynowej, które są związane z błoną komórkową. Obecność enzymów rozkładających tenże peptyd powoduje, że glutation nie może być pobrany z krwi, tylko komórka musi go syntetyzować, dlatego pochodzący z pożywienia stanowi źródło jedynie aminokwasów, zwłaszcza cysteiny [6,56,57,60]. Dostarczanie tych aminokwasów nie musi się przekładać na wzrost poziomu GSH w komórce, ponieważ syntetaza γ-glutamylocysteinowa reguluje jego syntezę w zależności od potrzeb komórki.
Glutation pełni w organizmie rolę antyoksydanta oraz kofaktora enzymu – peroksydazy glutationowej stanowiącej endogenny, enzymatyczny układ antyoksydacyjny. Jedną z ważniejszych jego reakcji jako przeciwutleniacza jest detoksykacja nadtlenku wodoru, która jest katalizowana przez peroksydazę glutationową, zwłaszcza w mitochondriach, gdzie brakuje katalazy (głównego enzymu rozkładającego H2O2). Istnieją doniesienia, że glutation oraz N-acetylocysteina (prekursor cysteiny, podstawowego aminokwasu glutationu) zapobiega apoptozie komórki, wywołanej działaniem nadtlenku wodoru. Obniżenie poziomu GSH i towarzyszące temu nagromadzenie się RFT, przyczynia się do dysfunkcji mitochondriów odgrywających decydującą rolę w kaskadzie apoptotycznej, m.in. uwolnienie do cytoplazmy komórki cytochromu c oraz czynnika inicjującego apoptozę (apoptosis-inducing factor – AIF). Podobna reakcja może zachodzić w komórkach nabłonka barwnikowego siatkówki, prowadząc tym samym do ich apoptozy, a w dalszym etapie do rozwoju AMD [22,31,32]. Jego wyższość jako endogennego antyoksydanta w porównaniu z innymi polega na możliwości własnej regeneracji z udziałem reduktazy glutationowej [61] oraz regeneracji witaminy C, o czym wcześniej już wspomniano.
4. Antyoksydanty i profilaktyka AMD
Obecnie przyjmuje się, że stres oksydacyjny stanowi jedno z ogniw w patomechanizmie AMD, dlatego też stosowanie związków o charakterze przeciwutleniającym jest elementem strategii profilaktyczno-terapeutycznej tego schorzenia [4,15,33,37,56,59]. Tym bardziej że nie ma jeszcze skutecznej terapii AMD, zwłaszcza jej postaci suchej (degeneracyjnej, zanikowej).
Autorzy spróbowali wyjaśnić, które związki szczególnie powinny być brane pod uwagę podczas projektowania preparatów farmaceutycznych zalecanych w profilaktyce lub jako składniki pomocnicze w leczeniu AMD. Jednakże zgodnie z zasadą: „co za dużo to niezdrowo” lub „nadmiar dobrego może szkodzić”, również w tym wypadku nadmiar antyoksydantów może być przyczyną stresu oksydacyjnego. Na rycinie 4 przedstawiono kaskadę wzajemnych relacji między poszczególnymi antyoksydantami, które będąc w równowadze współdziałają i znoszą powstające rodniki. Nadmiar jednego z elementów prowadzi do zaburzenia homeostazy i powstawania rodników, bowiem każdy związek o charakterze antyoksydacyjnym może się stać prooksydacyjnym [16,44]. Z tych powodów, w profilaktyce AMD ważny jest dobór składników preparatów czy też rodzaj diety (bogatej w antyoksydanty), które mają hamować występowanie stresu oksydacyjnego w siatkówce. Oprócz tego istotne są również odpowiednie dawki, które są trudne do określenia, a które powinny eliminować rodniki, a nie stanowić podłoża do ich powstawania.

Ryc. 4. Funkcjonalne interakcje między antyoksydantami
Na podstawie najnowszych badań wiadomo, że nadmiar witaminy A (a także jej prekursora β-karotenu) prowadzi do zwiększenia ilości lipofuscyny ocznej, dlatego nie powinna ona występować w preparatach rekomendowanych w AMD, tym bardziej że jest ona powszechnie dostępna w pożywieniu i w krajach cywilizowanych człowiek nie cierpi na jej niedobór. W tabeli 1 przedstawiono warte rekomendacji preparaty zawierające omawiane w artykule składniki, a niezawierające witaminy A ani β-karotenu (szerokie omówienie preparatów farmaceutycznych zawierających antyoksydanty, a także wielonienasycone kwasy tłuszczowe omega 3, zainteresowany czytelnik znajdzie w innych pracach autorów [38,56,57,58]). Ostatnie badania laboratoryjne Bhosale i wsp. [5] wykazały antagonistyczną funkcję luteiny i zeaksantyny wobec tworzenia A2E. Jednakże badania wieloośrodkowe sprawdzające skuteczność prewencyjno-terapeutyczną antyoksydantów w AMD nie zawsze dają dobre wyniki, a nawet niekiedy nie dowodzą ich roli, co wskazuje na wieloczynnikowość i złożoność etiopatogenezy AMD. Innym problemem wciąż jeszcze nierozwiązanym może być zdolność dystrybucji ksantofilów do miejsca działania.
W ofercie handlowej znajduje się kilkadziesiąt preparatów suplementacyjnych zawierających różne kompozycje antyoksydantów (występujące w postaci konkretnych związków bądź ekstraktów roślinnych), bez lub w towarzystwie witamin, mikroelementów, a także kwasów tłuszczowych omega 3 [56]. W nazwie bądź na opakowaniu/etykiecie wielu takich preparatów można odnaleźć sugestię rekomendacyjną odnośnie narządu wzroku – jednak czy wszystkie są ekwiwalentne w rozumieniu ich potencjału terapeutycznego? W opinii autorów niniejszego opracowania nie można stawiać znaku równości pomiędzy wszystkimi omawianymi preparatami. Czy istnieją więc obiektywne kryteria oceny ich wartości i przydatności w prewencji, a nawet terapii AMD? Badania doświadczalne i kliniczne wartości terapeutycznej poszczególnych składników i ich kombinacji są w toku – musimy zatem poczekać na udokumentowane wnioski końcowe. W tabeli 1 zamieszczono 16 preparatów wyróżniających się spośród wielu innych tym, że zawierają jeden, dwa lub trzy barwniki plamkowe: we wszystkich występuje luteina w ilości 3–20 mg/tabletkę bądź kapsułkę, zeaksantyna (0,2–2 mg) jest obecna w 11 preparatach (zaznaczone kolorem żółtym), natomiast mezozeaksantyna – tylko w jednym preparacie (10 mg). Choć nie ma definitywnych przesłanek wartościujących poszczególne preparaty, podejściem racjonalnym – i reprezentowanym przez obecnych autorów – wydaje się wybór takiego preparatu, który zawiera przynajmniej dwa pigmenty plamkowe. Pozostaje jeszcze czasokres i częstotliwość przyjmowania wybranego preparatu. Niepełna znajomość złożonej etiopatogenezy AMD uniemożliwia podjęcie leczenia przyczynowego, a charakter postępujący schorzenia podpowiada, że wszelkie zabiegi prewencyjno-terapeutyczne powinny trwać tak długo, jak trwa choroba, a więc do końca życia. Natomiast odnośnie częstotliwości stosowania preparatów suplementacyjnych, to regularność suplementowania (codziennie lub co drugi dzień, w zależności od preparatu i diety) wydaje się ważniejsza niż liczba tabletek/kapsułek zażywanych w ciągu dnia.
———————————————————————————————————————-
1 Szczegółowe omówienie procesu określanego terminem parainflammation, również w kontekście AMD, zainteresowany czytelnik znajdzie w obszernym niedawnym opracowaniu Nowaka [35].
2 Na peroksydację lipidów składają się następujące procesy: inicjacja, propagacja i terminacja. Proces inicjacji polega na reakcji OH• z WNKT wytwarzając rodnik lipidowy, który w reakcji z tlenem daje LOO· (rodnik nadtlenku lipidu), mający zdolność odrywania wodoru od innych cząsteczek i generowania kolejnych rodników L·. Takie rodniki w obecności żelaza Fe2+ ulegają przekształceniu do rodników alkoksylowych LO·, a następnie do peroksydowych, aby dalej ulec rozpadowi do reaktywnych aldehydów: 4-hydroksynonenalu, 4-hydroksyheksenalu, dialdehydu malonowego i akroleiny określanych jako wtórne, toksyczne mesendżery. Kwasy z szeregu omega 6, takie jak kwas linolowy czy arachidonowy, ulegają przekształceniu głównie do HNE (4-hydroksy-2-nonenalu), a omega 3 (EPA, DHA) do HHE (4-hydroksy- 2-heksenalu) [12,17].
PIŚMIENNICTWO
[1] Ahmed S.S., Lott M.N., Marcus D.M.: The macular xanthophylls. Surv. Ophthalmol., 2005; 50: 183-193
[PubMed]
[2] Albanes D., Heinonen O.P., Taylor P.R., Virtamo J., Edwards B.K., Rautalahti M., Hartman A.M., Palmgren J., Freedman L.S., Haapakoski J., Barrett M.J., Pietinen P., Malila N., Tala E., Liippo K., Salomaa E.R., Tangrea J.A., Teppo L., Askin F.B., Taskinen E., Erozan Y., Greenwald P., Huttunen J.K.: α-tocopherol and β-carotene supplements and lung cancer incidence in the alpha-tocopherol, beta-carotene cancer prevention study: effects of base-line characteristics and study compliance. J. Natl. Cancer Inst., 1996; 88: 1560-1570
[PubMed] [Full Text PDF]
[3] Algvere P.V., Marshall J., Seregard S.: Age-related maculopathy and the impact of blue light hazard. Acta Ophthalmol. Scand., 2006, 84; 4-15
[PubMed] [Full Text HTML] [Full Text PDF]
[4] Beatty S., Koh H., Phil M., Henson D., Boulton M.: The role of oxidative stress in the pathogenesis of age-related macular degeneration. Surv. Ophthalmol., 2000; 45: 115-134
[PubMed]
[5] Bhosale P., Serban B., Bernstein P.S.: Retinal carotenoids can attenuate formation of A2E in the retinal pigment epithelium. Arch. Biochem. Biophys., 2009; 483: 175-181
[PubMed] [Full Text PDF]
[6] Bilska A., Kryczyk A., Włodek L.: Różne oblicza biologicznej roli glutationu. Postepy Hig. Med. Dosw.: 2007; 61: 438-453
[PubMed] [Full Text PDF]
[7] Bone R.A., Landrum J.T., Cao Y., Howard A.N., Alvarez-Calderon F.: Macular pigment response to a supplement containing meso-zeaxanthin, lutein and zeaxanthin. Nutr. Metab., 2007; 4: 12
[PubMed] [Full Text HTML] [Full Text PDF]
[8] Bone R.A., Landrum J.T., Fernandez L., Tarsis S.L.: Analysis of the macular pigment by HPLC: retinal distribution and age study. Invest. Ophthalmol. Vis. Sci., 1988; 29: 843-849
[PubMed] [Full Text PDF]
[9] Bone R.A., Landrum J.T., Hime G.W., Cains A., Zamor J.: Stereochemistry of the human macular carotenoids. Invest. Ophthalmol. Vis. Sci., 1993; 34: 2033-2040
[PubMed] [Full Text PDF]
[10] Bone R.A., Landrum J.T., Tarsis S.L.: Preliminary identification of the human macular pigment. Vision Res., 1985; 25: 1531-1535
[PubMed]
[11] Boulton M., Rózanowska M., Rózanowski B., Wess T.: The photoreactivity of ocular lipofuscin. Photochem. Photobiol. Sci., 2004; 3: 759-764
[PubMed]
[12] Catala A.: Lipid peroxidation of membrane phospholipids generates hydroxy-alkenals and oxidized phospholipids active in physiological and/or pathological conditions. Chem. Phys. Lipids, 2009; 157: 1-11
[PubMed]
[13] Chucair A.J., Rotstein N.P., SanGiovanni J.P., During A., Chew E.Y., Politi L.E.: Lutein and zeaxanthin protect photoreceptors from apoptosis induced by oxidative stress: relation with docosahexaenoic acid. Invest. Ophthalmol. Vis. Sci., 2007; 48: 5168-5177
[PubMed] [Full Text HTML] [Full Text PDF]
[14] Dong L.M., Stark W.J., Jefferys J.L., Al-Hazzaa S., Bressler S.B., Solomon S.D., Bressler N.M.: Progression of age-related macular degeneration after cataract surgery. Arch. Ophthalmol., 2009; 127: 1412-1419
[PubMed]
[15] Drobek-Słowik M., Karczewicz D., Safranow K.: Potencjalny udział stresu oksydacyjnego w patogenezie zwyrodnienia plamki związanego z wiekiem (AMD). Postepy Hig. Med. Dosw., 2007; 61: 28-37
[PubMed] [Full Text PDF]
[16] El-Agamey A., Lowe G.M., McGarvey D.J., Mortensen A., Phillip D.M., Truscott T.G., Young A.J.: Carotenoid radical chemistry and antioxidant/pro-oxidant properties. Arch. Biochem. Biophys., 2004; 430: 37-48
[PubMed]
[17] Esterbauer H.: Cytotoxicity and genotoxicity of lipid-oxidation products. Am. J. Clin. Nutr., 1993; 57 (Suppl. 5): 779S-785S
[PubMed] [Full Text PDF]
[18] Fessel J.P., Hulette C., Powell S., Roberts L.J.2nd, Zhang J.: Isofurans, but not F2-isoprostanes, are increased in the substantia nigra of patients with Parkinson’s disease and with dementia with Lewy body disease. J. Neurochem., 2003; 85: 645-650
[PubMed] [Full Text HTML] [Full Text PDF]
[19] Gilca M., Stoian I., Atanasiu V., Virgolici B.: The oxidative hypothesis of senescence. J. Postgrad. Med., 2007; 53: 207-213
[PubMed]
[20] Gruszecki W.I., Strzałka K.: Carotenoids as modulators of lipid membrane physical properties. Biochim. Biophys. Acta, 2005; 1740: 108-115
[PubMed]
[21] Handelman G.J., Dratz E.A., Reay C.C., van Kuijk J.G.: Carotenoids in the human macula and whole retina. Invest. Ophthalmol. Vis. Sci., 1988, 29; 850-855
[PubMed] [Full Text PDF]
[22] Jin G.F., Hurst J.S., Godley B.F.: Hydrogen peroxide stimulates apoptosis in cultured human retinal pigment epithelial cells. Curr. Eye Res., 2001, 22; 165-73
[PubMed]
[23] Johnson E.J., Neuringer M., Russell R.M., Schalch W., Snodderly D.M.: Nutritional manipulation of primate retinas, III: Effects of lutein or zeaxanthin supplementation on adipose tissue and retina of xanthophyll-free monkeys. Invest. Ophthalmol. Vis. Sci., 2005; 46: 692-702
[PubMed] [Full Text HTML] [Full Text PDF]
[24] Junghans A., Sies H., Stahl W.: Macular pigments lutein and zeaxanthin as blue light filters studied in liposomes. Arch. Biochem. Biophys., 2001; 391: 160-164
[PubMed]
[25] Kim S.R., Nakanishi K., Itagaki Y., Sparrow J.R.: Photooxidation of A2-PE, a photoreceptor outer segment fluorophore, and protection by lutein and zeaxanthin. Exp. Eye Res., 2006; 82; 828-839
[PubMed]
[26] Krinsky N.I., Landrum J.T., Bone R.A.: Biologic mechanisms of the protective role of lutein and zeaxanthin in the eye. Annu. Rev. Nutr., 2003; 23: 171-201
[PubMed]
[27] Krohne T.U., Kaemmerer E., Holz P.G., Kopitz J.: Lipid peroxidation products reduce lysosomal protease activities in human retinal pigment epithelial cells via two different mechanisms of action. Exp. Eye Res., 2010; 90: 261-266
[PubMed]
[28] Landrum J.T., Bone R.A.: Lutein, zeaxanthin, and the macular pigment. Arch. Biochem. Biophys., 2001; 385: 28-40
[PubMed]
[29] Lawson J.A., Kim S., Powell W.S., FitzGerald G.A., Rokach J.: Oxidized derivatives of ω-3 fatty acids: identification of IPF3α-VI in human urine. J. Lipid Res., 2006; 47: 2515-2524
[PubMed] [Full Text HTML] [Full Text PDF]
[30] Lawson J.A., Rokach J., FitzGerald G.A.: Isoprostanes: formation, analysis and use as indices of lipid peroxidation in vivo. J. Biol. Chem., 1999; 274: 24441-24444
[PubMed] [Full Text HTML] [Full Text PDF]
[31] Liang F.Q., Godley B.F.: Oxidative stress-induced mitochondrial DNA damage in human retinal pigment epithelial cells: a possible mechanism for RPE aging and age-related macular degeneration. Exp. Eye Res., 2003; 76: 397-403
[PubMed]
[32] Medina S., Martinez M., Hernanz A.: Antioxidants inhibit the human cortical neuron apoptosis induced by hydrogen peroxide, tumor necrosis factor alpha, dopamine and beta-amyloid peptide l-42. Free Radic. Res., 2002; 36: 1179-1184
[PubMed]
[33] Nowak J.Z.: Age-related macular degeneration (AMD): pathogenesis and therapy. Pharmacol. Rep., 2006, 58: 353-363
[PubMed] [Full Text PDF]
[34] Nowak J.Z.: Druzy, złogi podstawne, proces zapalny i zwyrodnienie plamki związane z wiekiem (AMD). Magazyn Okulistyczny, 2005; 3: 174-186
[35] Nowak J.Z.: Przeciwzapalne “prowygaszeniowe” pochodne wielonienasyconych kwasów tłuszczowych omega-3 i omega-6. Postepy Hig. Med. Dosw., 2010; 64: 115-132
[PubMed]
[36] Nowak J.Z.: Rola lipofuscyny w etiopatogenezie zwyrodnienia plamki związanego z wiekiem (AMD). Magazyn Okulistyczny, 2005; 2: 103-114
[37] Nowak J.Z.: W poszukiwaniu biomarkerów dla zwyrodnienia plamki związanego z wiekiem (AMD). Mag. Lek. Okul., 2009; 3: 132-143
[38] Nowak J.Z.: Wielonienasycone kwasy tłuszczowe omega-3 w siatkówce i praktyce medycznej – blaski i cienie. Mag. Lek. Okul., 2009; 3: 208-220
[39] Nowak J.Z.: Zwyrodnienie plamki związane z wiekiem a układ odpornościowy: ile immunologii jest w AMD? Mag. Lek. Okul., 2009; 3: 102-114
[40] Nowak J.Z., Wiktorowska-Owczarek A.: Neowaskularyzacja w tkankach oka: mechanizmy i rola czynników pro- i antyangiogennych. Klin. Oczna, 2004; 106: 90-97
[PubMed]
[41] Potargowicz E., Szerszenowicz E., Staniszewska M., Nowak D.: Mitochondria jako źródło reaktywnych form tlenu. Postepy Hig. Med. Dosw., 2005; 59: 259-266
[PubMed] [Full Text PDF]
[42] Roberts J.E.: Ocular phototoxicity. J. Photochem. Photobiol. B, 2001, 64; 136-143
[PubMed]
[43] Roberts L.J.2nd, Montine T.J., Markesbery W.R., Tapper A.R., Hardy P., Chemtob S., Dettbarn W.D., Morrow J.D.: Formation of isoprostane-like compounds (neuroprostanes) in vivo from docosahexaenoic acid. J. Biol. Chem., 1998; 273: 13605-13612
[PubMed] [Full Text HTML] [Full Text PDF]
[44] Robman L., Vu H., Hodge A., Tikellis G., Dimitrov P., McCarty C., Guymer R.: Dietary lutein, zeaxanthin, and fats and the progression of age-related macular degeneration. Can. J. Ophthalmol., 2007; 42: 720-726
[PubMed] [Full Text PDF]
[45] Shamsi F.A., Boulton M.: Inhibition of RPE lysosomal and antioxidant acivity by the age pigment lipofuscin. Invest. Ophthalmol. Vis. Sci., 2001; 42: 3041-3046
[PubMed] [Full Text HTML] [Full Text PDF]
[46] Sies H., Stahl W., Sundquist A.R.: Antioxidant functions of vitamins. Vitamin E and C, β-carotene, and other carotenoids. Ann. NY Acad. Sci., 1992; 30: 669
[PubMed]
[47] Sommerburg O.G., Siems W.G., Hurst J.S., Lewis J.W., Kliger D.S., van Kuijk F.J.: Lutein and zeaxanthin are associated with photoreceptors in the human retina. Curr. Eye Res., 1999; 19: 491-495
[PubMed]
[48] Song W.L., Lawson J.A., Reilly D., Rokach J., Chang C.T., Giasson B., FitzGerald G.A.: Neurofurans, novel indices of oxidant stress derived from docosahexaenoic acid. J. Biol. Chem., 2008; 283: 6-16
[PubMed] [Full Text HTML] [Full Text PDF]
[49] Song W.L., Paschos G., Fries S., Reilly M.P., Yu Y., Rokach J., Chang C.T., Patel P., Lawson J.A., Fitzgerald G.A.: Novel eicosapentaenoic acid-derived F3-isoprostanes as biomarkers of lipid peroxidation. J. Biol. Chem., 2009; 284: 23636-23643
[PubMed]
[50] Sparrow J.R.: Therapy for macular degeneration: insight from acne. Proc. Natl. Acad. Sci. USA, 2003; 100: 4353-4354
[PubMed] [Full Text HTML] [Full Text PDF]
[51] Sparrow J.R., Cai B.: Blue light-induced apoptosis of A2E-containing RPE: involvement of caspase-3 and protection by Bcl-2. Invest. Ophthalmol. Vis. Sci., 2001; 42: 1356-1362
[PubMed] [Full Text HTML] [Full Text PDF]
[52] Sparrow J.R., Nakanishi K., Parish C.A.: The lipofuscin fluorophore A2E mediates blue light-induced damage to retinal pigmented epithelial cells. Invest. Ophthalmol. Vis. Sci., 2000; 41: 1981-1989
[PubMed]
[53] Thurnham D.I.: Macular zeaxanthins and lutein – a review of dietary sources and bioavailability and some relationships with macular pigment optical density and age-related macular disease. Nutr. Res. Rev., 2007; 20: 163-179
[PubMed]
[54] Thurnham D.I., Tremel A, Howard A.N.: A supplementation study in human subjects with a combination of meso-zeaxanthin, (3R,3’R)-zeaxanthin and (3R,3’R,6’R)-lutein. Br. J. Nutr., 2008; 100: 1307-1314
[PubMed]
[55] van Leeuwen R., Boekhoorn S., Vingerling J.R., Witteman J.C., Klaver C.C., Hofman A., de Jong P.T.: Dietary intake of antioxidants and risk of age-related macular degeneration. JAMA, 2005; 294: 3101-3107
[PubMed] [Full Text HTML] [Full Text PDF]
[56] Wiktorowska-Owczarek A., Nowak J.Z.: AMD a stres oksydacyjny. 1.Analiza czynników wpływających na powstawanie wolnych rodników tlenowych w siatkówce. 2.Analiza składników preparatów farmaceutycznych stosowanych w profilaktyce AMD. Mag. Lek. Okul., 2008; 3: 200-212
[57] Wiktorowska-Owczarek A., Nowak J.Z.: Glutation – endogenny tripeptyd o właściwościach przeciwutleniających: implikacje okulistyczne. Magazyn Okulistyczny, 2006; 3: 152-159
[58] Wiktorowska-Owczarek A., Nowak J.Z.: Luteina i zeaksantyna – dwa karotenoidy pełniące ochronną funkcję w zwyrodnieniu plamki związanym z wiekiem. Okulistyka, 2004; 4: 29-34
[59] Wiktorowska-Owczarek A., Nowak J.Z.: Oxidative damage in age-related macular degeneration (AMD) and antioxidant protection as a therapeutic strategy. Polish J. Environ. Stud., 2006; 15: 69-72
[60] Winiarska K.: Glutation: niezwykłe funkcje pospolitego tripeptydu. Postepy Biochem., 2000; 46: 318-326
[PubMed]
[61] Winkler B.S., Orselli S.M., Rex T.S.: The redox couple between glutathione and ascorbic acid: a chemical and physiological perspective. Free Radic. Biol. Med., 1994; 17: 333-349
[PubMed]
[62] Wrona M., Różanowska M., Sarna T.: Zeaxanthin in combination with ascorbic acid or a-tocopherol protects ARPE-19 cells against photosensitized peroxidation of lipids. Free Radic. Biol. Med., 2004; 36: 1094-1101
[PubMed]
Autorzy deklarują brak potencjalnych konfliktów interesów.