
Catalase: structure, properties, functions
Dorota Ścibior 1 , Hanna Czeczot 1
Streszczenie
Katalaza (EC 1.11.1.6), obecna głównie w peroksysomach komórek ssaków, jest enzymem zbudowanym z czterech identycznych podjednostek o masie cząsteczkowej około 60 kDa. Każda z nich zawiera w centrum aktywnym grupę hemową oraz cząsteczkę NADPH. Wykazuje dwie aktywności: katalazową i peroksydazową. Przy dużym stężeniu nadtlenku wodoru główną jej funkcją jest udział w jego rozkładzie do wody i tlenu (aktywność katalazowa). Natomiast przy małym stężeniu H2O2 dominuje aktywność peroksydazowa katalazy, a substratami są związki o charakterze donorów wodoru np. etanol, metanol, fenol i inne. Praca przedstawia aktualny stan wiedzy na temat budowy, właściwości i funkcji katalazy w organizmach żywych.
Słowa kluczowe:katalaza • budowa • właściwości • funkcje
Summary
Catalase (EC 1.11.1.6) is an enzyme which is present mainly in the peroxisomes of mammalian cells. It is a tetrameric enzyme consisting of four identical, tetrahedrally arranged subunits of 60 kDa, each containing in its active center a heme group and NADPH. Catalase has two enzymatic activities depending on the concentration of H2O2. If the concentration of H2O2 is high, catalase acts catalytically, i.e. removes H2O2 by forming H2O and O2 (catalatic reaction). However, at a low concentration of H2O2 and in the presence of a suitable hydrogen donor, e.g. ethanol, methanol, phenol, and others, catalase acts peroxidically, removing H2O2, but oxidizing its substrate (peroxidatic reaction). The review article presents current knowledge about the structure, properties, and functions of catalase in living organisms.
Key words:catalase • structure • properties • functions
Wykaz skrótów:
Arg – arginina; Asn – asparagina; Asp – asparaginian; CAT – katalaza; GSH – glutation; GSH-Px – peroksydaza glutationowa; G-6-PD – dehydrogenaza glukozo-6-fosforanowa; Hb – hemoglobina; His – histydyna; H2O2 – nadtlenek wodoru; MetHb – methemoglobina; NAD+ – utleniony nukleotyd nikotynoamidoadenionowy; NADH – zredukowany nukleotyd nikotynoamido-adenionowy; NADP+ – utleniony fosforan dinukleotydu nikotynoamido-adeninowego; NADPH – zredukowany fosforan dinukleotydu nikotynoamidoadeninowego; O2*– – anionorodnik ponadtlenkowy; OH* – rodnik hydroksylowy; 6-PGD – dehydrogenaza 6-fosfoglukonianowa; Phe – fenyloalanina; Pro – prolina; RFT – reaktywne formy tlenu; SH2 – związki będące donorami wodoru; SOD – dysmutaza ponadtlenkowa; Tyr – tyrozyna; Val – walina.
WSTĘP
Prawidłowemu metabolizmowi w komórkach organizmu towarzyszy powstawanie reaktywnych form tlenu (RFT) i ich pochodnych. W wyniku niepełnej redukcji O2 powstaje anionorodnik ponadtlenkowy O2*– (superoxide radical), który ulega spontanicznej i/lub enzymatycznej dysmutacji do nadtlenku wodoru (H2O2, hydrogen peroxide). W reakcji H2O2 z jonami metali (żelazo, miedź) powstaje najbardziej reaktywny rodnik hydroksylowy (OH*, hydroxyl radical). Wolne rodniki tlenowe i ich reaktywne pochodne łatwo reagują ze składnikami komórkowymi, takimi jak lipidy, białka i DNA. Skutkiem tych oddziaływań są uszkodzenia błon komórkowych, niewłaściwa aktywność bądź inaktywacja enzymów oraz mutacje genetyczne. Nadmiar wolnych rodników i ich pochodnych w komórkach i tkankach organizmu powoduje wiele zmian patologicznych [9,49,68,104].
W obronie przed uszkodzeniami oksydacyjnymi komórki wytworzyły wiele systemów ochronnych, które stanowią barierę antyoksydacyjną organizmu. W jej skład wchodzą enzymy, białka sekwestrujące metale, związki drobnocząsteczkowe (np. glutation, kwas moczowy, cysteina i inne), witaminy C, E i A. Enzymatyczną barierę antyoksydacyjną stanowią wyspecjalizowane enzymy o aktywności przeciwutleniającej. Głównym enzymem o działaniu antyoksydacyjnym jest dysmutaza ponadtlenkowa (SOD). W reakcji dysmutacji eliminuje ona ze środowiska komórki anionorodniki ponadtlenkowe, z których powstaje nadtlenek wodoru. Jest on rozkładany do wody i tlenu z udziałem katalazy lub peroksydazy glutationowej (ryc.1) [12,13,47,67,100,101].
Ryc. 1. Rozkład H2O2 z udziałem katalazy i peroksydazy glutationowej; CAT – katalaza; G-6-PD – dehydrogenaza glukozo-6-fosforanowa; GSH – zredukowany glutation; GSSG – utleniony glutation; GSH-Px – peroksydaza glutationowa; GSH-R – reduktaza glutationowa; H2O2 – nadtlenek wodoru; NADP+ – utleniony fosforan dinukleotydu nikotynoamido-adeninowego; NADPH – zredukowany fosforan dinukleotydu nikotynoamido- -adeninowego; 6-PGD – dehydrogenaza 6-fosfoglukonianowa
Pierwszą reakcję katalizuje katalaza (CAT):

Natomiast druga wymaga obecności glutationu (GSH) i jest katalizowana przez peroksydazę glutationową (GSH-Px):

Peroksydaza glutationowa odpowiada za katabolizm większości nadtlenku wodoru powstającego w komórkach [20,31,91]. Ma ona większe niż katalaza powinowactwo do H2O2, co wskazuje na szczególną jej rolę w detoksykacji tego związku, gdy jego stężenie w komórce jest małe. Przy dużym stężeniu H2O2 rolę tę przejmuje katalaza [12,14,95,108].
Działanie obu enzymów zapewnia usuwanie nadmiaru H2O2, dzięki czemu nie dochodzi do powstawania innych wolnych rodników i reaktywnych pochodnych tlenu [7,67,93].
H2O2 – SZCZEGÓLNY SUBSTRAT DLA CAT
H2O2 jest wytwarzany w komórkach w wielu reakcjach enzymatycznych, obejmujących m.in. reakcję dysmutacji, w której O2*– jest przekształcany do H2O2 i O2. Reakcja ta jest katalizowana przez dysmutazę ponadtlenkową. Nadtlenek wodoru może również powstawać w reakcjach nieenzymatycznych np. w procesie autoutlenienia związków tiolowych czy askorbinianu [6,7,27,99].
H2O2 jest naturalnym produktem metabolizmu komórkowego, a jednak ze względu na swoje właściwości utleniające, jest związkiem toksycznym dla komórek organizmu, ponieważ może uszkadzać białka, lipidy, cukry oraz DNA. Działanie to jest ułatwione ponieważ H2O2 łatwo dyfunduje przez błony biologiczne i może się pojawiać w innych kompartmentach/przedziałach komórki niż te, w których powstaje [7,16,48,57,99]. Nadtlenek wodoru w pH 7,0 (obojętnym) łatwo utlenia grupy tiolowe, imidiazolowe, fenolowe, tioestrowe oraz indolowe [48,89].
H2O2 to związek stosunkowo stabilny, który podlega reakcji dysproporcjonowania. Jest to możliwe dzięki występowaniu w jego strukturze bardzo słabego wiązania O–O, dla którego energia dysocjacji wynosi tylko 51 kcal/mol [6,48].

Biologicznie ważnymi reakcjami in vivo, w których uczestniczy nadtlenek wodoru są utlenianie grup tiolowych oraz utlenianie jonów metali przejściowych (Fe+2 do Fe+3; Cu+1 do Cu+2) [49,57,89].

Zdolność H2O2 do utleniania reaktywnych grup tiolowych powoduje uszkodzenia białek komórkowych, powstawanie mostków disiarczkowych, które zaburzają konformacje białek i zmieniają ich funkcje biologiczne. Z kolei utlenianie jonów metali przejściowych z udziałem H2O2 prowadzi do powstawania reaktywnego rodnika hydroksylowego OH* (reakcja Fentona i reakcja Habera-Weissa) [16,68,89].
Nadtlenek wodoru powoduje również utlenianie nienasyconych kwasów tłuszczowych w procesie peroksydacji lipidów, co powoduje zaburzenia struktury błony komórkowej i zmiany w jej płynności [6,7,49].
Wyżej wymienione właściwości H2O2 decydują o tym, że jest on uważany za związek toksyczny. Utrzymanie jego małych stężeń w komórkach (10–9–10–7 M) jest możliwe dzięki aktywności katalazy i peroksydazy glutationowej. W fizjologicznych warunkach oba te enzymy kontrolują stężenie H2O2 w komórkach, tak żeby nie dochodziło do jego nagromadzenia w nadmiarze, co mogłoby prowadzić do toksycznych oddziaływań na białka, lipidy czy DNA [6,48,57,95].
BIOCHEMICZNA I MOLEKULARNA CHARAKTERYSTYKA KATALAZY (CAT)
Historia katalazy jest dobrze udokumentowana w literaturze naukowej. Pierwsze informacje na temat jej właściwości biochemicznych pojawiły się w 1900 r. [70]. W badaniach wykorzystywano katalazę pochodzącą z różnych źródeł. Materiałem biologicznym były zarówno organizmy roślinne jak i zwierzęce, ponadto bakterie i drożdże (np. tytoń, drożdże, krew, wątroba bydlęca, erytrocyty ludzkie i inne tkanki) [71,72,75,96,114]. Dziś katalaza to enzym o najlepiej poznanej strukturze chemicznej. Po raz pierwszy (1937 r.) została ona wyizolowana z wątroby wołowej [105]. Od tego czasu katalaza była przedmiotem licznych badań i jednym z pierwszych enzymów, które zostały oczyszczone [28]. W dalszym ciągu katalaza i jej funkcje są obiektem zainteresowań i intensywnych badań.
WYSTĘPOWANIE
Katalaza (oksydoreduktaza nadtlenek wodoru: nadtlenek wodoru, EC 1.11.1.6) występuje w komórkach zwierzęcych i roślinnych. Jej obecność wykazano również w komórkach bakteryjnych. Większość bakterii beztlenowych (oprócz Propionibacteria shermanii) i niektóre z bakterii tlenowych np. Baccillus popillie, Mycoplasma pneumonice nie ma tego enzymu [64,102,108,114,120].
U Procaryota katalaza jest umiejscowiona w przestrzeni międzybłonowej, pomiędzy wewnętrzną i zewnętrzną błoną komórkową [9,71]. Z kolei w komórkach Eukariota katalaza jest umiejscowiona głównie w peroksysomach, gdzie występuje z innymi enzymami klasy oksydoreduktaz np. oksydazą L-aminokwasów, oksydazą a-hydroksykwasów, oksydazą acylokoenzymu A, oksydazą moczanową i innymi [16,49,101,102]. W peroksysomach uczestniczy w unieczynnianiu H2O2 – ubocznego produktu utleniania kwasów tłuszczowych. W peroksysomach wątroby ssaków katalaza może stanowić do 16% wszystkich białek. Jednoczesna obecność w peroksysomach enzymów, które generują H2O2 i enzymu odpowiedzialnego za jego rozkład jest biologicznie korzystne [9,24,27,67].
H2O2 powstaje również w komórkach w reakcjach katalizowanych przez oksydazę ksantynową oraz mitochondrialny i mikrosomalny system transportujący elektrony [16,27,52].
Nieduże ilości katalazy wykazano w mitochondriach (np. wątroby) oraz retikulum endoplazmatycznym. Ponieważ nadtlenek wodoru in vivo może łatwo dyfundować przez błony, nieznaczne jego ilości mogą się pojawiać również w cytosolu [7,92,118].
W komórkach S. cerevisiae i E. coli występują 2 typy katalazy: typ A (katalaza atypowa, peroksysomalna) i typ T (katalaza typowa, cytoplazmatyczna) [24,53,75].
Największą aktywność katalazy wykazano w wątrobie, nerkach, krwi (erytrocyty), szpiku, błonach śluzowych, podczas gdy jej najmniejszą aktywność stwierdzono w tkance łącznej [4,9,32,56,77,101].
Obecność katalazy w peroksysomach komórek organizmu zapewnia im ochronę przed toksycznym działaniem nadtlenku wodoru i innych pochodnych. Jest to możliwe, ponieważ przekształca ona H2O2 do tlenu cząsteczkowego i wody, bez wytwarzania wolnych rodników, a powstający w tej reakcji tlen jest wykorzystany w innych przemianach metabolicznych [12,18,25,100].
Katalaza pełni szczególną rolę w metabolizmie erytrocytów, które są wystawione na działanie dużych stężeń tlenu. Wraz z innymi enzymami antyoksydacyjnymi (peroksydazą i reduktazą glutationową) oraz reduktazą methemoglobiny ochrania ona erytrocyty przed skutkami stresu oksydacyjnego [20,32,47,77]. Ma to szczególne znaczenie w przypadku erytrocytów, ponieważ są one narażane na stosunkowo duże stężenia ponadtlenków i nadtlenków. Ponadtlenki powstają w erytrocytach w wyniku autooksydacji hemoglobiny (Hb) do methemoglobiny (metHb), która nie ma zdolności przenoszenia tlenu. W ludzkich erytrocytach, około 3% Hb ulega w ciągu 24 godzin takiemu przekształceniu. Zaobserwowano, że ilość methemoglobiny, powstającej w erytrocytach eksponowanych na działanie nadtlenku wodoru, jest odwrotnie proporcjonalna do aktywności katalazy [4,32,47].
BUDOWA GENU CAT
W latach 80. XX wieku dzięki nowoczesnym technikom molekularnym (zastosowanie zrekombinowanych fagów; recombinant phage clones) udało się poznać budowę i lokalizację ludzkiego genu katalazy.
Gen ten znajduje się na chromosomie 11 (ramię 13) [63,88,116]. Składa się z 13 eksonów, które oddzielone są od siebie przez 12 intronów. Długość całego genu szacuje się na 34 kpz, w tym region kodujący stanowi 1581 pz. Znana jest wielkość i lokalizacja poszczególnych eksonów, wchodzących w skład genu katalazy. Wielkość intronów mieści się w granicach 400 pz do 10.5 kpz. Największy z nich jest umiejscowiony pomiędzy eksonami 1 i 2 [88,98].
Ekson 13 zawiera kodony dla 21 aminokwasów C-końca białka katalazy, kodon TGA kończący translację białka oraz fragment o długości 628 pz, który nie ulega translacji [10,87,88].
N-końcową resztę Ala w białku katalazy koduje trójka nukleotyów GCT, znajdująca się bezpośrednio za kodonem ATG, któremu przypisuje się udział w inicjacji translacji. Rejon długości 368 pz powyżej kodonu ATG nie zawiera jego powtórzeń. Jest on natomiast otoczony sekwencją ACGCTATGC i ściśle jej odpowiadającą sekwencją zachowawczo zgodną – sekwencja „konsensus” (consensus sequences) CC(A/G)CCATG(G), które różnią się między sobą tylko dwoma nukleotydami i są uważane za kodony inicjacji translacji [10,98,116].
W przeciwieństwie do większości genów, transkrybowanych przez polimerazę RNA II, gen katalazy jest pozbawiony w regionie promotora sekwencji TATA (TATA box). Prawdopodobnie polimeraza II RNA kieruje się do promotora dzięki sekwencji inicjatorowej w miejscu startu transkrypcji, których w genie katalazy jest 8 [58,63,66].
Powyżej miejsc startu transkrypcji znajdują się sekwencje CCAAT (CCAAT box) i GGGCGG (GC box). Sekwencje CCAAT znajdują się w pozycji –97, –126, –229 powyżej miejsc startu transkrypcji genu katalazy. Przy braku TATA w genie katalazy sekwencje te prawdopodobnie uczestniczą w wiązaniu czynników transkrypcyjnych [58,85,87].
Powtórzenia sekwencji GGGCGG i jej dokładnej odwrotności CCGCCC, występujące w pozycji –71, –281,–314 powyżej miejsca startu transkrypcji, tworzą regiony bogate w pary GC, które prawdopodobnie są częścią promotora genu katalazy. Obecność GC box stwierdzano w promotorach wielu innych genów. Z regionami bogatymi w GC oraz ich sekwencjami „konsensus” wiąże się konstytutywny czynnik transkrypcyjny SP1 [58,85,88]
Sygnał poliadenylacji (5’-AATAAA-3’) w genie katalazy znajduje się w odległości 18 pz powyżej miejsca startu poliadenylacji. Poniżej miejsca sygnału poliadenylacji występują często sekwencje bogate w T (od 10 do 12 reszt T) [85,87].
BUDOWA LUDZKIEJ KATALAZY
Intensywne badania przeprowadzone w latach 80–90. XX wieku w wielu ośrodkach badawczych dostarczyły szczegółowej wiedzy na temat budowy katalazy, pochodzącej od wielu gatunków organizmów. Wszystkie enzymy mają wspólny schemat budowy, różnią się liczbą podjednostek i strukturą poszczególnych domen budujących białko [17,29,71]. Najlepiej opisana w literaturze naukowej jest budowa katalazy wołowej, natomiast w przypadku organizmu ludzkiego, katalaza pochodząca z erytrocytów [30,65,78,86,90,111].
Całkowita masa cząsteczkowa katalazy u ssaków waha się w granicach 220–350 kDa. W przypadku wątrobowej katalazy wołowej i katalazy ludzkiej z erytrocytów wynosi ona 240 kDa. Ludzka katalaza ma budowę podjednostkową. Jest homotetramerem, zbudowanym z czterech identycznych podjednostek, każda o masie cząsteczkowej 60 kDa [25,28,79,93,97].
Każdą podjednostkę tworzą cztery domeny polipeptydowe. Strukturę pierwszorzędową monomeru katalazy stanowi łańcuch polipeptydowy zbudowany z 526 aa z grupą hemową oraz cząsteczką NADPH. Strukturę II rzędową katalazy tworzą regularnie powtarzające się motywy a
-helisy i b
-kartki; dominuje a
-helisa. W obrębie regularnych struktur drugorzędowych, w pojedynczych miejscach pojawiają się regiony nieuporządkowane, którym przypisuje się istotne znaczenie w utrzymaniu całego tetrameru. Mocno hydrofobowe wnętrze pojedynczego monomeru katalazy jest utworzone przez ośmiokrotnie skręcone przeciwrównoległe b
-cylindry (b
1–8). Tworzą one dwie powierzchnie, obejmujące reszty aminokwasowe od Met 339 do Ala 345 oraz koniec cylindra b
2, wchodzący klinem pomiędzy cylindry b
4 i b
5. N-koniec monomeru tzw. „ramię wiążące” utworzone jest przez reszty aminokwasowe 5-70. Łączy ono w dość skomplikowany sposób dwie podjednostki katalazy, tworząc połączenie z sąsiadującą podjednostką poprzez jej długą wstęgowatą pętlę, zbudowaną z reszt aminokwasowych 380-438. Helikalne domeny na pojedynczej powierzchni b
-cylindra są utworzone przez cztery C-końcowe helisy (a
16, a
17, a
18, a
19) oraz przez cztery helisy utworzone przez reszty zlokalizowane pomiędzy cylindrem b
4 i b
5 (a
4, a
5, a
6, a
7) a cząsteczką NADPH [8,17,78,79,113].
Dimery katalazy, powstające w wyniku wymiany „ramion wiążących”, łączą się w pary i tworzą tetramer. Połączenie to ma podstawowe znaczenie, ponieważ „ramiona wiążące”, pochodzące od jednego dimeru, tworzącego tetramer katalazy, odpowiadają za osłonę hemu w miejscu aktywnym umiejscowionym w drugim dimerze tego tetrameru. Sugeruje to, że struktura przestrzenna katalazy może mieć bardzo duży wpływ na funkcjonowanie tego enzymu. Struktura ludzkiej katalazy, a także katalaz innego pochodzenia wskazuje, że tetrameryzacja jest procesem niezbędnym do prawidłowego funkcjonowania tego enzymu. Istotne znaczenie ma również sposób połączenia podjednostek w tetramer [65,86,110,120].
Zaobserwowano, że wymiana „ramion wiążących” podczas tworzenia tetrameru zachodzi tylko między określonymi dimerami katalazy, co sugeruje ich biologiczne przyporządkowanie względem siebie. Połączenie 4 monomerów katalazy stanowi strukturę czwartorzędową enzymu. Organizacja kompleksu multimerycznego jest bardziej skomplikowana niż ułożenie pojedynczych monomerów. Obejmuje ona zmiany w pofałdowaniu struktury każdego z monomerów, których występowanie ma na celu optymalizacje interakcji pomiędzy podjednostkami. Najbardziej zaangażowane w oddziaływania między podjednostkami są ramiona utworzone przez końcowe aminokwasy „ramion wiążących”. Najbardziej elastyczna część cząsteczki jest w ten sposób odpowiedzialna za większość strukturalnych interakcji między podjednostkami enzymu. Wskutek oddziaływań między podjednostkami domena utworzona przez końcowe aminokwasy jest prawie całkowicie ukryta między sąsiadującymi podjednostkami tetrameru. Z kolei oddziaływanie sąsiadujących ze sobą par „ramion wiążących” prowadzi do wytworzenia struktur o charakterze przeciwrównoległej b
-kartki. Pomiędzy podjednostkami występują również liczne wiązania utworzone w większości przypadków przez argininę, asparaginę i kwas glutaminowy [65,86,109,111].
Prawidłowa tetrameryzacja zapewnia odizolowanie hemu w miejscu aktywnym od reszty enzymu, co z kolei pozwala na prawidłowy przebieg reakcji katalizowanej przez enzym. Brak takiej izolacji hemu od środowiska, w którym działa katalaza, mógłby bowiem prowadzić do powstawania rodnika hydroksylowego.
Badania nad ludzką katalazą wykazały, że jest ona białkiem mocno uwodnionym. Woda pełni funkcję „wypełniacza” pomiędzy czterema domenami polipeptydowymi pojedynczej podjednostki katalazy, a także pomiędzy podjednostkami tetrameru katalazy. Tylko hydrofobowe b-cylindry i bezpośrednie otoczenie miejsca aktywnego są całkowicie odizolowane od cząsteczek wody [29,86,93].
HEM W STRUKTURZE KATALAZY
Każda z podjednostek katalazy ma układ hemowy z położonym centralnie atomem żelaza (Fe3+). Grupa hemowa jest osadzona głęboko w każdej z podjednostek katalazy. W przypadku monomerów katalazy ludzkiej hem jest wbudowany do wnętrza hydrofobowego kanału o długości 30A. Te wąskie kanały uniemożliwiają dostęp do hemu cząsteczkom większym niż H2O2. Szczegółowe badania wykazały, że cząsteczka hemu jest ułożona pionowo w stosunku do b
-cylindrów b
2, b
3 i b
4, pomiędzy a
-helisami (a
4 i a
12) jednej podjednostki pierwszego dimeru a helisą a drugiej podjednostki drugiego dimeru [11,29,120].
Żelazo w miejscu aktywnym katalazy jest związane pięcioma wiązaniami koordynacyjnymi. Cztery z nich to wiązania o długości ~1,9A, utworzone z azotem umiejscowionym w strukturze hemu. Piąte wiązanie koordynacyjne żelazo tworzy z tyrozyną w pozycji 358 łańcucha polipeptydowego białka katalazy. Tyr (358) jest ustawiona dokładnie w kierunku jonu żelaza. Odległość między atomem Fe a grupą fenolową Tyr wynosi 1,85 A. Taki układ prawdopodobnie odpowiada za deprotonację tlenu grupy fenolowej tyrozyny wywołaną siłą elektronową atomu żelaza. Część fenolowa Tyr pełni funkcję liganda żelaza układu hemowego. Oddziaływanie fenolowych reszt Tyr z żelazem w centrum aktywnym katalazy jest ważnym czynnikiem decydującym o preferowaniu jako substratów związków o charakterze nadtlenków. Heterolityczny rozkład wiązań O–O zapobiega powstawaniu rodnika hydroksylowego i jest niezbędny do funkcjonowania katalazy [29,60,65,79,111].
Oddziaływanie reszty Tyr (358) z resztami argininy (Arg 72, Arg 117 i Arg 365) zapewnia właściwą lokalizację hemu w strukturze enzymu i pomaga w utrzymaniu wysokiego potencjału redox podczas przebiegu reakcji katalizowanej przez katalazę. Oprócz ujemnie naładowanej reszty Tyr układ ten jest stabilizowany przez reszty Tyr 358, Arg 354, His 218 i Asp 348. Otoczenie atomu Fe w hemie ulega dynamicznym zmianom tak, aby utrzymać każde miejsce aktywne w podjednostkach katalazy w stanie gotowości do unieczynniania H2O2 [12,25,29,90,93].
ROLA NADPH
Ludzka katalaza zawiera cztery ściśle połączone z nią cząsteczki NADPH. Na każdy monomer przypada jedna cząsteczka NADPH [61]. W każdej podjednostce znajduje się miejsce przyłączenia NADPH. Ma ono postać zagłębienia na powierzchni cząsteczki katalazy, zlokalizowanego pomiędzy domeną heliakalną a b
-cylindrem, przy czym atom C4 aktywnego nikotynamidu cząsteczki NADPH położony jest w odległości 19A od najbliższej grupy hemowej. NADPH jest zwinięty wokół łącznika fosforanowego, co umożliwia niezbędne oddziaływania międzycząsteczkowe i odizolowanie aktywnego pierścienia nikotynamidowego od części białkowej enzymu, a także adenozyny [65,86,93,110].
Katalaza ma większe powinowactwo do NADPH niż NADH, co jest związane z obecnością reszty fosforanowej, biorącej udział w ważnych interakcjach w procesie łączenia się z białkiem katalazy. Pierścień nikotynamidowy jest upakowany w przestrzeni o charakterze niepolarnym pomiędzy resztami Val 302 i Pro 151 oraz cukrem adenozyny. Niepolarny charakter tego miejsca prawdopodobnie zabezpiecza przed przyłączaniem słabo wiązanych NADP+ i NAD+. Miejsce to nie zawiera grup, które byłyby dobrymi akceptorami protonów pochodzących od węgla C4 nikotynamidu, co może mieć funkcjonalne znaczenie w przypadku utlenienia NADPH przez pozostałą (pozostającą w spoczynku) część enzymu [30,60,61].
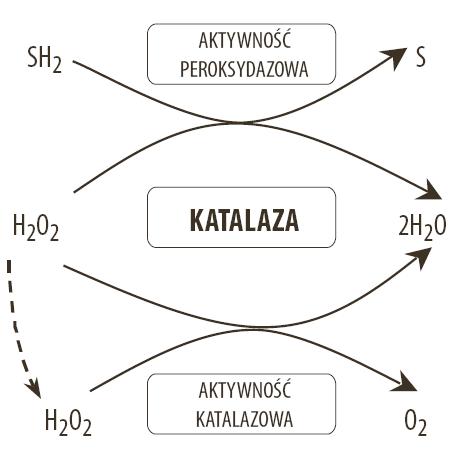
NADPH przypisuje się szczególną rolę w działaniu katalazy. Ten zredukowany nukleotyd obniża podatność enzymu na inaktywację wywołaną toksycznym działaniem nadtlenku wodoru. Ponadto stwierdzono, że katalaza ludzka i wołowa ma zdolność zarówno przyłączania, jak i uwalniania NADPH. Wskazuje to, że w komórkach narażonych na stres oksydacyjny katalaza przez uwalnianie NADPH może pełnić funkcje regulatorowe. Przypuszcza się, że uwalnianie NADPH z katalazy może zwiększyć unieczynnianie H2O2. Jest to możliwe dzięki mechanizmom związanym z działaniem peroksydazy glutationowej i reduktazy glutationowej, które wykorzystują w reakcji regeneracji GSH z GSSG cząsteczkę NADPH (ryc.2) [5,29,59,111].

Ryc. 2. Rola katalazy w rozkładzie H2O2; H2O2 – nadtlenek wodoru; SH2 – związek chemiczny – donor wodoru; S – utleniony związek chemiczny
Funkcja NADPH połączonego z katalazą nie jest do końca wyjaśniona. Istnieją na ten temat różne hipotezy:
• NADPH może zapobiegać, a nawet częściowo odwracać, inaktywację katalazy przez jej własny substrat – H2O2. Potwierdzają to badania Eatona i wsp. [26], według których NADPH jest jednym z endogennych czynników zaangażowanych w utrzymanie, a nawet w odzyskiwanie utraconej aktywności katalazy, dzięki stabilizowaniu określonej struktury tego enzymu.
• Katalaza, uwalniając NADPH, wpływa regulacyjnie na układ enzymów glutationozależnych: peroksydaza glutationowa/reduktaza glutationowa sprawia, że układ ten jest bardziej wydajny w warunkach stresu oksydacyjnego. NADPH potrzebny do regeneracji GSH z GSSG powstaje w cyklu pentozofosforanowym, w wyniku działania dehydrogenazy glukozo-6-fosforanowej. W warunkach nasilonego stresu oksydacyjnego w ludzkich erytrocytach, gdy ilość NADP+ niezbędna do wytwarzania NADPH jest ograniczona i dochodzi do spadku stężenia NADPH, katalaza może być dawcą NADPH. Jest to możliwe dzięki temu, że związek ten może być zastąpiony w katalazie przez NADH. Katalaza stanowi w erytrocytach magazyn NADP+, jego stężenie wynosi około 11–12 µM NADP, co stanowi 1/3 całkowitej zawartości tego związku w erytrocytach [29,60,61,77,120].
Regulacyjny wpływ NADPH na działanie enzymów glutationozależnych ma szczególne znaczenie przy założeniu, że to nie katalaza, ale peroksydaza i reduktaza glutationowa mają podstawowe znaczenie w usuwaniu H2O2 w erytrocytach ludzkich, na co wskazują badania [5,19,20,91].
OGÓLNY MECHANIZM REAKCJI KATALIZOWANEJ PRZEZ CAT
• Ostatnia koncepcja zakłada, że NADPH jest substratem lub kofaktorem enzymatycznej aktywności katalazy, co może mieć związek z innym mechanizmem jej działania niż został poznany i opisany [61].
Katalaza to enzym o podwójnej aktywności, wykazuje ona aktywność katalazową i peroksydazową.
Podstawowa funkcja katalazy w komórkach to udział w reakcji dysproporcjonowania nadtlenku wodoru:

Wobec niektórych związków chemicznych katalaza wykazuje również aktywność peroksydazową. Katalizuje ona reakcję utleniania etanolu, metanolu, mrówczanu, azotynów, chinonów i innych. W reakcji tej nadtlenek wodoru jest substratem, który jest redukowany do wody, przez związki będące donorami wodoru (SH2) [13,32,49,84,100,106].

Katalaza jest enzymem reagującym bardzo szybko z nadtlenkiem wodoru. W ciągu 1 minuty katalaza rozkłada do wody i tlenu około 6 mln cząsteczek tego związku. Stała szybkości katalizowanej przez katalazę reakcji wynosi 1,7×107×1 mol–1xs–1 [12,14,48,73].
Przekształca ona dwie cząsteczki nadtlenku wodoru do dwóch cząsteczek wody i jednej cząsteczki tlenu. Reakcja ta przebiega dwuetapowo [13,69,113].
Szczegółowa analiza katalitycznego rozkładu nadtlenku wodoru z udziałem katalazy pozwoliła dokładnie poznać każdy z etapów tej reakcji.
I etap

W pierwszym etapie reakcji katalazy (Fe (III) – CAT) z nadtlenkiem wodoru następuje redukcja nadtlenku wodoru do wody z udziałem Fe3+ układu hemowego katalazy, z utworzeniem związku I (Fe (V) – CAT), w którym żelazo hemu jest na 5 stopniu utlenienia. Powstawanie tego związku po raz pierwszy opisali i wyjaśnili mechanizm tej reakcji Chance i wsp. [12,16,59,84].
II etap
W drugim etapie następuje utlenienie przez związek I kolejnej cząsteczki nadtlenku wodoru, w wyniku czego powstaje tlen cząsteczkowy i woda.

Związek I powraca do stanu wyjściowego, w którym żelazo hemu jest na 3 stopniu utleniania [17,59].
Dzięki zastosowaniu nowoczesnych metod badawczych udało się ustalić, że w I etapie reakcji nadtlenek wodoru przez wąski hydrofobowy kanał trafia do centrum aktywnego katalazy, gdzie znajduje się hem. Taki sposób dostarczania substratu do centrum jest bardzo korzystny ponieważ uniemożliwia wejście substratów większych niż nadtlenek wodoru. Dochodzi do wzajemnych oddziaływań pomiędzy resztami His (w pozycji 75) i Asn (w pozycji 148) [17, 29]. Prowadzą one do pewnych przesunięć w obrębie cząsteczki nadtlenku wodoru. Jeden z protonów (H) zostaje za pośrednictwem reszty His (75) przeniesiony z jednego atomu tlenu na drugi. Prowadzi to do wydłużenia i polaryzacji wiązania O–O. Tak przekształcona cząsteczka H2O2 jest przyłączana do żelaza w centrum hemu. Połączenie to powoduje rozerwanie spolaryzowanego wiązania O–O, co prowadzi do uwolnienia cząsteczki wody i wytworzenia tzw. kompleksu I (Fe (V)=O). Przyłączenie pierwszej cząsteczki H2O2 otwiera hydrofobowy kanał i prawdopodobnie ułatwia przyłączenie kolejnej cząsteczki H2O2
, co z kolei być może zapobiega rozpadowi kompleksu I [17,59,61,120].
Podczas drugiego etapu reakcji Fe (V)=O reaguje z drugą cząsteczką nadtlenku wodoru, czemu towarzyszą – podobne jak w etapie pierwszym – przejścia protonów i dwóch elektronów. Prowadzi to do utworzenia wyjściowej postaci żelaza Fe (III) – CAT, cząsteczki wody i tlenu cząsteczkowego [79,86].
Reaktywność hemu uwarunkowana jest obecnością reszty Tyr (357), która tworzy piąte wiązanie koordynacyjne z żelazem w jego centrum. Połączenie to wspomaga utlenianie Fe (III) do Fe (IV) oraz bierze udział w usuwaniu z pierścienia hemowego elektronu. Skuteczność działania katalazy w tej reakcji zależy od oddziaływania His (75) i Asn (148) z intermediatami reakcji. Proponowany mechanizm reakcji katalizowanej przez CAT znajduje potwierdzenie w wynikach eksperymentalnych badań, w których modyfikowano resztę His (75) z użyciem 3-amino-1,2,4-triazolu, co powodowało hamowanie reakcji (brak przyłączania substratu) [23,79,86].
W celu heterolitycznego rozszczepienia nadtlenku wodoru, który charakteryzuje się budową symetryczną, katalaza wykorzystuje dwa rodzaje asymetrycznej interakcji z substratem. Heterolityczny rozpad nadtlenku wodoru jest możliwy dzięki jednoczesnemu oddziaływaniu z bogatym w elektrony jonem żelaza i pełniącymi rolę biorców elektronów resztami His 75 i Asn 148. Właściwa geometria wiązań koordynacyjnych jonu żelaza, jak i tworzenie połączenia z nadtlenkiem (kompleks I), zapewnia nie tylko elastyczność tego połączenia, ale również nasila reakcję rozszczepienia nadtlenku. Utworzenie kompleksu I jest prawdopodobnie związane z neutralizacją ładunku reszty Arg 354, która zachodzi dzięki przepływowi elektronów. Neutralizacja reszty argininy 354 ma na celu zarówno zmniejszenie ładunku odpychającego w stosunku do kationowej porfiryny, jak i zmniejszenie polaryzacji w wykazującym niedobór elektronów układzie Tyr 358 – Fe4+=O [59,60,97,110].
Potencjał redox kompleksu I jest modyfikowany bezpośrednio przez reszty Phe 153 i Phe 161 oraz przez tworzenie wiązań o charakterze soli pomiędzy resztami karboksylowymi hemu i resztami arginin w pozycjach 72, 112 i 365.
Przywrócenie funkcji katalitycznej katalazy (powrót do postaci wyjściowej) jest możliwe dzięki utlenieniu następnej cząsteczki nadtlenku wodoru. Ponieważ reakcja ta obejmuje przenoszenie elektronów, redukcja w zasadzie może zachodzić na odległość. Cząsteczka nadtlenku wodoru jest najprawdopodobniej pobierana przez hydrofobowy kanał, gdzie przyłącza się poprzez jeden z atomów tlenu do reszt His 75 i Asn 148. Drugi atom tlenu wchodzący w skład cząsteczki H2O2 układa się w taki sposób, aby zapewnić potencjał wzdłuż kanału i zredukować kompleks I w wyniku stopniowego transportu dwóch elektronów i protonu. Inne potencjalne substraty katalazy, takie jak np. etanol, mogą dzięki obecności grup hydroksylowych łączyć się podobnie jak nadtlenek wodoru z resztą His 75 w kanale hydrofobowym [59,79,86,120].
Kiedy reakcja kompleksu I z odpowiednim reduktorem zostaje wstrzymana, katalaza utlenia przyłączone NADPH wykorzystując go jako reduktor. W sytuacji niedostępności/ braku NADPH alternatywną przemianą kompleksu I wydaje się jednoelektronowa redukcja z wykorzystaniem elektronu pochodzącego z Tyr 370 [29,53,61,110,120].
W przypadku aktywności peroksydazowej katalazy kompleks I reaguje z donorami wodoru, np. etanolem czy metanolem, które w obecności tlenu pochodzącego z cząsteczki H2O2 są utleniane:

Zaobserwowano, że in vivo katalaza może również utleniać (NO2–) do (NO3–) [14,16,52,59].
Aktywność CAT zależy od wielu czynników, m.in. stężenia substratu, temperatury, pH, obecności aktywatorów i inhibitorów oraz pochodzenia enzymu. Inne warunki działania będą charakterystyczne dla katalazy pochodzenia roślinnego (np. z ziemniaka), a inne dla katalaz eukariotycznych czy bakteryjnych. Eukariotyczne katalazy są aktywne w szerokim zakresie pH (5,0–10,5). Za najbardziej optymalne pH dla katalazy uważa się jednak pH 7,0 [12,108,113].
Aktywność katalazy jest hamowana przez wiele związków chemicznych. Niekompetycyjnym inhibitorem aktywności katalazy jest siarczan miedzi, natomiast kompetycyjnym cyjanek. Hamowanie aktywności katalazy przez cyjanki zostało wykorzystane w 1923 r. do badań, które pozwoliły wykazać obecność w strukturze enzymu żelaza. Bardzo swoistym inhibitorem katalazy okazał się również 3-amino- 1,2,4 –tetriazol [73,86].
AKTYWNOŚĆ KATALAZY A CHOROBY
Katalaza jest enzymem, który pełni funkcje zarówno w katabolizmie nadtlenku wodoru, jak i w utlenianiu egzogennych substratów, takich jak metanol, etanol i inne [13,52,77,120]. Przypisuje się jej szczególną rolę w stanach zapalnych [49], mutagenezie i kancerogenezie [1], ochronie przed apoptozą [76,94,117]. Obniżenie aktywności katalazy towarzyszy wielu chorobom, m.in. zapaleniu płuc, gruźlicy, miażdżycy, cukrzycy, żółtaczce, nowotworom, chorobom neurodegeneracyjnym (chorobie Parkinsona, Alzheimera), zapaleniu nerek i innym [15,56,74,82,83].
Wszystkim tym chorobom towarzyszą stany zapalne. Zaobserwowano, że w przypadku chorób poprzedzonych stanami zapalnymi wzrasta aktywność katalazy w osoczu krwi. Jest to spowodowane najprawdopodobniej jej wyciekiem z uszkodzonych komórek, zwłaszcza z erytrocytów [41,80,82]. Wzrost aktywności katalazy w przebiegu zapalenia wydaje się mieć charakter reakcji obronnej organizmu [119].
Małą aktywność katalazy we krwi obserwowano u chorych z miażdżycą i cukrzycą, co wskazuje na długo trwający stres oksydacyjny w komórkach organizmu tych osób. Wykazano, że H2O2 uszkadza komórki b
trzustki, co przy niedoborze katalazy prowadzi do rozwoju cukrzycy [39,43,46,52].
Wyraźne obniżenie aktywności obserwowano w wielu typach nowotworów (głowy i szyi, płuc, przewodu pokarmowego, piersi, nerek czy białaczkach) [15,21,22,103,107].
Brak lub niedobór katalazy we krwi i innych tkankach organizmu prowadzi u ludzi do powstawania akatalasemii i hipokatalasemii. Brak katalazy (akatalasemia) lub jej niedobór (hipokatalasemia) powodują m.in. owrzodzenia jamy ustnej. Zmiany te, bardzo wcześnie, bo już w wieku 13–19 lat prowadzą u chorych do utraty zębów [26,80,81,109,115].
AKATALASEMIE
Po raz pierwszy w 1946 roku, Takahara i Miyamoto odkryli całkowity brak katalazy (akatalasemia) w krwinkach czerwonych Japończyków, który objawiał się powtarzającymi się stanami zapalnymi dziąseł i owrzodzeniami w obrębie jamy ustnej (choroba Takahara) [109].
Powstawanie akatalasemii i hipokatalasemii jest uwarunkowane genetycznie i związane z mutacjami w genie katalazy, które dziedziczy się w sposób autosomalny, recesywny. U homozygot stwierdzano całkowity brak katalazy (akatalasemia), natomiast u heterozygot obserwowano 50% obniżenie aktywności katalazy w porównaniu z jej poziomem u osób zdrowych (hipokatalasemia) [55,81,112].
Częstotliwość występowania zmutowanego genu katalazy w Japonii jest stosunkowo wysoka i wynosi w przypadku akatalasemii 0,8/1000, a hipokatalasemii 2–4/1000. Zaobserwowano, że np. w Hiroszimie i Nagasaki częstotliwość występowania heterozygot wynosi 0,09/1000 i 1,4/1000 w innych częściach Japonii [51,54,62]. Występowanie akatalasemii poza Japonią wykazano m.in. w Izraelu, Szwajcarii, na Węgrzach. Najwięcej przypadków akatalasemii stwierdzono w Japonii (91 pacjentów z 46 rodzin) oraz w Szwajcarii (11 pacjentów z 3 rodzin) [2,3,36,42,50,81].
W Szwajcarii i Izraelu u homozygot obserwowano śladową aktywność katalazy, co wskazuje na inny typ mutacji w genie katalazy od tych, które są odpowiedzialne za powstawanie akatalasemii w Japonii [3,51]. Niestety w piśmiennictwie brak jest bardziej szczegółowych danych na ten temat.
W przypadku typu japońskiego akatalasemii, patologia ta powstaje w wyniku dwóch różnych mutacji w genie katalazy. Pierwszy typ mutacji (Japonese type A) to substytucja guaniny na adeninę (G->A) w 5 nukleotydzie 4 intronu, która powstaje w czasie jego wycinania (splicing). Mutacja ta określana jako mutacja składania mRNA (splicing mutation) jest odpowiedzialna za obniżenie aktywności katalazy, bez biochemicznych różnic między produktem białkowym prawidłowego i zmutowanego genu katalazy [62,115].
Drugi typ mutacji (Japonese type B) jest delecją tyminy z pozycji 358 eksonu 4, która prowadzi do zmiany ramki odczytu (frameshift mutation) i wprowadzenia kodonu terminacji TGA, kończącego wcześniej syntezę łańcucha polipeptydowego katalazy. Produktem jest skrócone, niestabilne i niewykazujące aktywności katalitycznej białko składające się z 133 reszt aminokwasowych [54].
Rodzinny niedobór katalazy stwierdzono również u 13 rodzin węgierskich. Częstotliwość wystąpienia akatalasemii i hipokatalasemii na Węgrzech wynosiła odpowiednio 0,05/1000 i 2,3/1000. Badania molekularne prowadzone przez Gotha i jego zespół w zakresie identyfikacji mutacji w genie katalazy, które są przyczyną powstawania akatalasemii i hipokatalasemii u ludzi na Węgrzech wykazały występowanie 3 nowych mutacji, innych od tych występujących w populacji japońskiej. Zastosowanie w badaniach metody sekwencjonowania, analizy polimorfizmu sekwencji mikrosatelitarnych oraz badanie ekspresji mutacji pozwoliły na dokładne określenie typu mutacji punktowych pojawiających się w genie katalazy typowych tylko dla populacji węgierskiej [33,38,40].
Mutacja węgierska A, wywołująca akatalasemie jest mutacją typu zmiany ramki odczytu na skutek insercji GA w pozycji 138 eksonu 2, co objawia się wcześniejszym pojawieniem kodonu stop TGA i syntezą skróconego do 133 AA białka katalazy [44]. Mutacja węgierska B, która jest odpowiedzialna za hipoktalasemie jest również mutacją typu ramki odczytu powstającą po insercji G w pozycji 79 eksonu 2, co prowadzi do powstawania białka zbudowanego tylko z 58 AA [37]. Z kolei mutacja węgierska typu C jest również odpowiedzialna za hipokatalasemie i powstaje jako wynik substytucji T->G w pozycji 5 intronu 7 i objawia się wyraźnym obniżeniem aktywności katalitycznej katalazy [42]. Węgierskie typy A, B i C niedoboru katalazy stwierdzono u 65% poddanych badaniom pacjentów na Węgrzech. Nie stwierdzono ich występowania w Japonii [37,44,45,112].
W badaniach prowadzonych przez Gotha na populacji węgierskiej w przypadku akatalasemii i hipokatalasemii stwierdzono polimorfizm 5’-końca genu katalazy. Wykazano mutacje A->T i C->A i C-> T w pozycjach -21, -20 i -18 powyżej miejsca inicjacji translacji oraz mutacje T->C w pozycji -4, -44-, -49 w regionie niekodującym i C->A w pozycji -18 i -17 w eksonie 1 [34,35,43].
Patogenny charakter tych mutacji prowadzi do częściowej utraty funkcji genu katalazy, co ujawnia się niedoborem lub brakiem prawidłowej postaci białka albo niekorzystnym oddziaływaniem skróconych postaci białka katalazy [42,58].
Akatalasemia jest często związana z występowaniem pewnych schorzeń. Goth i Eaton wykazali w populacji węgierskiej, zwiększoną liczbę chorych na cukrzycę z jednoczesnym dziedzicznym niedoborem katalazy (typ węgierski D) [39,46]. Autorzy sugerują, że długotrwały stres oksydacyjny i dziedziczony autosomalnie recesywnie niedobór katalazy predysponuje do oksydacyjnych uszkodzeń komórek b
trzustki, zaburzeń w wydzielaniu insuliny i rozwoju cukrzycy [43,52].
PODSUMOWANIE
Katalaza jest przykładem szczególnie skutecznie działającego enzymu w komórkach organizmów żywych, chroni je przed skutkami toksycznego działania nadtlenku wodoru. Ma dwie funkcje katalityczne zależne od stężenia H2O2 w środowisku. Przy dużym stężeniu H2O2 w organizmie katalaza usuwa H2O2, przekształcając go do H2O i O2. Kiedy stężenie H2O2 jest małe, a obecne są odpowiednie donory wodoru (np. etanol, metanol) katalaza wykazuje aktywność peraksydazową, usuwa H2O2 powodując jednocześnie utlenienie tych związków. Obniżenie aktywności katalazy występuje w wielu chorobach, którym towarzyszy stres oksydacyjny, szczególnie w chorobach, którym towarzyszą stany zapalne.
Jej niedobór lub brak spowodowany mutacjami w genie, które są dziedziczone w sposób autosomalny recesywny prowadzi do powstania akatalasemii i hipokatalasemii.
PIŚMIENNICTWO
[1] Adachi S., Nagano S., Ishimori K., Watanabe Y., Morishima I., Egawa T., Kitagawa T., Makino R.: Roles of proximal ligand in heme proteins: replacement of proximal histidine of human myoglobin with cysteine and tyrosine by site-directed mutagenesis as models for P-450, chloroperoxidase, and catalase. Biochemistry, 1993; 32: 241-252
[PubMed]
[2] Aebi H., Baggiolini M., Dewald B., Lauber E., Suter H., Micheli A., Frei J.: Observations in two Swiss families with acatalasia. Enzym. Biol. Clin., 1964; 4: 121-151
[PubMed]
[3] Aebi H.E., Wyss S.R.: Molecular hybridization in heterozygotes for Swiss-type acatalasemia. Monogr. Hum. Genet., 1978; 10: 200-204
[PubMed]
[4] Agar N.S., Sadrzadeh S.M., Hallaway P.E., Eaton J.W.: Erythrocyte catalase: a somatic oxidant defense? J. Clin. Invest., 1986; 77: 319-321
[PubMed] [Full Text HTML] [Full Text PDF]
[5] Almarsson O., Sinha A., Gopinath E., Bruice T.C.: Mechanism of one-electron oxidation of NAD(P)H and function of NADPH bound to catalase. J. Am. Chem. Soc., 1993; 115: 7093-7102
[6] Anonymous.: Hydrogen peroxide: A review. IARC. Monographs on the evaluation of carcinogenic risk of humans. 1999; 2: 671-689
[7] Antunes F., Cadenas E.: Estimation of H2O2 gradients across biomembranes. FEBS Lett., 2000; 475: 121-126
[PubMed]
[8] Baker R.D., Cook C.O., Goodwin D.C.: Properties of catalase-peroxidase lacking its C-terminal domain. Biochem. Biophys. Res. Commun., 2004; 320: 833-839
[PubMed]
[9] Bartosz G.: Druga twarz tlenu. Wydawnictwo Naukowe PWN, wyd. II, Warszawa 2003
[10] Bell G.I., Najarian R.C., Mullenbach, G.T., Hallewell R.A.: cDNA sequence coding for human kidney catalase. Nucleic Acids Res., 1986; 14: 5561-5562
[PubMed] [Full Text HTML] [Full Text PDF]
[11] Beyer W.F.Jr., Fridivich I.: Catalases- with and without heme. Basic Life Sci., 1988; 49: 651-661
[PubMed]
[12] Boon E.M., Downs A., Marcey D.: Catalase: H2O2: H2O2 oxidoreductase. Reviews on structure and function of catalases. In: Biomolecules at Kenyon, eds David Marcey, 1997
[13] Cadenas E.: Basic mechanisms of antioxidant activity. Biofactors, 1997; 6: 391-397
[PubMed]
[14] Carpena X., Wiseman B., Deemagarn T., Singh R., Switala J., Ivancich A., Fita I., Loewen P.C.: A molecular switch and electronic circuit modulate catalase activity in catalase-peroxidases. EMBO Rep., 2005; 6: 1156-1162
[PubMed]
[15] Casado A., de la Torre R., Lopez-Fernandez M.E., Carrascosa D., Casado M.C., Ramirez M.V.: Superoxide dismutase and catalase blood levels in patients with malignant diseases. Cancer Lett., 1995; 93: 187-192
[PubMed]
[16] Chance B., Sies H., Boveris A.: Hydroperoxide metabolism in mammalian organs. Physiol. Rev., 1979; 59: 527-605
[PubMed]
[17] Chelikani P., Carpena X., Fita I., Loewen P.C.: An electrical potential in the access channel of catalases enhances catalysis. J. Biol. Chem., 2003; 278: 31290-31296
[PubMed] [Full Text HTML] [Full Text PDF]
[18] Chelikani P., Fita I., Loewen P.C.: Diversity of structures and properties among catalases. Cell. Mol. Life. Sci., 2004, 61: 192-208
[PubMed]
[19] Cohen G., Hochstein P.: Glutathione peroxidase: the primary agent for elimination of hydrogen peroxide in erythrocytes. Biochemistry, 1963; 2: 1420-1428
[PubMed]
[20] Comhair S.A., Erzurun S.C.: The regulation and role of extracellular glutathione peroxidase. Antioxid. Redox Signal., 2005; 7: 72-79
[PubMed]
[21] Czeczot H., Skrzycki M., Gawryszewska E., Podsiad M., Porembska Z.: Evaluation of antioxidant status in patients with primary hepatocellular carcinoma. Pol. Merk. Lek., 2003; 15: 118-122
[PubMed]
[22] Czeczot H., Skrzycki M., Podsiad M., Gawryszewska E., Nyckowski P., Porembska Z.: Antioxidant status of patients with primary colorectal cancer and liver metastases of colorectal cancer. Pol. Merk. Lek., 2005; 18: 58-61
[PubMed]
[23] Darr D., Fridovich I.: Irreversible inactivation of catalase by 3-amino-1,2,4-triazole. Biochem. Pharmacol., 1986; 35: 3642
[PubMed]
[24] De Duve C.: The peroxisome in retrospect. Ann. N. Y. Acad. Sci., 1996; 804: 1-10
[PubMed]
[25] Deisseroth A., Dounce A.L.: Catalase: physical and chemical properties, mechanism of catalysis, and physical role. Physiol. Rev., 1970; 50: 319-375
[PubMed]
[26] Eaton J.W., Ma M.: Acatalasemia. In: Scriver CR, Beaudet AL, Sly WS, Valle D, eds, The Metabolic and Molecular Bases of Inherited Disease. New York: McGraw-Hill, Inc., 1995, 2371-2383
[27] Embley T.M., Martin W.: A hydrogen-producing mitochondrion. Nature, 1998; 396: 517-519
[PubMed] [Full Text PDF]
[28] Eventoff W., Tanaka N., Rossmann M.G.: Crystalline bovine liver catalase. J. Mol. Biol., 1976; 103: 799-801
[PubMed]
[29] Fita I., Rossman M.G.: The active center of catalase. J. Mol. Biol., 1985; 185: 21-37
[PubMed]
[30] Fita I., Silva A.M., Murthy M.R.N., Rossmann, M.G.: The refined structure of beef liver catalase. Acta Cryst., 1986; 42: 497-515
[31] Flohe L.: Glutathione peroxidase: fact and fiction. Ciba Found Symp., 1978, 65: 95-122
[PubMed]
[32] Gaetani G.F., Ferraris A.M., Rolfo M., Mangerini R., Arena S., Kirkman H.N.: Predominant role of catalase in the disposal of hydrogen peroxide within human erythrocytes. Blood, 1996; 87: 1595-1599
[PubMed] [Full Text PDF]
[33] Goth L.: Two cases of acatalasemia in Hungary. Clin. Chim. Acta, 1992; 207: 155-158
[PubMed]
[34] Goth L.: Further genetic heterogeneity in acatalasemia. Electrophoresis, 1997; 18: 1942-1943
[PubMed]
[35] Goth L.: Genetic heterogeneity of the 5′ uncoding region of the catalase gene in Hungarian acatalasemic and hypocatalasemic subjects. Clin. Chim. Acta, 1998; 271: 73-78
[PubMed]
[36] Goth L.: A new type of inherited catalase deficiencies: its characterization and comparison to the Japanese and Swiss type of acatalasemia.Blood Cells Mol. Dis., 2001; 27: 512-517
[PubMed]
[37] Goth L.: A novel catalase mutation (a G insertion in exon 2) causes the type B of the Hungarian acatalasemia. Clin. Chim. Acta, 2001; 311: 161-163
[PubMed]
[38] Goth L., Alizadeh B.N., Sussman H.H.: Further characterisation of Hungarian acatalasemia by Hinf1 polymorphism of catalase gene. Enzyme Protein, 1993; 47: 156-159
[PubMed]
[39] Goth L., Eaton J.W.: Hereditary catalase deficiencies and increased risk of diabetes. Lancet, 2000; 356: 1820-1821
[PubMed]
[40] Goth L., Gorzsas A., Kalmar T.: A simple PCR-heteroduplex screening method for detection of a common mutation of the catalase gene in Hungary. Clin. Chem., 2000; 46: 1199-1200
[PubMed] [Full Text HTML] [Full Text PDF]
[41] Goth L., Lenkey A., Bigler W.N.: Blood catalase deficiency and diabetes in Hungary. Diabetes Care, 2001; 24: 1839-1840
[PubMed] [Full Text HTML] [Full Text PDF]
[42] Goth L., Rass P., Madarasi I.: A novel catalase mutation detected by polymerase chain reaction-single strand conformation polymorphism, nucleotide sequencing, and western blot analyses is responsible for the type C of Hungarian acatalasemia. Electrophoresis, 2001; 22: 49-51
[PubMed]
[43] Goth L., Rass P., Pay A.: Catalase enzyme mutations and their association with diseases. Mol. Diagn., 2004; 8: 141-149
[PubMed]
[44] Goth L., Shemirani A., Kalmar T.: A novel catalase mutation (a GA insertion) causes the Hungarian type of acatalasemia. Blood Cells Mol. Dis., 2000; 26: 151-154
[PubMed]
[45] Goth L., Vitai M.: The effects of hydrogen peroxide promoted by homocysteine and inherited catalase deficiency on human hypocatalasemic patients. Free Radic. Biol. Med., 2003; 35: 882-888
[PubMed]
[46] Goth L., Vitai M., Rass P., Sukei E., Pay A.: Detection of a novel familial catalase mutation (Hungarian type D) and the possible risk of inherited catalase deficiency for diabetes mellitus. Electrophoresis, 2005; 26: 1646-1649
[PubMed]
[47] Guemouri L., Artur Y., Herbeth B., Jeandel C., Cuny G., Siest G.: Biological variability of superoxide dismutase, glutathione peroxidase, and catalase in blood. Clin. Chem., 1991; 37: 1932-1937
[PubMed]
[48] Halliwell B., Clement M.V., Long L.H.: Hydrogen peroxide in the human body. FEBS Lett., 2000; 486: 10-13
[PubMed]
[49] Halliwell B., Gutteridge J.M.C.: Free radicals in biology and medicine., 1999, 3rd ed. New York: Oxford University Press
[50] Hamilton H.B, Neel J.V.: Genetic heterogeneity in human acatalasia. Am. J. Hum. Genet., 1963, 15: 408-419
[PubMed]
[51] Hamilton H.B., Neel J.V., Kobara T.Y., Ozaki K.: The frequency in Japan of carriers of the rare ‘recessive’ gene causing acatalasemia. J. Clin. Invest., 1961; 40: 2199-2208
[PubMed] [Full Text HTML] [Full Text PDF]
[52] Heales S.J.: Catalase deficiency, diabetes and mitochondrial function. Lancet, 2001; 357: 314
[PubMed]
[53] Hillar A., Nicholls P, Switala J., Loewen P.C.: NADPH binding and control of catalase compound II formation: comparison of bovine, yeast and Escherichia coli enzymes. Biochem. J., 1994; 300: 531-539
[PubMed] [Full Text HTML] [Full Text PDF]
[54] Hirono A, Miwa S, Fujii H, Ishida F, Yamada K, Kubota K.: Molecular study of eight Japanese cases of glucose-6-phosphate dehydrogenase deficiency by non-radioisotopic single-strand conformation poly-morphism analysis. Blood, 1994; 83: 3363-3368
[PubMed] [Full Text PDF]
[55] Hirono A., Sasaya-Hamada F., Kanno H., Fujii H., Yoshida T., Miwa S.: A novel human catalase mutation (358 T–>del) causing Japanese-type acatalasemia. Blood Cells. Molecules and Diseases, 1995; 21: 232-234
[PubMed]
[56] Houdou S., Kuruta A., Hasegawa M., Konomi H., Takashima S., Suzuki Y., Hashimoto T.: Developmental immunohistochemistry of catalase in the human brain. Brain Res., 1991; 556: 267-270
[PubMed]
[57] Imlay, J.A., Chin S.M., Linn S.: Toxic DNA damage by hydrogen peroxide through the Fenton reaction in vivo and in vitro. Science, 1988; 240: 640-642
[PubMed]
[58] Jiang Z., Akey J.M., Shi J., Xiong M., Wang Y., Shen Y., Xu X., Chen H., Wu H., Xiao J., Lu D., Huang W., Jin L.: A polymorphism in the promoter region of catalase is associated with blood pressure levels. Hum. Genet., 2001; 109: 95-98
[PubMed]
[59] Kalko S.G., Gelpi J.L., Fita I., Orozco M.: Theoretical study of the mechanisms of substrate recognition by catalase. J. Am. Chem. Soc., 2001; 123: 9665-9672
[PubMed]
[60] Kirkman H.N., Gaetani G.F.: Catalase: a tetrameric enzyme with four tightly bound molecules of NADPH. Proc. Natl. Acad. Sci., 1984; 81: 4343-4347
[PubMed] [Full Text HTML] [Full Text PDF]
[61] Kirkman H.N., Rolfo M., Ferraris A.M., Gaetani G.F.: Mechanisms of protection of catalase by NADPH. Kinetics and stoichiometry. J. Biol. Chem., 1999, 274, 13908-13914
[PubMed] [Full Text HTML] [Full Text PDF]
[62] Kishimoto Y., Murakami Y., Hayashi K., Takahara S., Sugimura T., Sekiya T.: Detection of a common mutation of the catalase gene in Japanese acatalasemic patients. Hum. Genet., 1992; 88: 487-490
[PubMed]
[63] Kittur S.D., Hoppener J.W., Antonarakis S.E., Daniels J.D., Meyers D.A., Maestri N.E., Jansen M., Korneluk R.G., Nelkin B.D., Kazazian H.H.Jr.: Linkage map of the short arm of human chromosome 11: location of the genes for catalase calcitonin, and insulin-like growth factor II. Proc. Nat. Acad. Sci. USA, 1985; 82: 5064-5067
[PubMed] [Full Text HTML] [Full Text PDF]
[64] Klotz M., Klassen G.R., Loewen P.C.: Phylogenetic relationships among prokaryotic and eukaryotic catalases. Mol. Biol. Evol., 1997; 14: 951-958
[PubMed] [Full Text PDF]
[65] Ko T.P., Safo M.K., Musayev F.N., Di Salvo M.L., Wang C., Wu S.H., Abraham D.J.: Structure of human erythrocyte catalase. Acta Crystallogr. D. Biol. Crystallogr., 2000 ; 56: 241-245
[PubMed]
[66] Korneluk R.G., Quan F., Lewis W.H., Guise K.S., Willard H.F., Holmes M.T., Gravel R.A.: Isolation of human fibroblast catalase cDNA clones. Sequence of clones derived from spliced and unspliced mRNA. J. Biol. Chem., 1984; 259: 13819-13823
[PubMed] [Full Text PDF]
[67] Kulikowska-Karpińska E., Moniuszko-Jakoniuk J.: The antioxidant barrier in the organism. Pol. J. Environ. Stud., 2004; 13: 5-13
[Abstract]
[68] Liczmański A.E.: Oxygen toxicity. I. Damage of living cells. Post. Biochem., 1988, 34: 273-291
[PubMed]
[69] Lledías F., Rangel P., Hansberg W.: Oxidation of catalase by singlet oxygen. J. Biol. Chem., 1998; 273: 10630-10637
[PubMed] [Full Text HTML] [Full Text PDF]
[70] Loew O.: Physiological studies of Connecticut leaf tobacco. U.S. Dept. Agri. Repts., 1900; 65: 5-57
[71] Loewen P.C.: Bacterial catalases. Cold Spring Harbor Laboratory Press, 1997, Cold Spring Harbor, NY
[72] Loewen P.C., Carpena X., Rovira C., Ivancich A., Perez-Luque R., Haas R., Odenbreit S., Nicholls P., Fita I.: Structure of Helicobacter pylori catalase, with and without formic acid bound, at 1.6 A resolution. Biochemistry, 2004; 43: 3089-3103
[PubMed]
[73] Loewen P.C., Klotz M.G., Hassett D.J.: Catalase – an “old” enzyme that continues to surprise us. ASM News., 2000; 66: 76-82
[74] Markesbery W.R.: Oxidative stress hypothesis in Alzheimer’s disease. Free Radic. Biol. Med., 1997; 23: 134-147
[PubMed]
[75] Mate M.J., Zamocky M., Nykyri L.M., Herzog C., Alzari P.M., Betzel C., Koller F., Fita I.: Structure of catalase-A from Saccharomyces cerevisiae. J. Mol. Biol., 1999; 286: 135-149
[PubMed]
[76] Miyamoto T., Hayashi M., Takeuchi A., Okamoto T., Kawashima S., Takii T., Hayashi H., Onozaki K.: Identification of a novel growth-promoting factor with a wide target cell spectrum from various tumor cells as catalase. J. Biochem., 1996; 120: 725-730
[PubMed]
[77] Mueller S., Riedel H.D., Stremmel W.: Direct evidence for catalase as the predominant H2O2-removing enzyme in human erythrocytes. Blood, 1997; 90: 4973-4978
[PubMed] [Full Text HTML] [Full Text PDF]
[78] Murthy M.R., Reid T.J.3rd, Sicignano A., Tanaka N., Musick W.D., Rossmann M.G.: Structure of beef liver catalase. J. Mol. Biol., 1981; 152: 465-499
[PubMed]
[79] Nicholls P., Fita I., Loewen P.C.: Enzymology and structure of catalases. Adv. Inorg. Chem., 2001; 51: 51-106
[80] Niikawa N., Fukushima Y., Taniguchi N., Iizuka S., Kajii T.: Chromosome abnormalities involving 11p13 and low erythrocyte catalase activity. Hum. Genet., 1982; 60: 373-375
[PubMed]
[81] Ogata M.: Acatalasemia. Hum. Genet., 1991; 86: 331-340
[PubMed]
[82] Omar R.A., Chyan Y.J., Andorn A.C., Poeggeler B., Robakis N.K., Pappolla M.A.: Increased expression but reduced activity of antioxidant enzymes in Alzheimer’s disease. J. Alzheimers Disease., 1999; 1: 139-145
[PubMed]
[83] Pastor M.C., Sierra C., Dolade M., Navarro E., Brandi N., Cabre E., Mira A., Seres A.: Antioxidant enzymes and fatty acid status in erythrocytes of Down’s syndrome patients. Clin. Chem., 1998; 44: 924-929
[PubMed] [Full Text HTML] [Full Text PDF]
[84] Percy M.E.: Catalase: an old enzyme with a new role? Can. J. Biochem. Cell Biol., 1984; 62: 1006-1014
[PubMed]
[85] Proudfoot N.J., Brownlee G.G.: Nucleotide sequences of globin messenger RNA. Br. Med. Bull., 1976; 32: 251-256
[PubMed]
[86] Putnam C.D., Arvai A.S., Bourne Y., Tainer J.A.: Active and inhibited human catalase structures: ligand and NADPH binding and catalytic mechanism. J. Mol. Biol., 2000; 296: 295-309
[PubMed]
[87] Quan F., Korneluk R.G., MacLeod H.L., Tsui L.C., Gravel R.A.: An RFLP associated with the human catalase gene. Nucleic Acids Res., 1985, 13: 8288
[PubMed] [Full Text HTML] [Full Text PDF]
[88] Quan F., Korneluk R.G., Tropak M.B., Gravel R.A.: Isolation and characterization of the human catalase gene. Nucleic Acid Res., 1986; 14: 5321-5335
[PubMed] [Full Text HTML] [Full Text PDF]
[89] Radi R., Beckman J.S., Bush K.M., Freeman B.A.: Peroxynitrite oxidation of sulfhydryls. The cytotoxic potential of superoxide and nitric oxide. J. Biol. Chem., 1991; 266: 4244-4250
[PubMed] [Full Text PDF]
[90] Reid T.J.3rd, Murthy M.R., Sicignano A., Tanaka N., Musick W.D., Rossmann M.G.: Structure and heme environment of beef liver catalase at 2.5 A resolution. Proc. Natl. Acad. Sci. USA, 1981; 78: 4767-4771
[PubMed] [Full Text HTML] [Full Text PDF]
[91] Rhee S.G., Yang K.S., Kang S.W., Woo H.A., Chang T.S.: Controlled elimination of intracellular H(2)O(2): regulation of peroxiredoxin, catalase, and glutathione peroxidase via post-translational modification. Antioxid. Redox Signal., 2005; 7: 619-626
[PubMed]
[92] Roels F., Geerts A.: Cytoplasmic catalase: cytochemistry and. physiology. Ann. NY Acad. Sci., 1982; 386: 534-536
[93] Safo M.K., Musayev F.N., Wu S.H., Abraham D.J., Ko T.P.: Structure of tetragonal crystals of human erythrocyte catalase Acta Crystallogr. D. Biol. Crystallogr., 2001; 57: 1-7
[PubMed]
[94] Sasnoor L.M., Kale V.P., Limaye L.S.: Prevention of apoptosis as a possible mechanism behind improved cryoprotection of hematopoietic cells by catalase and trehalose. Transplantation, 2005; 80: 1251-1260
[PubMed]
[95] Scandalios J.G.: Molecular Biology of Free Radical Scavenging Systems. New York, 1992; Cold Spring Harbor Labolatory Press, NY
[96] Scandalios J.G., Guan L., Polidoros A.N.: Catalase in plants: gene structure, properties, regulation, and expression. Cold Spring Harbor Laboratory Press, 1997, Cold Spring Harbor, NY
[97] Schonbaum G.R., Chance B.: Catalase. In: The. Enzymes, Vol. 13, 3rd edn. (Boyer PD, ed). New. York: Academic Press, 1976; 363-408
[98] Schroeder W.T., Saunders G. F.: Localization of the human catalase and apolipoprotein A-I genes to chromosome 11. Cytogenet. Cell Genet., 1987; 44: 231-233
[PubMed]
[99] Schubert J., Wilmer J.W.: Does hydrogen peroxide exist “free’ in biological systems? Free Radic. Biol. Med., 1991; 11: 545-555
[PubMed]
[100] Sies H.: Biochemistry of the peroxisome in the liver cell. Angew. Chem. Int. Ed. Engl. 1974, 13: 706-718
[PubMed]
[101] Sies H.: Oxidative stress: oxidants and antioxidants. Exp. Physiol., 1997; 82: 291-295
[PubMed] [Full Text PDF]
[102] Singh I.: Mammalian peroxisomes: metabolism of oxygen and reactive oxygen species. Ann. N. Y. Acad. Sci., 1996; 804: 612-627
[PubMed]
[103] Speranza M., Bagley A.C., Lynch R.E.: Cells enriched for catalase are sensitized to the toxicities of bleomycin, adriamycin, and paraquat. J. Biol. Chem. 1993; 268: 19039-19043
[PubMed] [Full Text PDF]
[104] Storz G., Imlay J.A.: Oxidative stress. Curr. Opin. Microbiol., 1999; 2:188-194
[PubMed]
[105] Sumner J.B., Dounce A.L.: Crystalline catalase. J. Biol. Chem., 1937; 121: 417-424
[106] Sun W., Kadima T.A., Pickard M.A., Dunford H.B.: Catalase activity of chloroperoxidase and its interaction with peroxidase activity. Biochem. Cell. Biol., 1994; 72: 321-331
[PubMed]
[107] Sun Y.: Free radicals, antioxidant enzymes, and carcinogenesis. Free Radic. Biol. Med., 1990; 8: 583-599
[PubMed]
[108] Switala J., Loewen P.C.: Diversity of properties among catalases. Arch. Biochem. Biophys., 2002; 401: 145-154
[PubMed]
[109] Takahara S.: Progressive oral gangrene, probably due to lack of catalase in blood (acatalasema): report of nine cases. Lancet., 1952; 11: 1101-1104
[PubMed]
[110] Ueda M., Kinoshita H., Maeda S.I., Zou W., Tanaka A.: Structure-function study of the amino-terminal stretch of the catalase subunit molecule in oligomerization, heme binding, and activity expression. Appl. Microbiol. Biotechnol., 2003; 61: 488-494
[PubMed]
[111] Vainshtein B.K., Melik-Adamyan W.R., Barynin V.V., Vagin A.A., Grebenko A.I.: Three-dimensional structure of the enzyme catalase. Nature., 1981; 293: 411-412
[PubMed]
[112] Vitai M., Goth L.: Reference ranges of normal blood catalase activity and levels in familial hypocatalasemia in Hungary. Clin. Chim. Acta., 1997; 261: 35-42
[PubMed]
[113] von Ossowski I., Hausner G., Loewen P.C.: Molecular evolutionary analysis based on the amino acid sequence of catalase. J. Mol. Evol., 1993; 37: 71-76
[PubMed]
[114] Welinder K.G.: Superfamily of plant, fungal and bacterial peroxidases. Curr. Opin. Struct. Biol., 1992; 2: 388-393
[115] Wen J.K., Osumi T., Hashimoto T., Ogata M.: Molecular analysis of human acatalasemia: identification of a splicing mutation. J. Molec. Biol., 1990; 211: 383-393
[PubMed]
[116] Wieacker P., Mueller C.R., Mayerova A., Grzeschik K.H., Ropers H.H.: Assignment of the gene coding for human catalase to the short arm of chromosome 11. Ann. Genet., 1980, 23: 73-77
[PubMed]
[117] Yabuki M., Kariya S., Ishisaka R., Yasuda T., Yoshioka T., Horton A.A., Utsumi K.: Resistance to nitric oxide-mediated apoptosis in HL-60 variant cells is associated with increased activities of Cu,Zn-superoxide dismutase and catalase. Free Radic. Biol. Med., 1999; 26: 325-332
[PubMed]
[118] Yamamoto K., Volkl A., Hashimoto T., Fahimi H.D.: Catalase in guinea-pig hepatocytes in localised in cytoplasm, nuclear matrix and peroxisomes. Eur. J. Cell Biol.,1988; 46: 129-135
[PubMed]
[119] Yasmineh W.G., Kaur T.P., Blazar B.R., Theologides A.: Serum catalase as marker of graft-vs-host disease in allogeneic bone marrow transplant recipients: pilot study. Clin. Chem., 1995; 41: 1574-1580
[PubMed]
[120] Zamocky M., Koller F.: Understanding the structure and function of catalases: clues from molecular evolution and in vitro mutagenesis. Prog. Biophys. Mol. Biol., 1999; 72: 19-66
[PubMed]