
High doses of tocopherol in the prevention and potentiation of dioxin in experimental inflammation – potential application
Ireneusz Całkosiński 1 , Joanna Rosińczuk-Tonderys 2 , Monika Szopa 3 , Maciej Dobrzyński 4 , Andrzej Gamian 5Streszczenie
Emisja do środowiska toksycznych i stabilnych substancji chloroorganicznych, z których jedną z ważniejszych grup są dioksyny, stanowi istotny problem ekologiczny i zdrowotny. Polichlorowane dibenzodioksyny (PCDD), określane potocznie jako dioksyny, stanowią grupę 75 związków aromatycznych z przyłączonymi w różnych pozycjach i ilościach atomami chloru. Podobną budową i właściwościami charakteryzują się polichlorowane dibenzofurany (PCDF) oraz polichlorowane bifenyle (PCB).
W ostatnich czterech latach daje się zauważyć znaczące zainteresowanie biologicznym działaniem dioksyn i ich obecnością w łańcuchach pokarmowych. Wykazano, że jedną z ważniejszych dróg ich oddziaływania na organizm jest pobudzanie kaskady cytokin zapalnych i wywoływanie stresu oksydacyjnego. Skutecznym sposobem przeciwdziałania zatruciom może być podawanie antyutleniaczy oraz czynników przeciwzapalnych. Działanie ochronne wspomnianych substancji, w tym wysokich dawek tokoferolu, potwierdza wiele badań nad zwierzętami doświadczalnymi.
Słowa kluczowe:dioksyny • TCDD • tokoferol • odczyn zapalny
Summary
Emission of toxic and persistent organochlorine compounds into the environment is known to have been recently a very important environmental and health problem. One of the major groups of these compounds is the polychlorinated dibenzodioxins (PCDDs), commonly named dioxins, including 75 compounds composed of benzene rings with bound chlorine atoms in different positions and quantities. Polychlorinated dibenzofurans (PCDFs) and polychlorinated biphenyls (PCBs) are known to have a similar structure and properties.
The biological effects of dioxins in food chains have been widely studied in the literature on the subject for the last four years. The major pathways of their impact on the organism are suspected to stimulate the cascade of inflammatory cytokines and the induction of oxidative stress. An effective way to prevent poisoning may, therefore, be expected to be administration of antioxidants and anti-inflammatory agents. Recent studies in experimental animals have confirmed the protective action of high doses of tocopherol and mentioned substances against dioxins.
Key words:dioxin • TCDD • tocopherol • inflammation
1. Toksyczne działanie dioksyn
Do trwałych zanieczyszczeń organicznych (TZO), (persistent organic ollutants – POPs), zalicza się grupę związków określanych jako dioksyny. Powstają one jako produkty uboczne wielu procesów technologicznych i spalania związków organicznych. Charakteryzują się m.in. takimi cechami jak: zdolność wywoływania efektów toksycznych, trwałość we wszystkich elementach środowiska (tzw. persystencja), zdolność do biokumulacji i do transportu atmosferycznego na duże odległości, zdolność do wywoływania niekorzystnych skutków w środowisku i u człowieka [137].
Narastająca emisja dioksyn do środowiska stanowi obecnie istotny problem ekologiczny i zdrowotny [84,116]. Mimo że badania nad działaniem dioksyn na organizm są prowadzone od ponad dwudziestu lat, a obecnie emisja tych związków do środowiska jest ograniczana przez restrykcyjne przepisy prawne, ujęte w Raportach Konwencji Sztokholmskiej, jak i ustawodawstwie krajowym, to znajomość społeczna tej problematyki ekologiczno-zdrowotnej jest niewielka. W ostatnich trzech latach ponownie wzrosło zainteresowanie oddziaływaniem biologicznym tych związków z powodu pojawienia się nowych źródeł ich emisji [21].
W grupie chloroorganicznych związków zaliczanych do TZO wyróżnia się polichlorowane dibenzodioksyny PCDD, polichlorowane dibenzofurany PCDF i polichlorowane bifenyle PCB (ryc. 1). Polichlorowane dibenzodioksyny PCDD określane potocznie jako dioksyny stanowią grupę 75 związków zbudowanych z pierścieni benzenowych, do których przyłączone są w różnych pozycjach i ilości (Clx,Cly) atomy chloru. Wymienione związki mogą tworzyć kongenery różniące się liczbą i usytuowaniem atomów chloru w szkielecie benzenowym. W powyższej grupie wzorcowym związkiem jest 2,3,7,8-tetrachlorodibenzo-p-dioksyna (2,3,7,8 TCDD).

Ryc. 1. Wzory strukturalne przedstawicieli poszczególnych grup TZO; a) – polichlorobifenyle (PCB), b) – polichlorodibenzofurany (PCDF), c) – polichlorodibenzodioksyny (PCDD), d) – 2,3,7,8-tetrachlorodibenzo-p-dioksyna (TCDD)
Dioksyny jako grupa związków od dnia 1.06.1997 r. została uznana przez Międzynarodową Agencję Badań nad Nowotworami (IARC) za karcynogeny grupy A, które to działanie wykazano na zwierzętach doświadczalnych [53,131].
Dioksyna 2,3,7,8-tetrachlorodibenzo-p-dioksyna (TCDD) jest najlepiej zbadanym związkiem mającym dużą toksyczność i stanowiącym wzorzec odniesienia toksyczności dla innych związków tej grupy [1,72]. Toksyczność TCDD jest określana jako równoważny współczynnik toksyczności (toxic equivalent factor – TEF) i jest on równy jedności (tab. 1) [56].
Tabela 1. Toksyczność dioksyn określana równoważnikiem współczynnika toksyczności

Dioksyny powstają w każdym procesie termicznym, jeśli w środowisku spalania znajduje się materiał organiczny oraz chlor i sprzyjające czynniki, tj. temperatura spalania 200-900°C, mały dopływ tlenu i obecność wody. Istnieje wiele źródeł emisji dioksyn do środowiska m.in. związanych z różnorodnymi procesami spalania, bądź też wytwarzania ich w wielu procesach technologicznych, a także tworzenia się w wyniku oddziaływania promieni świetlnych na składowane odpady z tworzyw sztucznych. Istotne jest również zjawisko powstawania dioksyn z PCB (polichlorowanych bifenyli), związki te zalicza się do tej samej grupy ze względu na duże podobieństwo chemiczne [81].
Dioksyny również mogą być tworzone w wielu procesach technologicznych, w których jest używany chlor. Dotyczy to przemysłu tekstylnego, papierniczego czy produkującego środki ochrony roślin. Bezpośrednim, znaczącym źródłem tych związków są procesy spalania różnego rodzaju substancji organicznych, w tym tworzyw sztucznych, co jest związane z technologiami utylizacji odpadów. Ostatnie doniesienia literaturowe wykazują, że znaczącym źródłem emisji PCB i kongenerów PCCD były powstałe w wyniku różnych przyczyn pożary, np. atak terrorystyczny na World Trade Center (WTC). Przeprowadzone pomiary w rejonie pożaru wykazały duży wzrost stężenia policyklicznych aromatycznych węglowodorów nie tylko w strefie zero, ale i w znacznej od niej odległości [46,104,105,120,121]. Poczynione obserwacje mogą wskazywać na istnienie podwyższonego ryzyka zachorowania na chorobę nowotworową ludności zamieszkującej w bliskiej odległości od WTC [105]. Stwierdzono również znaczny wzrost emisji dioksyn związany z utylizacją okrętów w krajach rozwijających się (Indie, Bangladesz).
Badania przeprowadzone na terenach objętych trzydzieści lat wcześniej działaniami wojennymi, w których zastosowano herbicydy zawierające w swoim składzie dioksyny, wykazały wielokrotne podwyższenie stężenia PCDF w glebie. Obserwacje te mają istotne znaczenie do wykazania powiązań między produkcją żywności na terenach skażonych policyklicznymi związkami aromatycznymi a włączaniem tych związków do łańcucha pokarmowego [82].
Liczne doniesienia literaturowe wskazują, że dioksyny, zaliczane do ksenobiotyków wykazujących hormonopodobne działanie, występują w środowisku wielkomiejskim w znacznym stężeniu, co jest związane z różnego rodzaju procesami utylizacji i indywidualnej produkcji ciepła [53,54,100,101,116].
Ograniczanie emisji trwałych związków organicznych (TZO), do których są zaliczane PCDD/PCDF występujące w różnych proporcjach, co uwarunkowuje ich stopień toksyczności, jest regulowane przez przepisy prawne Konwencji Sztokholmskiej. W dyrektywach tego raportu do głównych źródeł emisji środowiskowej TZO zaliczane są otwarte pożary budynków, lasów oraz wysypisk śmieci. Do identyfikacji emisji TZO i określania w nich stosunku PCDD/PCDF jest używana metoda Toleit [43]. Grupa wymienionych wyżej związków chloroorganicznych, występująca w środowisku jedynie w śladowych stężeniach, ma jednak znaczące konsekwencje biologiczne, co jest związane ze szczególną ich zdolnością do długotrwałego kumulowania w organizmie. Prowadzi to do naruszania różnych mechanizmów homeostatycznych, które są odpowiedzialne za adaptację organizmu do środowiska. Długotrwałe, silne działanie biologiczne dioksyn może się przejawiać nawet w drugim pokoleniu, które nie było bezpośrednio narażone na działanie tych związków [14,17,18,19,21,130].
Znaczącymi drogami wnikania dioksyn do organizmu są: droga pokarmowa, aerogenna oraz przez skórę.
Istotnym elementem sprzyjającym wnikaniu tych związków do organizmu jest ich powinowactwo do tłuszczy, w których się rozpuszczają, mogą być transportowane i magazynowane w organizmie [72,114]. Większość dioksyn (około 90%) wnikająca do organizmu drogą pokarmową [42] pochodzi z różnorodnych łańcuchów pokarmowych (ryc. 2). Potwierdzają to badania przeprowadzone na grupie wolontariuszy, które wykazały, że prawie 87% podanej dawki dioksyn wchłania się drogą pokarmową [138]. Przez to związki te występują w bazie pokarmowej niepodlegającej bezpośrednim wpływom skażeń emitowanych przez przemysł, czego przykładem jest, proporcjonalnie do wieku, duża zawartość skumulowanych dioksyn w organizmie fok spożywanych m.in. przez Eskimosów i niedźwiedzie polarne żyjące w Kanadzie i na Grenlandii [94,95,114]. Powyższe badania dowodzą, że ilość zdeponowanych dioksyn w tkance tłuszczowej konsumentów ostatecznych jest wprost proporcjonalna do długości ich życia w środowisku skażonym dioksynami i odkładania tkanki tłuszczowej w wyniku spożywania pokarmów bogatych w tłuszcze zawierające dioksyny [52,114,130]. Badania fok żyjących u wybrzeży Grenlandii, będących źródłem pożywienia niedźwiedzi polarnych i ludności eskimoskiej wykazały znaczne stężenia dioksyn, przy czym stwierdzono korelację między wiekiem tych ssaków a stężeniem tych ksenobiotyków w ich ciele [95]. Również u norweskich niedźwiedzi polarnych wykazano znaczne stężenie PCDD/PCDF w ich mleku porównywalne ze stężeniem tych związków w mleku fok żyjących na tym samym obszarze [94]. Stwierdzono, że w mniejszym stopniu wnikanie dioksyn do organizmu odbywa się drogą aerogenną (8%) (powietrze zanieczyszczone dymami i pyłami, na których te związki są absorbowane) oraz przez skórę (2% – za pośrednictwem lipidów skóry). Wartości te są szacunkowe, ponieważ różne czynniki mogą w znaczący sposób na nie wpływać, np. sadze, które wnikają przez skórę i drogi oddechowe [54]. Znajomość źródeł emisji dioksyn przyczynia się do wyodrębnienia grup zawodowych o szczególnym narażeniu na działanie PCDD/PCDF, do których zalicza się: kominiarzy, u których stwierdzono obecność polichlorowanych aromatycznych związków w moczu [12,137], strażaków, żołnierzy biorących udział w operacjach wojennych i antyterrorystycznych. Pracownicy zatrudnieni w pewnych gałęziach przemysłu oraz osoby zamieszkujące w pobliżu tych zakładów są narażeni na działanie dioksyn. Spektakularnym tego przykładem jest zatrucie pracowników i okolicznej ludności w wyniku katastrofy w Seveso (Włochy) [5,88].

Ryc. 2. Schemat wnikania i przemian dioksyn w organizmie
Istnieje ponadto niebezpieczeństwo wykorzystywania dioksyn w aktach terrorystycznych [117], ze względu na ich właściwości organoleptyczne związane z niewykrywalnością w pokarmie z powodu braku smaku i zapachu oraz dobrej rozpuszczalności w tłuszczach. Znaczącą właściwością tych substancji jest pojawienie się objawów klinicznych zatrucia po dłuższym okresie od ich spożycia czego przykładem jest otrucie byłego prezydenta Ukrainy Wiktora Juszczenki [115].
Przeprowadzone badania u ludzi po 20 i 30 latach od chwili zatrucia wykazały, że stężenie dioksyn w lipidach surowicy kształtowało się w podobnym przedziale wartości, który zaobserwowano w chwili skażenia. Stwierdzono ponadto, że stężenie dioksyn korelowało ze wskaźnikiem BMI (Body Mas Index), którego istotny wzrost częściej występował u kobiet. Tłumaczy to się uwalnianiem po pewnym czasie od zatrucia zdeponowanych w tkance tłuszczowej dioksyn do krwi wskutek przewagi procesów katabolicznych związanych z wiekiem [5,12,88].
Doniesienia literaturowe dotyczące toksycznego działania dioksyn na poszczególne gatunki zwierząt nie są zbyt liczne. Wykazano, że najbardziej wrażliwym gatunkiem na działanie TCDD określane przez DL50 w µg/kg m.c. jest świnka morska [109]. Szczury są natomiast 45 razy bardziej oporne na działanie TCDD w rzędzie wielkości zbliżonym do małp. Badania przeprowadzone na samicach szczurach poddanych działaniu TCDD wykazały u tych zwierząt spadek płodności, a urodzone przez nie potomstwo cechowało się skróconą przeżywalnością [16]. Tę specyficzną gatunkowo wrażliwość na dioksyny tłumaczy się zróżnicowaną ekspresją genów odpowiedzialnych za metabolizm lipidów [8].
Wykazano, że wrażliwość na działanie dioksyn jest również zależna od płci. Większą wrażliwość stwierdzono u samic szczurów, u których toksyczność TCDD określona przez DL50 wynosiła 22-50 µg/kg m.c., natomiast u samców dawka ta kształtowała w przedziale 45-50 µg/kg m.c. [69]. W badaniach własnych stwierdzono, że dawka DL50 TCDD wynosząca 20 µg/kg m.c. rozpuszczona w DMSO podana podskórnie 8 samicom szczepu Bufallo spowodowała w ciągu 72 godzin śmierć wszystkich osobników.
2. Mechanizm toksyczności
Znajomość mechanizmu działania dioksyn na metabolizm komórki tłumaczy powstawanie wielu zaburzeń czynności organizmu poddanego ekspozycji na te związki (ryc. 3).
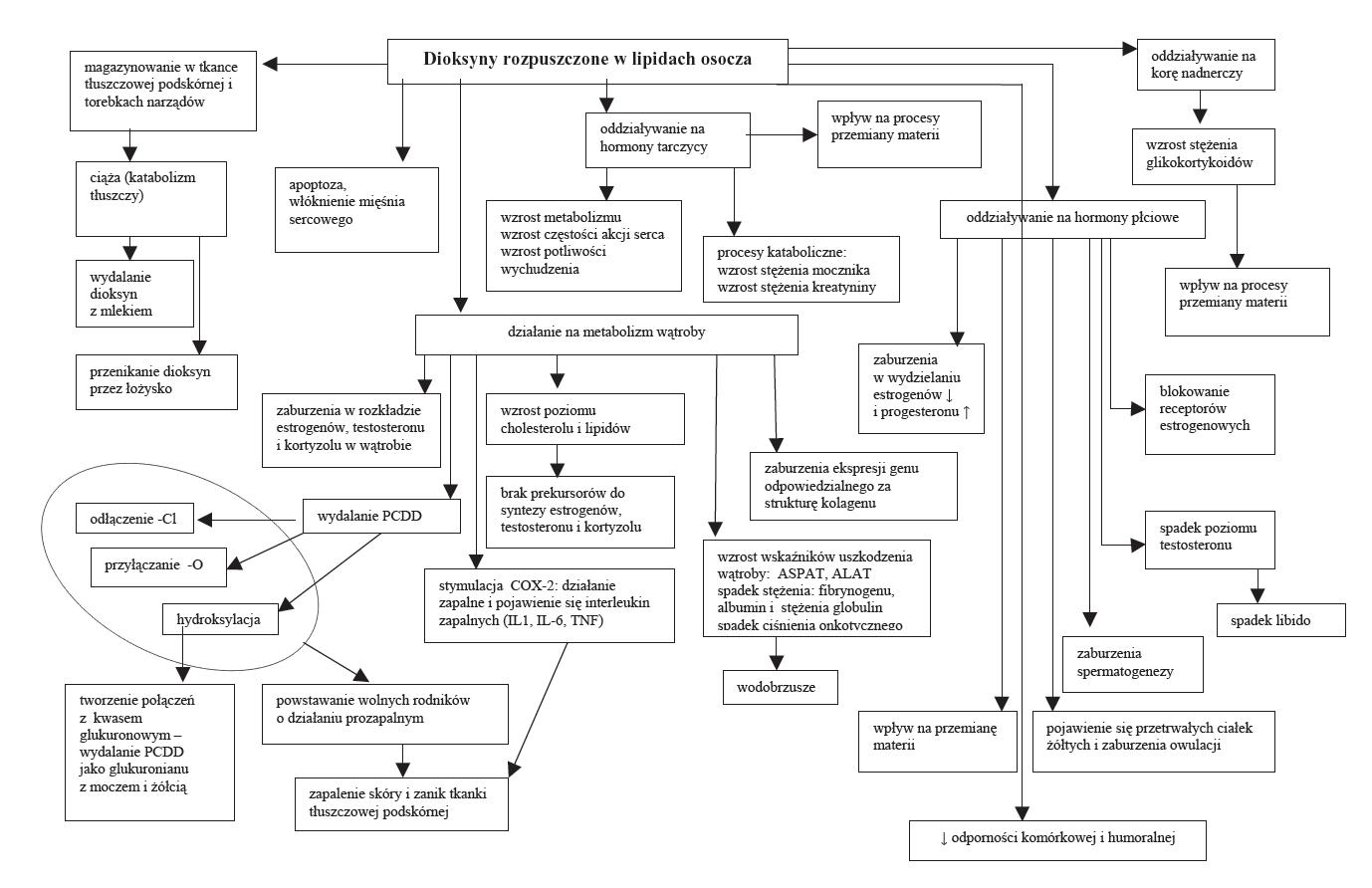
Ryc. 3. Schemat oddziaływania dioksyn na funkcje narządów
Dioksyny ze środowiska zewnętrznego transportowane są przez lipidy osocza krwi do wątroby i do tkanki tłuszczowej, gdzie dochodzi do ich kumulacji. TCDD łączy się w cytosolu hepatocytu z receptorem węglowodorów arylowych – AhR (aryl hydrocarbon receptor) [99,102].
Receptor ten wykazuje duże podobieństwo do receptorów hormonów steroidowych AhE. Uaktywniony kompleks dioksyna-receptor (ligand-receptor) przechodzi do jądra, gdzie łączy się z fragmentem DNA (XRE – dioxin response element), wykazującym powinowactwo do tych związków (ryc. 4).
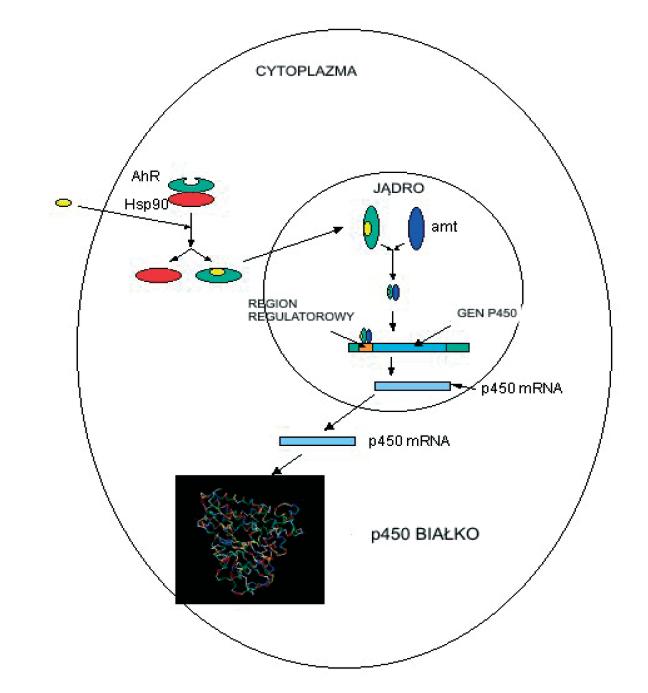
Ryc. 4. Receptorowy mechanizm działania dioksyn
W wyniku tego połączenia dochodzi do stymulacji transkrypcji genu cytochromu P-450 i w efekcie do wzmożonego wytwarzania cytochromu P-450, a także cytochromu P-448 (hydroksylazy węglowodorów aromatycznych – AHH). Największe stężenie hydroksylazy AHH występuje w siateczce śródplazmatycznej hepatocytu. Enzym ten bierze udział w przemianach dioksyn, które przebiegają w dwóch etapach: pierwszy to hydroksylacja dioksyn, katalizowana przez monooksygenazy (cytochromy P-450), drugi to sprzęganie hydroksylowanych dioksyn z kwasem glukuronowym lub glutationem bądź ich acetylowanie. Powstałe związki stają się rozpuszczalne w wodzie i są wydalane z organizmu z żółcią lub moczem.
Metabolizm dioksyn w organizmie jest utrudniony ze względu na obecność w ich cząsteczkach pierścieni aromatycznych, charakteryzujących się stosunkowo dużą stabilnością i trwałością chemiczną. Stabilność pierścienia dodatkowo wzmacnia przyłączenie atomów chloru. Pierwszym etapem przemian dioksyn jest hydroksylacja ich pierścieni aromatycznych polegająca na podstawianiu grup hydroksylowych -OH w miejsce atomu wodoru lub chloru. Jednym z produktów hydroksylacji jest cis-1,2-dihydro-1,2-dihydroksybenzen, który następnie jest przekształcany przez dehydrogenazy do katecholu lub hydrochinonu (ryc. 5).

Ryc. 5. Mechanizm rozkładu dioksyny w organizmie – proces hydroksylacji
Reakcja ta może być poprzedzona epoksydacją, w następstwie której do wiązania podwójnego pierścienia przyłącza się atom tlenu, ostatecznie zastąpiony przez grupę -OH pod wpływem hydroksylazy epoksydowej (ryc. 6).

Ryc. 6. Mechanizm rozkładu dioksyny – proces epoksydacji
Hydroksylacja powoduje zwiększenie hydrofilowości dioksyn, a tym samym umożliwia dalsze etapy ich rozkładu, takie jak rozszczepienie pierścienia oraz sprzęganie z kwasem glukuronowym katalizowane przez odpowiednie transferazy [60].
Powyższe procesy związane z różnokierunkowym rozpadem cząsteczki dioksyny są przyczyną powstawania wolnych rodników. W wyniku przedstawionych wyżej przemian cząsteczek dioksyn w organizmie następuje generacja wolnych rodników powodujących stres oksydacyjny, który ma właściwości prozapalne i przyczynia się do nasilenia istniejących odczynów zapalnych. Powyższe prozapalne właściwości dioksyn wykazano w badaniach własnych na podstawie zachowania się stężenia interleukin prozapalnych, zwłaszcza u zwierząt poddanych działaniu TCDD dochodzi do intensywnego wzrostu TNF [14].
Ostanie doniesienia wykazują, że w eliminacji dioksyn mogą mieć znaczenie niektóre cytokiny. Stwierdzono, że aktywowanie receptora AhR przez IL-4 spowodowało ekspresję genu CYP1A1, odpowiedzialnego za metabolizm ksenobiotyków, takich jak TCDD [119].
Stopniowe i powolne kumulowanie się dioksyn w organizmie (komórki wątroby, tkanka tłuszczowa) oraz wiązanie się z DNA, ma istotny wpływ na przemiany białkowe w hepatocytach i układzie krwiotwórczym. Może to być przyczyną obserwowanych zakłóceń w syntezie białek osoczowych i zmian w obrazie krwi obwodowej [62,99]. Dowodem na to jest stwierdzenie, że w wątrobie 25-75% podanej dawki dioksyn jest deponowane w ciągu jednego dnia. Badania Fowlera i wsp. [45] wykazały, że po podaniu 5-25 µg/kg m.c. TCDD, wystąpiły zmiany w siateczce gładkiej endoplazmatycznej hepatocytów między 6 a 9 dniem od podania dioksyny. Takie dawki tego związku powodowały zmniejszenie wydzielania żółci oraz zwiększenie stężenia koproporfiryn [61]. Podanie dawki 100 µg/kg m.c. TCCD myszom powodowało wzrost masy ich wątroby w 72 godzinie od czasu podania, któremu nie towarzyszyły wyraźne zmiany histopatologiczne w tym narządzie. Dochodziło ponadto do wzrostu aktywności aminotransferazy alaninowej w surowicy [139]. W badaniach autora stwierdzono u szczurów po 3 tygodniach od podania 5 µg/kg m.c. TCCD zmiany makroskopowe i histopatologiczne w hepatocytach, przejawiające się ich stłuszczeniem, a także znacznie podwyższonym stężeniem cholesterolu w surowicy. W badaniach innych autorów przeprowadzonych na małpach (Callithrix jacchus) i myszach, którym podano TCDD obserwowano m.in. wzrost stężenia laminy A i podwyższenie aktywności transferyny AlAT w wątrobie [93,139].
Obecnie prowadzone prace doświadczalne w ośrodkach zagranicznych dotyczą wpływu różnych dawek dioksyn na OUN oraz mięsień sercowy zwierząt doświadczalnych. Badania te wykazują prozapalny charakter dioksyn związany z wywoływaniem stresu oksydacyjnego i ich kardiotoksyczne oddziaływania, objawiające się rozrostem tkanki łącznej [91,93,106].
Wykazano, że dioksyny mogą zaburzać wydzielanie TSH i hormonów tarczycy (T3, T4), powodując wzrost ich stężenia we krwi. Natomiast poprzez oddziaływanie na receptor AhE powodują zakłócenia w regulacji wydzielania hormonów płciowych [28,29,51,52], co przejawia się zaburzeniami cyklu estrogenowego [16,18,23,24,80]. Badania Sonne’a i wsp., [114] oraz Verreault i wsp. [130] również wykazały negatywne oddziaływanie dioksyn kumulowanych w organizmie niedźwiedzi polarnych na procesy rozrodcze. U zwierząt tych sekcyjnie stwierdzono zmniejszenie masy narządów płciowych oraz gonad u obu płci, zaburzenia spermato- i oogenezy; obserwowano ponadto występowanie osobników hemafrodytycznych. Występowały również poronienia oraz duża śmiertelność okołoporodowa. Stwierdzono, że dioksyny mogą być wydalane w czasie laktacji w postaci aktywnej w tłuszczu mleka [94,99]. Potwierdza to zmniejszona przeżywalność młodych niedźwiedzi, karmionych przez niedźwiedzice, których mleko zawierało dużą ilość dioksyn. Wynikiem tego były obserwowane zaburzenia odporności [114]. Powyżej opisane anomalie w układzie rozrodczym i zaburzenia behawioralne u osobników młodocianych pokrywają się z wcześniejszymi obserwacjami autora uzyskanymi w doświadczeniach prowadzonych na szczurach, u których również stwierdzono zmniejszenie masy ciała oraz zaburzenia dojrzewania osobników młodocianych pochodzących od matek poddanych działaniu dioksyn [16]. Przyczyną opisanych zmian w regulacji hormonalnej jest supresyjne działanie dioksyn na transkrypcję genów kodujących receptory estrogenowe [122].
Największe niebezpieczeństwo niosą niewielkie, często pomijane, stężenia dioksyn w środowisku. Uszkadzają one układ immunologiczny i powodują zaburzenia w reprodukcji.
Badania wykazały, że najbardziej wrażliwy na działanie dioksyn jest układ immunologiczny. Wpływają one na wiele procesów odpornościowych z czym wiąże się występowanie zespołu chlorakne, przebiegającego w postaci przewlekłych alergii skórnych [12,34,88,115,117]. Dioksyny powodują inwolucję grasicy [62], wzrost stężenia kortykosteroidów [86] oraz zmiany w składzie białek osocza, przejawiające się zwiększeniem stężenia α- i β-globulin [133], jak i opóźnioną reakcją immunologiczną [133].
Dioksyny oddziaływając na receptor komórkowy AhR przyczyniają się do pobudzenia wytwarzania niektórych cytokin wpływających na rozwój komórek krwi. TCDD, będąca agonistą tego receptora hamuje ekspresję mRNA IL-6, pod wpływem podanego lipopolisacharydu (LPS). Wynikiem tego jest oddziaływanie na komórki hematopoetyczne i limfocyty B. Przyczynia się to również do zmniejszenia syntezy globulin [63]. Również badania Rodriguez-Sosa i wsp. wykazały, że TCDD dodana do hodowli limfocytów B stymulowanych przez LPS i IL-4 powoduje zahamowanie wydzielania kilku klas immunoglobulin, tj. IgG1, IgE i IgM. Wykazano również, że TCDD nie przyczynia się do wzrostu syntezy IgE przez transformowane limfocyty B. TZO powodują długofalową immunosupresję prelimfocytów B w szpiku kostnym, związaną z pobudzeniem procesów apoptozy. Wyniki badań wskazują, że aktywacja AhR oddziałuje na hematopoezę niedojrzałych limfocytów w wyniku działania małych dawek wielu związków zaliczanych do TZO [87]. Opisane efekty immunosupresyjne znajdują swoje odzwierciedlenie w zmniejszonej przeżywalności osobników młodocianych [65]. W doświadczeniach na małpach (Callithrix jacchus) wykazano zmniejszenie bezwzględnej i względnej liczby limfocytów w odniesieniu do ogólnej liczby leukocytów w 3 tygodniu od podania TCDD w ilości 300 ng/kg m.c. Zaobserwowano również spadek o 20% populacji limfocytów CD4. Zmiany te są tłumaczone oddziaływaniem dioksyn na grasicę [89,90]. Badania grasicy małp poddanych działaniu różnych dawek TCDD wykazały wzrost TGF-β i integryn zależny od dawki dioksyn [91].
Badania na myszach immunizowanych SRBC, którym podano TCDD w dawce 5 µg/kg m.c. wykazały spadek liczby limfocytów CD4 i CD8 w porównaniu do grupy kontrolnej, w której liczba ta wzrosła [68]. Ekspozycja na TCDD powoduje zmiany w odporności wrodzonej i nabytej zarówno w komórkowej, jak i humoralnej. Zachowanie się systemu immunologicznego po działaniu dioksyn jest swoistym wskaźnikiem odzwierciedlającym możliwość przystosowania organizmu [59].
Macierz zewnątrzkomórkowa odgrywa istotną rolę w odczynie zapalnym, gdzie mogą przeważać procesy naprawcze, związane z rozrostem tkanki łącznej zastępującej elementy uszkodzone, bądź też mogą przeważać procesy destrukcyjne, przyczyniające się do rozszerzania procesu zapalnego. Ostatnie badania wykazują, że dioksyny mogą oddziaływać na tę strukturę. Badania na myszach wykazały znaczący udział stresu oksydacyjnego na ekspresję wątrobowego genu odpowiedzialnego za syntezę kolagenu [30]. W badaniach własnych stwierdzono, że dioksyny znacząco przyczyniają się do obniżenia stężenia prokolagenu typu 1 [15].
Dioksyny uważa się za substancje przyczyniające się do wywoływania stresu oksydacyjnego i tym samym właściwości prozapalnych, co może mieć znaczenie w zaburzeniach syntezy kolagenu. Wykazano, że TCDD hamuje proces regeneracyjny uszkodzonych płetw ryb (Archocentrus nigrofasciatus), w skład których wchodzi tkanka łączna, w wyniku oddziaływania przez receptor AhR wpływający na tworzenie macierzy [4]. Stwierdzono ponadto hamujący efekt TCDD na syntezę kolagenu typu I, osteopontyny i sialoproteiny kości oraz zredukowanie aktywności fosfatazy alkalicznej w hodowlach tkanki kostnej [111].
Riecke i wsp. [106] oraz Oberemm i wsp. [93] wykazali kardiotoksyczne działanie stosowania różnych dawek TCDD podawanych podskórnie małpom z gatunku marmoseta. Badania te wykazały odmienny charakter działania TCDD w sercu przejawiający się wzrostem ilości kolagenu, fibronektyny i lamininy.
Nieliczne doniesienia [132] wykazują wpływ dioksyn na syntezę prostaglandyn i tromboksanów, pojawiających się w początkowej fazie odczynu zapalnego, mających znaczący udział w następnym etapie tego odczynu, związanym m.in. z syntezą niektórych białek ostrej fazy, jak i zmianami w obrazie krwi [14]. Udowodniono, że TCDD indukuje stres oksydacyjny związany z powstawaniem reaktywnych form tlenu, tj. nadtlenek wodoru w różnych narządach (mózg, jądra) przy jednoczesnym obniżeniu stężenia antyoksydacyjnych enzymów, tj. dysmutaza nadtlenkowa, katalaza, reduktaza i peroksydaza glutationu [74,75].
W związku z tym, że dioksyny powszechnie występują w środowisku w różnych stężeniach, a ich działanie na organizm wykazuje swoiste i długotrwałe skutki, mogą one znacząco zmieniać przebieg podstawowej i wieloetapowej reakcji obronnej organizmu, jaką jest odczyn zapalny, będący również powodem wywoływania stresu oksydacyjnego [14].
3. Stres oksydacyjny w odczynie zapalnym
Odczyn zapalny jest wielofazową reakcją obronną, mającą na celu zneutralizowanie czynnika patogennego oraz wzbudzenie procesów reperacyjnych, przywracających w znacznym stopniu pierwotną homeostazę. Charakter wieloetapowy tego procesu rozłożony w czasie, charakteryzuje się zróżnicowaną dynamiką, która określa jego przebieg jako ostry lub przewlekły [70,140]. W przebiegu reakcji zapalnej wyróżnia się fazę ostrą, pojawiającą się po kilkudziesięciu sekundach od zadziałania czynnika patogennego, trwającą około 12 godz., która następnie przechodzi w fazę zapalenia przewlekłego. Dynamika rozwoju poszczególnych faz doświadczalnego odczynu zapalnego, wraz z jego obrazem ocenionym zarówno w mikroskopie elektronowym, jak i histopatologicznie, zostały szczegółowo przedstawione w monografii autora [14].
Należy podkreślić, że proces zapalny, toczący się w różnych narządach, podlega ogólnym regułom tego zjawiska, jednak wiele wskaźników diagnostycznych odpowiedzi zapalnej wykazuje specyfikę narządową. Również dynamika poszczególnych faz odczynu zapalnego cechuje się różnym natężeniem zmian wskaźników diagnostycznych, tj. hematologicznych, biochemicznych i immunologicznych surowicy [14,18].
Intensywność reakcji organizmu w odczynie zapalnym jest zależna od wielu mechanizmów homeostazy, w których występują sprzężenia dodatnie i ujemne. Uruchomione mechanizmy obrony komórkowej i humoralnej mogą neutralizować czynniki uszkadzające, które są zarazem bodźcami stresowymi.
Sprzężenie ujemne jest realizowane w odczynie zapalnym przez różne mechanizmy ograniczające rozprzestrzenianie się tego procesu, bądź zabezpieczające przed jego uogólnieniem. Przykładem sprzężenia ujemnego jest modyfikacja przewodnictwa neuronalnego w podwzgórzu, gdzie neurony w trakcie odczynu zapalnego toczącego się w narządach, syntetyzują i wydzielają cytokiny prozapalne (TNF, IL-1). Cytokiny te powodują pobudzenie osi podwzgórzowo-przysadkowo-adrenergicznej – HPA axis (hypothalamic-pituitary-arenal axis) do wydzielania przez korę nadnerczy glikokortykosteroidów zwrotnie hamujących wytwarzanie tych cytokin [31,55,127]. Wykazano ponadto, że odczyn zapalny powoduje w podwzgórzu wzrost stężenia CRH (corticotropin-releasing hormone), który to czynnik ma znaczący udział regulatorowy w osi podwzgórzowo-przysadkowo-adrenergicznej. Stwierdzono również, że CRH oprócz funkcji regulatorowej w osi podwzgórzowo-przysadkowo-adrenergicznej działa jako miejscowy pośredni czynnik prozapalny wraz z substancją P. Według badań Crofforda i wsp. [33] CRH oprócz swojej głównej roli w kaskadzie przeciwzapalnej, związanej z wydzieleniem glikokortykosteroidów przez HPA axis, funkcjonuje jako aktywny lokalnie mediator ostrego i przewlekłego zapalenia.
Upośledzenie skuteczności działania wyżej opisanych mechanizmów może być związane z wystąpieniem czynników zakłócających (tab. 2), które przy swojej słabej sile i dynamice działania samoistnie nie są w stanie wywołać odczynu zapalnego [14,15,17,57,73]. Tym niemniej można z dużym prawdopodobieństwem przypuszczać, że długotrwałe ich działanie spowoduje wydłużenie czasu trwania odpowiedzi zapalnej oraz zmianę jej charakteru, polegającą na wolniejszej eliminacji czynnika chorobotwórczego i wytworzeniem słabszej bariery odgraniczającej miejsce zapalenia. Można wyróżnić wiele czynników zakłócających odczyn zapalny (tab. 2).
Tabela 2. Czynniki zakłócające odpowiedź zapalną organizmu

Przedstawione wyżej czynniki dłużej oddziałując na organizm powodują zakłócenia w reakcji zapalnej. Są one w wielu wypadkach wynikiem wzrastającego zanieczyszczenia środowiska i w związku z tym, że trudno je wyeliminować, podejmuje się próby ochrony przed ich działaniem, np. związkami usprawniającymi metabolizm organizmu. Do tych związków zalicza się m.in. witaminy, takie jak tokoferol czy kwas askorbinowy, które mogą zmniejszać stres oksydacyjny powstający w odczynie zapalnym [76,112,134].
Skuteczność działania czynnika wywołującego odczyn zapalny jest zależna od jego siły i czasu działania. Zbyt duża siła czynnika, określana jako bodziec nocyceptywny, oddziaływając na tkankę wywołuje w krótkim czasie destrukcję struktur komórkowych, eliminując tym samym różne etapy reakcji zapalnej w miejscu działania tego bodźca. Znaczący jest również w tej odpowiedzi czas działania czynnika drażniącego, warunkującego przebieg zapalenia (ostre, przewlekle). Reakcja na czynnik uszkadzający struktury tkankowe przejawia się odczynem zapalnym na poziomie lokalnym lub ogólnoustrojowym, wyrażającym się odpowiedzią biochemiczną, hematologiczną i immunologiczną. Zakres i rodzaj procesu zapalnego jest określany przez swoiste cechy czynnika uszkadzającego, a także przez wrażliwość tkankową i narządową. Zazwyczaj reakcja zapalna stanowi lokalną odpowiedź narządową lub układową, która może przejść w zespół ogólnoustrojowej reakcji zapalnej – SIRS (systemic inflammatory response syndrome). Czynniki wywołujące zapalenie mogą być zewnątrz- lub wewnątrzpochodne. Ze względu na rodzaj czynnika indukującego proces zapalny wyróżnia się różne grupy czynników (ryc. 7).

Ryc. 7. Typy odpowiedzi w odczynie zapalnym i możliwości ich monitorowania
Istotną rolę w odczynie zapalnym pełni wątroba, która syntetyzuje wiele białek ostrej fazy. Zaburzenie funkcji tego narządu przez któryś z czynników wymienionych w tabeli 1 może znacząco wywierać wpływ na przebieg odczynu zapalnego zmieniając jego wskaźniki diagnostyczne [6,14,128] (ryc. 7).
Dynamika i natężenie odczynu zapalnego świadczą o możliwości progresji bądź wygasania tego procesu, który można monitorować oznaczając stężenie prozapalnych cytokin, tj. IL-1, IL-6 i TNF w surowicy. Wzrost stężenia cytokin w surowicy następuje w wyniku wydzielania przez wiele komórek leukocytarnych gromadzących się w ognisku zapalnym, co stwierdzono m.in. w badaniach autora w indukowanym zapaleniu opłucnej [14]. Ostatnie doniesienia wskazują na znaczącą rolę makrofagów występujących w jamie opłucnowej w regulacji lokalnego eksperymentalnego zapalenia opłucnej, wywołanego podaniem karageniny. W odczynie tym stwierdzono już po pierwszych godzinach od zainicjowania procesu zapalnego wzrost stężenia IL-1 w wysięku opłucnowym, a w dalszym okresie narastanie stężenia IL-6 i liczby różnorodnych form leukocytarnych oraz zwiększenie objętości wysięku opłucnowego [129].
W odczynie zapalnym wywołanym karageniną obserwowano ponadto zwiększony wychwyt adhezyjnych cząsteczek, takich jak: ICAM-1 (intercellular adhesion molecule 1) oraz P-selektyny, które odgrywają istotną rolę w napływie leukocytów do ogniska zapalnego. Ponadto w przebiegu procesu zapalnego przy braku ochronnego działania antyoksydantów może dochodzić do uszkodzeń DNA, zmniejszenia oddychania mitochondrialnego i obniżenia stężenia NAD+ [37]. Wykazano [13], że usunięcie opłucnowych makrofagów z ogniska zapalnego znacząco redukowało napływ neutrofilów do jamy opłucnowej oraz wzrost stężenia cytokin, tj. TNF-α, IL-6, wytwarzanych przez te komórki. Przeniesienie natomiast makrofagów nietransgenicznych częściowo odnawiało naciek neutrofilowy, co może wskazywać m.in. na rolę komórek leukocytarnych w wytwarzaniu cytokin prozapalnych. Zmniejszenie napływu komórek prozapalnych do ogniska zapalnego, powstawanie wysięku oraz wytwarzanie PGE2 uzyskuje się przez hamowanie cyklooksygenaz (COX-1 i COX-2). Zablokowanie COX-1 może powodować znaczne zmniejszenie odczynu zapalnego, co wykazały badania Gilroya [49]. Ostatnie badania wykazały swoistą aktywność prozapalną, którą ma głównie izoforma COX-2, która w przypadku ekspozycji na TCDD jest indukowana za pośrednictwem AhR w sposób przedstawiony na rycinie 8.

Ryc. 8. Mechanizm prozapalnego działania dioksyny przez stymulację COX-2; TCDD – dioksyny, AhR – receptor węglowodorów arylowych, Ca2+ – niezwiązane jony wapnia, cPlA2 – cytosolowa fosfolipaza A2, AA- kwas arachidonowy, COX2 – cyklooksygenaza 2
W wyniku migracji neutrofilów do ogniska zapalnego, regulowanej przez cytokiny, selektyny i integryny, które znajdują się na powierzchni śródbłonka naczyń krwionośnych, dochodzi do powstania stresu oksydacyjnego, związanego z wydzielaniem aktywnych metabolitów tlenu, tj. nadtlenków wodoru, tlenku azotu (NO) [27,37,123], które również powodują wzrost aktywności syntetazy tlenku azotu [37,38,123,138]. Dane literaturowe podkreślają prozapalną rolę NO, jednak według niektórych doniesień tlenek azotu może ograniczać nasilenie odczynu zapalnego, co należałoby wiązać z jego właściwością polegającą na zmniejszaniu oporu naczyniowego. Dowodem jest to, że zahamowanie syntetazy NO (NOS) przyczyniało się do zaostrzenia procesu zapalnego, w którym zaobserwowano wzrost histaminy i leukotrienu B4 [98].
Istnieją sugestie, że stres oksydacyjny powstały w doświadczalnym zapaleniu opłucnej, indukowanym karageniną, może być potencjalnym mechanizmem powstania zapalenia płuc. W przebiegu tego procesu dochodziło do gromadzenia płynu w przestrzeni opłucnowej, który zawierał dużą liczbę neutrofilów. Ponadto obserwowano infiltrację neutrofilów do tkanki płucnej, co przyczyniało się do powstania stresu oksydacyjnego powodującego peroksydację lipidów, przekształcenie azotynów w azotany oraz podwyższenie stężenia TNF, IL-1β [35], co również stwierdzono w badaniach cytowanych wcześniej autorów. Wzrost tych prozapalnych cytokin był związany z aktywacją przez anion nadtlenkowy jądrowego czynnika NF-κB (nuclear factor-κB), będącego transkrypcyjnym przekaźnikiem odgrywającym znaczącą rolę w indukcji genów zaangażowanych w odpowiedzi zapalnej [36]. Niedotlenienie towarzyszące odczynowi zapalnemu przyczynia się do aktywacji NF-κB (w wyniku degradacji jego inhibitora) i odgrywa zasadniczą rolę w transkrypcyjnym pobudzeniu genu IL-6, który może być również pobudzany przez inne bodźce, tj. IL-1 i TNF-α. Zmniejszenie aktywacji NF-κB indukowane przez hipoksję uzyskiwano przez podanie α-tokoferolu [85].
Charakter i dynamika odczynu zapalnego mogą być określone za pomocą wielu różnych wskaźników diagnostycznych (ryc. 7), których zachowanie charakteryzuje rodzaj reakcji zapalnej [14]. Toczący się odczyn zapalny wywiera również swój wpływ na niektóre parametry układu erytrocytarnego, które mogą być oceniane w analizie wskaźników hematologicznych. Również w procesie zapalnym istotną rolę odgrywa funkcja sekrecyjna wątroby, związana m.in. z wytwarzaniem białek ostrej fazy, biorących udział w reakcji zapalnej [14,50,128]. Monitorowanie niektórych wskaźników diagnostycznych wątroby (aktywności aminotransferaz AspAT, AlAT, stężenia cholesterolu) jest istotne do oceny zaburzenia jego funkcji w przypadku narażenia organizmu na oddziaływanie substancji szkodliwych, takich jak dioksyny (tab.2) [14].
Jednym z najważniejszych czynników antyoksydacyjnych jest tokoferol, dlatego w następnym rozdziale omówiono jego działanie ze szczególnym uwzględnieniem protekcyjnej funkcji mogącej zapobiegać negatywnym skutkom stresu oksydacyjnego, pojawiającego się zarówno w przebiegu odczynu zapalnego, jak i w wyniku oddziaływania dioksyn.
4. Rola tokoferolu w stresie oksydacyjnym
Grupa tokoferoli określana jako witaminy E reprezentowana jest przez wiele produktów kondensacji metylobenzochinonów z fitolem (ryc. 9). Wykazano, że w grupie tej osiem naturalnych tokoferoli cechuje się aktywnością biologiczną.
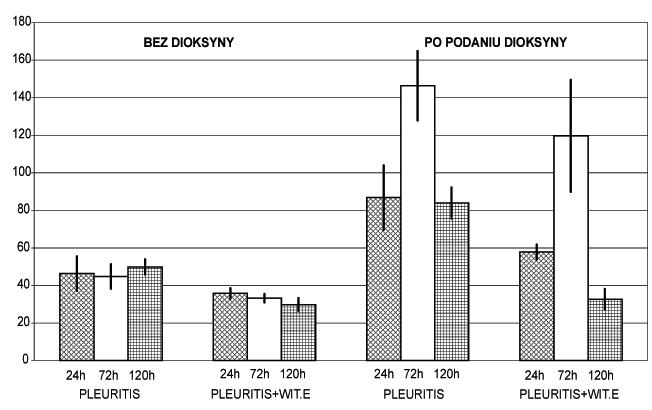
Ryc. 9. Wpływ tokoferolu na zmiany aktywności AlAT w surowicy w odczynie zapalnym (pleuritis) bez dioksyny i po podaniu dioksyny
W przemyśle farmaceutycznym jest produkowanych kilka pochodnych tokoferolu [110]:
• dl-α-tokoferol (2,5,7,8-tetrametylo-2-(4,8,12-tridecylo) chroman-6-ol,
• dl-α-tokoferol (2,5,7,8-tetrametylo-2-(4,8,12-tridecylo) chroman-6-ol,
• d-α-octan tokoferolu; dl-octan α-tokoferolu,
• d-bursztynylo α-tokoferol; dl-bursztynylo α-tokoferol,
• inne syntetyczne pochodne tokoferoli cechujące się aktywnością biologiczną o strukturze chemicznej objętej tajemnicą firmową.
Przy czym witamina E, jako środek farmakologiczny najczęściej jest stosowana jako d-α-octan tokoferolu, którego 1 mg ma aktywność 1,36 IU jednostki międzynarodowej (ryc. 10).

Ryc. 10. Wzór strukturalny α-tokoferolu
Witamina E rozpuszczona w tłuszczach jest wchłaniana w jelitach podczas trawienia lipidów. Wchłonięte chylomikrony docierają z krwią do tkanek zawierających lipazę lipoproteinową oraz do krążenia wrotnego wątroby jako chylomikrony resztkowe. Z wątroby transport tej witaminy do krwiobiegu odbywa się za pośrednictwem lipoprotein o małej gęstości (LDL). Istotną cechą tokoferolu, w odróżnieniu od innych witamin jest to, że witamina ta po wypełnieniu swojej funkcji antyoksydacyjnej nie ulega recyrkulacji, w związku z czym musi być zastąpiona nową dawką tokoferolu. Antyoksydacyjne działanie tokoferolu polega na zabezpieczeniu fosfolipidów błon mitochondrialnych, siateczki śródplazmatycznej i błon plazmatycznych przed utlenianiem wchodzących w ich skład wielonienasyconych kwasów tłuszczowych.
Niedobór tokoferolu zwiększa przepuszczalność błony lizosomalnej, przez co wzmożone jest przenikanie enzymów lizosomalnych w komórkach. Objawy niedoboru tokoferolu mogą się przejawiać keratynurią, zmniejszoną opornością osmotyczną, skróceniem czasu życia krwinek czerwonych, niedokrwistością hemolityczną i tworzeniem ciałek wtrętowych Heinza [7,60,126,136]. Przyczyną niedoboru witaminy E może być dysfunkcja wchłaniania w jelitach związana z zapaleniem trzustki, zwłóknieniem torbielowatym trzustki, resekcją żołądka, zespołem sprue, dietą zawierającą tłuszcze wielonienasycone [9,66,77]. U zwierząt niedobór witaminy E powoduje dystrofię mięśniową, tworzenie barwników lipidowych [47], niedokrwistości [44], zaburzenia przemiany kreatyny [48], zaburzenia płodności [67]. U człowieka prawidłowe stężenie tokoferolu w osoczu wynosi 0,7-1,6 mg/100 ml. Obniżenie stężenia poniżej 0,4 mg/100ml osocza wskazuje na niedobór tokoferolu. Pociąga to za sobą spadek oporności osmotycznej erytrocytów na związki działające utleniająco (np. nadtlenek wodoru). Zjawisko to jest wykorzystane w teście hemolizy według Rosego i Györgyego. W teście tym wartość hemolizy przekraczająca 25% wskazuje na hipowitaminozę E [71]. Przyjęto, że do utrzymania prawidłowego stężenia tokoferolu w osoczu jest niezbędne dostarczenie go w ilości 25-30 mg α-tokoferolu dziennie.
Stwierdzono, że niedobór witaminy E w przeciążeniach organizmu spowodowanych wysiłkiem fizycznym, odczynami zapalnymi, przyczynia się do znacznego wzrostu wolnych rodników, co przejawia się różnego rodzaju zaburzeniami, takimi jak: atrofia mięśni szkieletowych, zmiany dystroficzne w sercu, pojawienie się ognisk martwiczych w narządach miąższowych. Przy niedoborze tej witaminy występują ponadto uszkodzenia naczyń włosowatych oraz dochodzi do zwiększonej krzepliwości krwi. Również w okresie noworodkowym istnieje zapotrzebowanie na większą ilość pobieranego tokoferolu w związku z intensywnymi przemianami metabolicznymi, dlatego przy braku tej witaminy obserwowano obniżoną syntezą hemoglobiny i mniejszą przeżywalność erytrocytów. Wykazano znaczące korelacje między niskim stężeniem w surowicy a-tokoferolu a zwiększoną zachorowalnością na choroby sercowo-naczyniowe. Uważa się, że ochronne działanie tokoferolu zabezpieczające przed peroksydacją wielonienasyconych kwasów tłuszczowych jest związane z przeciwdziałaniem rozwojowi miażdżycy, nowotworów i uszkodzenia narządów miąższowych [11,40,97].
Powyższe obserwacje potwierdzono w badaniach przeprowadzonych na królikach z eksperymentalnie wywołaną arteriosklerozą, którym podawano tokoferol w karmie (w ilości 500 mg/kg pożywienia) przez 24 tygodnie. Doświadczenia te potwierdziły antyoksydacyjne działanie tej witaminy, co przejawiało się zmniejszoną infiltracją makrofagów w endotelium, brakiem kumulacji lipidów, stabilizacją płytki miażdżycowej i zmniejszeniem procesów apoptotycznych [78].
Stwierdzono, że witamina E w dawkach przekraczających kilkakrotnie dawkę optymalną (25-30 mg/dzień) nie wywiera znaczących efektów farmakologicznych [32], natomiast tokoferol stosowany w dużych dawkach wpływał na dojrzewanie płciowe królików i szczurów oraz zwiększał aktywność wydzielniczą tarczycy.
Antynowotworowe właściwości tokoferolu zaobserwowano w wyniku znacznego zmniejszenia mutagennego działania DMBA (7,12-dimetyloben-zantracenu) na zwierzęta [124], którym podawano witaminę E w dawce 10 mg/kg m.c. na dzień. Według niektórych doniesień antynowotworowe działanie α-tokoferolu może być związane ze spadkiem aktywności katepsyny B i L oraz wzrostem stężenia inhibitorów cysteinowych peptydaz [83,108]. Autorzy ci sugerują, że podawanie tokoferolu stymuluje makrofagi do wytwarzania IL-1, która pobudza fibroblasty do wydzielenia IL-6. Interleukina ta podwyższa stężenie kininogenu będącego inhibitorem cysteinowych peptydaz, co może również znacząco wpływać na przebieg odczynu zapalnego, niejednokrotnie towarzyszącego procesowi nowotworowemu.
W badaniach Lauritzena [76] przeprowadzonych na zwierzętach stwierdzono, że w przebiegu doświadczalnego bakteryjnego zapalenia płuc dochodziło do spadku stężenia kwasu askorbinowego i α-tokoferolu w surowicy. Powstały deficyt antyoksydantów przyczyniał się do zwiększenia stężenia wolnych rodników w przebiegu odczynu zapalnego. Według tych autorów wskaźniki, takie jak kwas askorbinowy i α-tokoferol mogą służyć do określania skutków i dynamiki leczenia procesu zapalnego.
Najnowsze badania wskazują na protekcyjne oddziaływanie witaminy E, stosowanej w większych dawkach celem niwelacji wolnych rodników powstających w stresie oksydacyjnym, uwarunkowanym różnymi czynnikami zapalnymi. W doświadczeniach na szczurach wykazano, że podawanie witaminy E w dawce 100 mg/kg m.c. przez 30 dni powodowało u nich spadek wolnych rodników w osoczu, wątrobie, jądrach i mózgu [39]. Eliminacja w organizmach myszy negatywnych skutków wzrostu stężenia wolnych rodników, które powstały w wyniku ekspozycji na dym tytoniowy, była związana z podawaniem tym zwierzętom dl-α-octanu tokoferolu (α-TA). W doświadczeniach tych wykazano, że w popłuczynach oskrzelowo-pęcherzykowych (BAL) myszy występowało zmniejszenie stężenia cytokiny IL-1β oraz podwyższenie stężenia IL-2 i IL-4. Podawanie tokoferolu zapobiegało ponadto peroksydacji lipidów w wątrobie, dysfunkcji płuc i redukcji masy ciała [134]. Również w doświadczeniach na szczurach [92] wykazano, że podawanie α-TA eliminowało negatywne skutki działania nikotyny, związane z wywoływaniem stresu oksydacyjnego. Przejawiało się to wspomaganiem przez tę witaminę regeneracji uszkodzonej tkanki kostnej u tych zwierząt. W badaniach tych Norazlina i wsp. wykazali też supresyjne działanie tokoferolu na wydzielanie cytokin prozapalnych IL-1 i IL-6 do surowicy, których stężenie wzrastało po uprzednim podaniu nikotyny w dawce 7 mg/kg przez okres 3 miesięcy.
Podanie dużych dawek tokoferolu powodowało obniżenie stężenia prozapalnych cytokin (IL-1, IL-6, TNF) wydzielanych przez monocyty do surowicy oraz obniżenie stężenia białka CRP (C-reactive protein) u diabetyków, u których stwierdzano predyspozycje do powstawania odczynów zapalnych. Ponadto dochodziło u nich – w wyniku stosowania tokoferolu – do obniżenia stężenia PAI-1 (plasminogen activator inhibitor 1) i peroksydowanych lipidów, powstałych pod wpływem stresu oksydacyjnego [64]. Powyższe wyniki sugerują, że podawanie tokoferolu stwarza dodatkową możliwość terapeutyczną zmniejszając natężenie odczynu zapalnego spowodowanego przez różne czynniki. Potwierdzają to badania własne, w których stwierdzono, że podawanie przez 3 tygodnie dużych dawek tokoferolu obniżało stężenia cytokin prozapalnych indukowanych podaniem TCDD i procesem zapalnym. W szczególności stwierdzono znaczące obniżenie stężenia TNF – ryc. 11 [15].

Ryc. 11. Wpływ tokoferolu na zmiany stężenia TNF w surowicy w odczynie zapalnym (pleuritis) bez dioksyny i po podaniu dioksyny
Obserwowano zmniejszenie wydzielania cytokin IL-1 i IL-6 oraz wolnych rodników, powstałych w wyniku podawania FeNTA (ferric nitrilotriacetate) szczurom, u których stosowano tokoferol w dawce 100 mg/kg m.c. [2]. Badania Devaraja i wsp. [40] wykazały znaczący spadek uwalniania reaktywnych form tlenu (anionu nadtlenkowego i nadtlenku wodoru) oraz zmniejszenie oksydacji lipidów, a także obniżenie stężenia IL-1β w surowicy u ochotników poddanych ośmiotygodniowej suplementacji α-AT w dawce 1200 UI/dzień. Ponadto w badaniach tych stwierdzono mniejszą aktywność kinazy C oraz zmniejszenie adhezji monocytów do śródbłonka naczyń krwionośnych. W wielu badaniach [112,113,125], w których wywoływano stres oksydacyjny różnymi metodami, m.in. indukcją odczynu zapalnego wykazano, że dochodziło do utleniania lipoprotein o niewielkiej gęstości, uwalniania cytokin prozapalnych IL-1β, IL-6 i TNF-α, chemokininy IL-8, inhibitora aktywatora plazminowego PAI-1, podwyższenia stężenia białka CRP oraz wzrostu adhezji monocytów do śródbłonka naczyń. Stosowanie różnych postaci tokoferolu zarówno u ludzi, jak i u zwierząt zmniejszało następstwa stresu oksydacyjnego związane z powstawaniem lipidów peroksydowanych, wytwarzaniem nadtlenku wodoru i anionu nadtlenkowego, zmniejszeniem uwalniania cytokin prozapalnych, chemokininy IL-8 oraz inhibitora aktywatora plazminogenu PAI-1. Podawanie tokoferolu działało supresyjnie na ekspresję m-RNA IL-6 i TNF-α, co było przyczyną obniżenia stężenia cytokin IL-6 i TNF-α [125]. Istotne jest brak obniżenia stężenia cytokin prozapalnych (IL-1β, IL-6, TNF-α) przez tokoferol przy niedoborze magnezu w organizmie [135].
Podawanie α-tokoferolu zmniejszało adhezję monocytów do endotelium oraz obniżało stężenie białka CRP w surowicy, hamowało kinazę białkową C, 5-lipooksygenazę oraz cyklooksygenazę 2 (COX-2). W związku z tym stosowanie α-AT może znacząco ograniczać odczyn zapalny poprzez wyhamowanie opisanych wcześniej mechanizmów i zapobiegać skutkom oksydacji. Niektórzy autorzy sugerują suplementację α-AT w dość dużych dawkach przez kilka tygodni [112].
Stres oksydacyjny jest decydującym sygnałem do fibrogenezy. Badania na myszach wykazały znaczący wpływ stresu oksydacyjnego na ekspresję genu wątrobowego odpowiedzialnego za syntezę kolagenu. Według niektórych autorów suplementacja lipofilnym antyoksydantem, którym jest tokoferol stosowany dłużej (8 tyg. w dawce 40 UI), bądź krócej (450 UI/48 godzin) spowodowała zmniejszenie ilości mRNA kolagenu o około 70% przy długim stosowaniu tej witaminy, zaś o 60% przy krótkim (ryc. 12) [30].

Ryc. 12. Wpływ tokoferolu na zmiany stężenia PICP (C-końcowy propeptyd prokolagenu typu I) w surowicy w odczynie zapalnym (pleuritis) bez dioksyny i po podaniu dioksyny
W najnowszych badaniach epidemiologicznych jest podnoszona kwestia protekcyjnego oddziaływania witaminy E, stosowanej w większych dawkach w celu niwelacji wolnych rodników powstających w stresie oksydacyjnym uwarunkowanym różnymi czynnikami zapalnymi i zatruciami, np. dioksynami [134]. W badaniach Hassoun i wsp. [57,58] wykazano, że dioksyny podawane szczurom przez 90 dni w sumarycznej dawce 5 µg/kg m.c., przyczyniały się do powstania stresu oksydacyjnego, który określano za pomocą pomiarów produktów nadtlenku azotu, peroksydacji lipidów i występowania uszkodzonych pojedynczych nici DNA. Wykazano, że jednoczesne podawanie przez 90 dni witaminy E działało protekcyjnie, zabezpieczając mózg przed produktami oksydacji.
Możliwość antyoksydacyjnego działania witaminy E wykazano w doświadczeniach na myszach, którym ją podawano jednocześnie z TCDD [3]. Witaminę E podano jednorazowo w ilości 150 mg/kg m.c., a następnie przez 5 kolejnych dni w ilości 40 mg m.c., przy czym w czwartym dniu zastosowano pojedynczą dawkę TCDD w ilości 50 µg/kg m.c. Wykazano, że u zwierząt poddanych jednoczesnemu działaniu witaminy E oraz TCDD nastąpiło znaczące zmniejszenie objawów zatrucia dioksynami, do których zalicza się spadek ogólnej masy ciała oraz obniżenie masy grasicy. Obserwowano ponadto zmniejszone wytwarzanie nadtlenku azotu przez komórki płynu otrzewnowego, jak i zmniejszenie częstości występowania uszkodzeń pojedynczych nici DNA. Badania na samicach szczurów, u których zastosowano letalną dawkę TCDD i jednocześnie podawano witaminę E wykazały znaczące zahamowanie peroksydacji mikrosomalnych lipidów oraz 10% wzrost przeżywalności zwierząt [118].
5. Podsumowanie
Na podstawie uzyskanych wyników w badaniach własnych [15] dłuższe stosowanie tokoferolu w dużych dawkach przyczynia się do obniżenia interleukin prozapalnych w surowicy ze szczególną redukcją wzrostu TNF występującego u zwierząt poddanych działaniu TCDD, jak i obniżeniem aktywności ALAT, co jest dowodem usprawnienia funkcji wątroby.
PIŚMIENNICTWO
[1] Ahlborg U.G., Brouwer A., Fingerhut M.A., Jacobson J.L., Jacobson S.W., Kennedy S.W., Kettrup A.A., Koeman J.H., Poiger H., Rappe C.: Impact of polychlorinated dibenzo-p-dioxins, dibenzofurans, and biphenyls on human and environmental health, with special emphasis on application of the toxic equivalency factor concept. Eur. J. Pharmacol., 1992; 228: 179-199
[PubMed]
[2] Ahmad N.S., Khalid B.A., Luke D.A., Ima Nirwana S.: Tocotrienol offers better protection than tocopherol from free radical-induced damage of rat bone. Clin. Exp. Pharmacol. Physiol., 2005; 32: 761-770
[PubMed]
[3] Alsharif N.Z., Hassoun E.A.: Protective effects of vitamin A and vitamin E succinate against 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD)-induced body wasting, hepatomegaly, thymic atrophy, production of reactive oxygen species and DNA damage in C57BL/6J mice. Basic Clin. Pharmacol. Toxicol., 2004; 95: 131-138
[PubMed]
[4] Andreasen E.A., Mathew L.K., Löhr C.V., Hasson R., Tanguay R.L.: Aryl hydrocarbon receptor activation impairs extracellular matrix remodeling during zebra fish fin regeneration. Toxicol. Sci., 2007; 95: 215-226
[PubMed] [Full Text HTML] [Full Text PDF]
[5] Baccarelli A., Pesatori A.C., Consonni D., Mocarelli P., Patterson D.G., Caporaso N.E., Bertazzi P.A., Landi M.T.: Health status and plasma dioxin levels in chloracne cases 20 years after the Seveso, Italy accident. Br. J. Dermatol., 2005; 152: 459-465
[PubMed]
[6] Bauman H., Gauldie J.: The acute-phase response. Immunol. Today, 7512342
[PubMed]
[7] Binder H.J., Herting D.C., Hurst V., Finch S.C., Spiro H.M.: Tocopherol deficiency in man. N. Engl. J. Med., 1965; 273: 1289-1297
[PubMed]
[8] Boverhof D.R., Burgoon L.D., Tashiro C., Sharratt B., Chittim B., Harkema J.R., Mendrick D.L., Zacharewski T.R.: Comparative toxicogenomic analysis of the hepatotoxic effects of TCDD in Sprague Dawley rats and C57BL/6 mice. Toxicol. Sci., 2006; 94: 398-416
[PubMed] [Full Text HTML] [Full Text PDF]
[9] Braunstein H.: Tocopherol deficiency in adults with chronic pancreatitis. Gastroenterology, 1961; 40: 224-231
[10] Bruner-Tran K.L., Osteen K.G.: Dioxin-like PCBs and endometriosis. Syst. Biol. Reprod. Med., 2010; 56: 132-146
[PubMed] [Full Text HTML] [Full Text PDF]
[11] Burdon R.H., Gill V., Rice-Evans C.: Oxidative stress and tumour cell proliferation. Free Radic. Res. Commun., 1990; 11: 65-76
[PubMed]
[12] Burton J.E., Michalek J.E., Rahe A.J.: Serum dioxin, chloracne, and acne in veterans of Operation Ranch Hand. Arch. Environ. Health., 1998; 5: 199-204
[PubMed]
[13] Cailhier J.F., Sawatzky D.A., Kipari T., Houlberg K., Walbaum D., Watson S., Lang R.A., Clay S., Kluth D., Savill J., Hughes J.: Resident pleural macrophages are key orchestrators of neutrophil recruitment in pleural inflammation. Am. J. Respir. Crit. Care Med., 2006; 173: 540-547
[PubMed] [Full Text HTML] [Full Text PDF]
[14] Całkosiński I.: Przebieg doświadczalnego zapalenia opłucnej po stosowaniu nitrogranulogenu (NTG) i 2,3,7,8- tetrachlorodibenzo-p-dioksyny (TCDD). Akademia Medyczna we Wrocławiu, Wrocław 2005
[15] Całkosiński I.: Wpływ tokoferolu na diagnostyczne wskaźniki odczynu zapalnego u szczurów poddanych dzialaniu dioksyny. Akademia Medyczna we Wrocławiu, Wrocław 2008
[16] Całkosiński I., Borodulin-Nadzieja L., Stańda M., Wasilewska U., Cegielski M.: Wpływ jednorazowej dawki TCDD na poziom estrogenów i reprodukcję u samic szczurów. Med. Wet., 2003; 59: 536-538
[17] Całkosiński I., Borodulin-Nadzieja L., Stańda M., Wasilewska U., Pietraszkiewicz T.: Wpływ magnetostymulacji leczniczej na stężenie pochodnych kolagenu w przebiegu doświadczalnego zapalenia opłucnej. Med. Wet., 2003; 59: 161-164
[18] Całkosiński I., Cegielski M., Dzięgiel P., Majda J.: Apoptotic changes in the myocardium in the course of experimentally-induced pleurisy. Folia Morphol., 2004; 63: 225-228
[PubMed]
[19] Całkosiński I., Dobrzyński M., Cegielski M., Sieja A., Całkosińska M.: Wielokierunkowe działanie 2,3,7,8-tetrachlorodibenzo-p-dioksyny (TCDD) na organizm ze szczególnym uwzględnieniem zmian w uzębieniu. Postępy Hig. Med. Dośw., 2006; 60: 237-240
[PubMed] [Full Text HTML] [Full Text PDF]
[20] Całkosiński I., Majda J., Borodulin-Nadzieja L., Wasilewska U., Stańda M., Skalik R.: Wpływ zapalenia opłucnej na zmiany gospodarki lipidowo-cholesterolowej i poziom hormonów płciowych u szczurów. Med. Wet., 2004; 60: 990-993
[21] Całkosiński I., Rosińczuk-Tonderys J., Gamian A., Chruszcz A.: Zmiany skórne i metaboliczne organizmu w zatruciu dioksynami. Katastrofy naturalne i cywilizacyjne, różne oblicza bezpieczeństwa. Wydawnictwo Wyższa Szkoła Oficerska Wojsk Lądowych im. gen. T. Kościuszki, Wrocław 2010; 355-369
[22] Całkosiński I., Skalik R., Borodulin-Nadzieja L., Wasilewska U., Janocha A., Cegielski M., Ponikowska B., Goździk A.; Influence of inflammatory reaction on blood concentration of cholesterol and other biochemical values with regard to cardiac muscle damage in rats. Bull. Vet. Instit. Pulawy, 2004; 48: 477-480
[23] Całkosiński I., Stańda M., Borodulin-Nadzieja L., Wasilewska U., Majda J., Cegielski, M., Dzięgiel P.: Wpływ dioksyn na procesy rozrodcze u szczurów w badaniach in vivo. Adv. Clin. Exp. Med., 2004; 13: 875-890
[Abstract]
[24] Całkosiński I., Stańda M., Borodulin-Nadzieja L., Wasilewska U., Majda J., Cegielski, M., Dzięgiel P.: Wpływ 2,3,7,8-tetrachlorodibenzo-p-dioksyny (TCDD) na strukturę narządów miąższowych oraz na stężenie cholesterolu i estradiolu u szczurów. Adv. Clin. Exp. Med., 2005; 14: 211-215
[25] Całkosiński I., Stańda M., Purzyc L., Pietraszkiewicz T., Borodulin-Nadzieja L.: Emisja dioksyn do środowiska i ich wpływ na parametry fizjologiczne organizmu. Materiały IX Konferencji Naukowej “Dziecko w środowisku zagrożonym ekologicznie”. Legnica, 2-3 czerwca 2000 Fund. Zagłębia Miedziowego, 5: 49-50
[26] Całkosiński I., Wasilewska U., Borodulin-Nadzieja L., Cegielski M., Dzięgiel P., Stańda M., Majda J.: Wpływ 2,3,7,8-tetrachlorodibenzo-p-dioksyny (TCDD) na czynność i strukturę jajników. Med. Wet., 2004; 60: 1218-1221
[27] Carp H., Janoff A.: Potential mediator of inflammation. Phagocyte-derived oxidants suppress the elastase-inhibitory capacity of alpha-1-proteinase inhibitors in vitro. J. Clin. Invest., 1980; 66: 987-995
[PubMed] [Full Text HTML] [Full Text PDF]
[28] Chaffin C.L., Heimler I., Rawlins R.G., Wimpee B.A., Sommer C., Hutz R.J.: Estrogen receptor and aromatic hydrocarbon receptor in the primate ovary. Endocrine, 1996; 5: 315-321
[PubMed]
[29] Chaffin C.L., Hutz R.J.: Regulation of the aromatic hydrocarbon receptor (AHR) by in utero and lactational exposure to 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD). J. Reprod. Dev., 1997; 43: 47-51
[30] Chojkier M., Houglum K., Lee K.S., Buck M.: Long- and short-term D-alpha-tocopherol supplementation inhibits liver collagen alpha1 (I) gene expression. Am. J. Physiol., 1998; 275: 1480-1485
[PubMed] [Full Text HTML] [Full Text PDF]
[31] Chrousos G.P.: The hypothalamic-pituitary-adrenal axis and immune-mediated inflammation. N. Engl. J. Med., 1995; 332: 1351-1362
[PubMed]
[32] Cohn V.H.: Fat-solube vitains. III. Vitamin K and vitamin E. W: Goodman L.S., Gilman A.: The Pharmacological Basis of Therapeutics, 4th Ed. London-Toronto: Collier-Macmillan Company 1970; 1690-1699
[33] Crofford L.J., Sano H., Karalis K., Webster E.L., Goldmuntz E.A., Chrousos G.P., Wilder R.L.: Local secretion of corticotropin-releasing hormone in the joints of Lewis rats with inflammatory arthritis. J. Clin. Invest., 1992; 90: 2555-2564
[PubMed] [Full Text HTML] [Full Text PDF]
[34] Crow K.: Chloracne. Trans. St. Jonhs Hosp. Dermatol. Soc., 1970; 56: 77-99
[35] Cuzzocrea S., Mazzon E., Dugo L., Serraino I., Ciccolo A., Centorrino T., De Sarro A., Caputi A.P.: Protective effects of N-acetylcysteine on lung injury and red blood cell modification induced by carrageenan in the rat. FASEB. J., 2001; 15: 1187-1200
[PubMed] [Full Text HTML] [Full Text PDF]
[36] Cuzzocrea S., Pisano B., Dugo L., Ianaro A., Ndengele M., Salvemini D.: Superoxide-related signaling cascade mediates nuclear factor-kappaB activation in acute inflammation. Antioxid. Redox Signal., 2004; 6: 699-704
[PubMed]
[37] Cuzzocrea S., Zingarelli B., Hake P., Salzman A.L., Szabo C.: Antiinflammatory effects of mercaptoethylguanidine, a combined inhibitor of nitric oxide synthase and peroxynitrite scavenger, in karageenan-induced models of inflammation. Free Radic. Biol. Med., 1998; 24: 450-459
[PubMed]
[38] Dąbrowska M.: Rola płytek krwi w zapaleniu. Diagn. Lab., 1997; 33: 429-433
[39] el-Demerdash F.M., Yousef M.I., Kedwany F.S., Baghdadi H.H.: Role of alpha-tocopherol and beta-carotene in ameliorating the fenvalerate-induced changes in oxidative stress, hemato-biochemical parameters, and semen quality of male rats. J. Environ. Sci. Health B., 2004; 39: 443-459
[PubMed]
[40] Devaraj S., Li D., Jialal I.: The effects of alpha tocopherol supplementation on monocyte function. Decreased lipid oxidation, interleukin 1 beta secretion, and monocyte adhesion to endothelium. J. Clin. Invest., 1996; 98: 756-763
[PubMed] [Full Text HTML] [Full Text PDF]
[41] Esser C, Rannug A, Stockinger B.: The aryl hydrocarbon receptor in immunity. Trends Immunol., 2009; 30: 447-454
[PubMed]
[42] Falandysz J.: Polichlorowane dibenzo-p-dioksyny w żywności. Bromat. Chem. Toksykol., 1989; 22: 3-4
[43] Fiedler H.: National PCDD/PCDF release inventories under the Stockholm Convention on Persistent Organic Pollutants. Chemosphere., 2007; 67: S96-S108
[PubMed]
[44] Fitch C.D.: Experimental anemia in primates due to vitamin E deficiency. Vitam. Horm., 1968; 26: 501-514
[PubMed]
[45] Fowler B.A., Lucier G.W., Brown H.W., McDaniel O.S.: Ultrastructural changes in rat liver cells following a single oral dose of TCDD. Environ. Health Perspect., 1973; 5: 141-148
[46] Gaborek B.J., Mullikin J.M., Pitrat A.T., Cummings L., May L.M.: Pentagon surface wipe sampling health risk assessment. Toxicol. Ind. Health., 2001; 17: 254-261
[PubMed]
[47] Gedigk P., Wessel W.: Elektronenmikroskopische Untersuchungen des Vitamin E-Mangel-Pigments im Myometrium der Ratte. Virchows Arch. Pathol. Anat. Physiol. Klin. Med., 1964; 337: 367-382
[PubMed]
[48] Gerber G.B., Gerber G., Koszalka T.R., Emmel V.M.: Creatin metabolism in vitamin E deficiency in the rat. Am. J. Physiol., 1962; 202: 453-460
[PubMed]
[49] Gilroy D.W., Tomlinson A., Willoughby D.A.: Differential effects of inhibitors of cyclooxygenase (cyclooxygenase 1 and cyclooxygenase 2) in acute inflammation. Eur. J. Pharmacol., 1998; 355: 211-217
[PubMed]
[50] Gordon A.H., Koj A.: The acute phase response to injury and infection. Elsevier/North Holland Biomedical Press. Amsterdam, New York, Oxford., 1985; 139-144
[51] Gregoraszczuk E.L., Zabielny E., Ochwat D.: Aryl hydrocarbon receptor (AhR)-linked inhibition of luteal cell progesterone secretion in 2,3,7,8-tetrachlorodibenzo-p-dioxin treated cells. J. Physiol. Pharmacol., 2001; 2: 303-311
[PubMed] [Full Text PDF]
[52] Gregoraszczuk E.L., Zabielny E., Pieklo R., Grochowalski A., Wójtowicz A., Mika R.: Effects of 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD) on luteal cell function. Tissue culture approach. Organohalogen. Compounds, 1999; 42: 67-71
[53] Grochowalski A.: Dioksyny. V Międzynarodowa Konferencja: Dioksyny w Przemyśle i Środowisku, Kraków, 2001; 1-38
[54] Grochowalski A., Chrząszcz R.: Determination of PCDFs/PCDDs in ambient air from. Cracow city, Poland. Organohal. Comp., 1995; 21: 321-326
[55] Harbuz M.S., Rees R.G., Eckland D., Jessop D.S, Brewerton D, Lightman S.L.: Paradoxical responses of hypothalamic corticotropin-releasing factor (CRF) messenger ribonucleic acid (mRNA) and CRF-41 peptide and adenohypophysial proopiomelanocortin mRNA during chronic inflammatory stress. Endocrinology, 1992; 130: 1394-1400
[PubMed]
[56] Harris M., Zacharewski T., Safe S.: Comparative potencies of Aroclor 1232, 1242, 1248, 1254 and 1260 in male wistar rats-assessment of the toxic equivalency factor (TEF) approach for polychlorinated biphenyls (PCBs). Fundam. Appl. Toxicol., 1993; 20: 456-463
[PubMed]
[57] Hassoun E.A., Vodhanel J., Abushaban A.: The modulatory effects of ellagic acid and vitamin E succinate on TCDD-induced oxidative stress in different brain regions of rats after subchronic exposure. J. Biochem. Mol. Toxicol., 2004; 18: 196-203
[PubMed]
[58] Hassoun E.A., Vodhanel J., Holden B., Abushaban A.: The effects of ellagic acid and vitamin E succinate on antioxidant enzymes activities and glutathione levels in different brain regions of rats after subchronic exposure to TCDD. J. Toxicol. Environ. Health A., 2006; 69: 381-393
[PubMed]
[59] Holsapple M.P., Snyder N.K., Wood S.C., Morris D.L.: A review of 2,3,7,8-tetrachlorodibenzo-p-dioxin-induced changes in immunocompetence. Toxicology, 1991; 69: 219-255
[PubMed]
[60] Horwitt M.K., Century B., Zeman A.: Erythrocyte survival time and reticulocyte levels after tocopherol depletion in man. Am. J. Clin. Nutr., 1963; 12: 99-106
[PubMed] [Full Text PDF]
[61] Hwang S.W.: Effect of 2,3,7,8-tetrachlorodibenzo-p-dioxin on the biliary excrection of indocyanide-Green in rats. Environ. Health Perspect., 1973; 5: 227-231
[PubMed] [Full Text HTML] [Full Text PDF]
[62] Ivens I.A., Loser E., Rinke M., Schmidt U., Neupert M.: Toxicity of 2,3,7,8-tetrabromodibenzo-p-dioxin in rats after single oral administration. Toxicology, 1992; 73: 53-69
[PubMed]
[63] Jensen B.A., Leeman R.J., Schlezinger J.J., Sherr D.H.: Aryl hydrocarbon receptor (AhR) agonists suppress interleukin-6 expression by bone marrow stromal cells: an immunotoxicology study. Environ Health., 2003; 2: 16
[PubMed] [Full Text HTML] [Full Text PDF]
[64] Jialal I., Devaraj S., Venugopal S.K.: Oxidative stress, inflammation, and diabetic vasculopathies: the role of alpha tocopherol therapy. Free Radic. Res., 2002; 36: 1331-1336
[PubMed]
[65] Karras J.G., Conrad D.H., Holsapple M.P.: Effects of 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD) on interleukin-4-mediated mechanisms of immunity. Toxicol. Lett., 1995; 75: 225-233
[PubMed]
[66] Kayden H.J., Silber R., Kossmann C.E.: The role of vitamin E deficiency in the abnormal autohemolysis of acanthocytosis. Trans. Ass. Amer. Phycns., 1965; 78: 334-342
[PubMed]
[67] Kenney M.A., Roderuck C.E.: Fatal syndrome associated with vitamin E status of pregnant rats. Proc. Soc. Exp. Biol. Med., 1963; 114: 257-261
[PubMed]
[68] Kerkvliet N.I., Brauner J.A.: Flow cytometric analysis of lymphocyte subpopulations in the spleen and thymus of mice exposed to an acute immunosuppressive dose of 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD). Environ. Res., 1990; 52: 146-154
[PubMed]
[69] Kociba R.J., Schwetz B.A.: Toxicity of 2,3,7,8-tetrachlorodibenzo-p-diokxin (TCDD). Drug Metab. Rev., 1982; 13: 387-406
[PubMed]
[70] Kostro K., Gliński Z., Wojcicka-Lorenowicz K., Krakowski L.: Immunologiczne i immunopatologiczne mechanizmy zapalenia. Med. Wet., 2000; 56: 479-485
[71] Körner W.F.: Chemische Analysen bei Hypo- und Hypervitaminosen. W: Opitz H., Schmid F.: Handbuch der Kinderheilkunde, Bd. 2/1. Berlin-Heidelberg- New York. Springer-Verlag 1966; 821-831
[72] Krauthacker B., Reiner E., Lindstrom G., Rappe C.: Residues of polychlorinated-dibenzodioxins, -dibenzofurans and -biphenyls in human milk samples collected in a continental town in Croatia, Yugoslavia. Arh. Hig. Rada. Toksikol., 1989; 40: 9-14
[PubMed]
[73] Kruzel M.L.: Rola laktoferyny w rozwoju ostrych stanów zapalnych Postępy Hig. Med. Dośw., 2003; 57: 377-404
[PubMed]
[74] Latchoumycandane C., Chitra K.C., Mathur P.P.: 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD) induces oxidative stress in the epididymis and epididymal sperm of adult rats. Arch. Toxicol., 2003; 77: 280-284
[PubMed]
[75] Latchoumycandane C., Mathur P.P.: Effects of vitamin E on reactive oxygen species-mediated 2,3,7,8-tetrachlorodi-benzo-p-dioxin toxicity in rat testis. J. Appl. Toxicol., 2002; 22: 345-351
[PubMed]
[76] Lauritzen B., Lykkesfeldt J., Skaanild M.T., Angen O., Nielsen J.P., Friis C.: Putative biomarkers for evaluating antibiotic treatment: an experimental model of porcine Actinobacillus pleuropneumoniae infection. Res. Vet. Sci., 2003; 74: 261-270
[PubMed]
[77] Leonard P.J., Losowsky M.S., Pulvertaft C.N.: Vitamin E deficiency. Brit. Med. J., 1966; 1: 1301-1302
[78] Li W., Hellsten A., Jacobsson L.S., Blomqvist H.M., Olsson A.G., Yuan X.M.: Alpha-tocopherol and astaxanthin decrease macrophage infiltration, apoptosis and vulnerability in atheroma of hyperlipidaemic rabbits. J. Mol. Cell. Cardiol., 2004; 37: 969-978
[PubMed]
[79] Li W., Matsumura F.: Significance of the nongenomic, inflammatory pathway in mediating the toxic action of TCCD to induce rapid and long-term cellular responses in 3T3-L1 adipocytes. Biochemistry, 2008; 47: 13997-14008
[PubMed]
[80] Li X., Johnson D.C., Rozman K.K.: Effects of 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD) on estrous cyclicyty and ovulation in female Sprague-Dawley rats. Toxicol. Lett., 1995; 78: 219-222
[PubMed]
[81] Liem A.K., Van Zorge J.A.: Dioxins and related compounds: status and regulatory aspects in selected countries. Environ. Sci. Pollut. Res., 1995; 2: 46-56
[82] Mai T.A., Doan T.V., Tarradellas J., de Alencastro L.F., Grandjean D.: Dioxin contamination in soils of Southern Vietnam. Chemosphere, 2007; 67: 1802-1807
[PubMed]
[83] Minakata K., Asano M., Sato T., Harada N.: Assay of alpha-cysteine proteinase inhibitor in serum or plasma. Hoppe Seylers Z. Physiol. Chem., 1982; 363: 493-498
[PubMed]
[84] Mniszek W., Wołek B.: Przegląd i ocena wyników prac badawczo-rozwojowych i publikacji dotyczących oddziaływania TZO na zdrowie i środowisko, w szczególności na Górnym i Dolnym Śląsku. Raport GF/POL/INV/R.17. Instytut Medycyny Pracy i Zdrowia Środowiskowego w Sosnowcu. Październik 2002
[85] Muraoka K., Shimizu K., Sun X., Zhang Y.K., Tani T., Hashimoto T., Yagi M., Miyazaki I., Yamamoto K.: Hypoxia, but not reoxygenation, induces interleukin 6 gene expression through NF-kappaB activation. Transplantation, 1997; 63: 466-470
[PubMed]
[86] Neal R.A., Beatty P.W., Gasiewicz T.A.: Studies of the mechanisms of toxicity of 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD). Ann. N.Y. Acad. Sci., 1979; 320: 204-213
[PubMed]
[87] Near R.I., Matulka R.A., Mann K.K., Gogate S.U., Trombino A.F., Sherr D.H.: Regulation of preB cell apoptosis by aryl hydrocarbon receptor/transcription factor-expressing stromal/adherent cells. Proc. Soc. Exp. Biol. Med., 1999; 221: 242-252
[PubMed]
[88] Neuberger M., Landvoigt W., Derntl F.: Blood levels of 2,3,7,8-tetrachlorodibenzo-p-dioxin in chemical workers after chloracne and in comparison groups. Int. Arch. Occup. Environ. Health, 1991; 63: 325-327
[PubMed]
[89] Neubert R., Golor G., Stahlmann R., Helge H., Neubert D.: Polyhalogenated dibenzo-p-dioxins and dibenzofurans and the immune system. 4. Effects of multiple-dose treatment with 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD) on peripheral lymphocyte subpopulations of a non-human primate (Callithrix jacchus). Arch. Toxicol., 1992; 66: 250-259
[PubMed]
[90] Neubert R., Jacob-Müller U., Stahlmann R., Helge H., Neubert D.: Polyhalogenated dibenzo-p-dioxins and dibenzofurans and the immune system. 1. Effects on peripheral lymphocyte subpopulations of a non-human primate (Callithrix jacchus) after treatment with 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD). Arch. Toxicol., 1990; 64: 345-359
[PubMed]
[91] Nottebrock C., Riecke K., Kruse M., Shakibaei M., Stahlmann R.: Effects of 2,3,7,8-tetrachloro-dibenzo-p-dioxin on the extracellular matrix of the thymus in juvenile marmosets (Callithrix jacchus). Toxicology, 2006; 226: 197-207
[PubMed]
[92] Norazlina M., Lee P.L., Lukman H.I., Nazrun A.S., Ima-Nirwana S.: Effects of vitamin E supplementation on bone metabolism in nicotine-treated rats. Singapore Med. J., 2007; 48: 195-199
[PubMed] [Full Text PDF]
[93] Oberemm A., Meckert C., Brandenburger L., Herzig A., Lindner Y., Kalenberg K., Krause E., Ittrich C., Kopp-Schneider A., Stahlmann R., Richter-Reichhelm H.B., Gundert-Remy U.: Differential signatures of protein expression in marmoset liver and thymus induced by single-dose TCDD treatment. Toxicology, 2005; 206: 33-48
[PubMed]
[94] Oehme M., Biseth A., Schlabach M., Wiig O.: Concentrations of polychlorinated dibenzo-p-dioxins, dibenzofurans and non-ortho substituted biphenyls in polar bear milk from Svalbard (Norway). Environ. Pollut., 1995; 90: 401-407
[PubMed]
[95] Oehme M., Schlabach M., Hummert K., Luckas B., Nordoy E.S.: Determination of levels of polychlorinated dibenzo-p-dioxins, dibenzofurans, biphenyls and pesticides in harp seals from the Greenland Sea. Sci. Total Environ., 1995; 162: 75-91
[PubMed]
[96] Olivero-Verbel J., Cabarcas-Montalvo M., Ortega-Zúniga C.: Theoretical targets for TCDD: A bioinformatics approach. Chemosphere, 2010; 80: 1160-1166
[PubMed]
[97] Pacanis A.: Witaminy antyoksydacyjne w prewencji miażdżycy. Diagn. Lab., 1996; 32: 589-595
[98] Paul-Clark M.J., Gilroy D.W., Willis D., Willoughby D.A., Tomlinson A.: Nitric oxide synthase inhibitors have opposite effects on acute inflammation depending on their route of administration. J. Immunol., 2001; 166: 1169-1177
[PubMed] [Full Text HTML] [Full Text PDF]
[99] Piskorska-Pliszczyńska J.: Toksyczność i mechanizm działania dioksyn. Med. Wet., 1996; 22: 94-98
[100] Piskorska-Pliszczyńska J.: Dioksyny i związane z nimi zagrożenia zdrowia. Med. Wet., 1999; 55: 491-496
[101] Piskorska-Pliszczyńska J.: Dioksyny i inne fałszywe hormony. Mat. VI Konf. Nauk. “Dioksyny w przemyśle i środowisku”. Kraków-Tomaszowice 26- 27.09.2002; 114-119
[102] Piskorska-Pliszczyńska J., Morrison V., Zacharewski T., Safe S.: DNA binding properties of the Ah receptor in wild-type and wariant mouse hepatoma cells. Arch. Biochem. Biophys., 1991; 290: 362-368
[PubMed]
[103] Pluim H.J., Koppe J.G., Olie K., Vd. Slikke J.W., Kok J.H., Vulsma T., Van Tijn D., De Vijlder J.J.: Effects of dioxins on thyroid function in newborn babies. Lancet, 1992; 339: 1303-1304
[PubMed]
[104] Rayne S.: Using exterior building surface films to assess human exposure and health risks from PCDD/Fs in New York City, USA, after the World Trade Center attacks. J. Hazard. Mater., 2005; 127: 33-39
[PubMed]
[105] Rayne S., Ikonomou M.G., Butt C.M., Diamond M.L., Truong J.: Polychlorinated dioxins and furans from the World Trade Center attacks in exterior window films from lower Manhattan in New York City. Environ. Sci. Technol., 2005; 39: 1995-2003
[PubMed]
[106] Riecke K., Grimm D., Shakibaei M., Kossmehl P., Schulze-Tanzil G., Paul M., Stahlmann R.: Low doses of 2,3,7,8-tetrachlorodibenzo-p-dioxin increase transforming growth factor beta and cause myocardial fibrosis in marmosets (Callithrix jacchus). Arch. Toxicol., 2002; 76: 360-366
[PubMed]
[107] Rodriguez-Sosa M., Elizondo G., Lopez-Duran R.M., Rivera I., Gonzalez F.J., Vega L.: Over-production of IFN-gamma and IL-12 in AhR-null mice. FEBS. Lett., 2005; 579: 6403-6410
[PubMed]
[108] Sakamoto W., Yoshikawa K., Shindoh M., Amemiya A., Handa H., Saeki T., Nagasawa S., Koyama J., Ogihara T., Mino M.: In vivo effects of vitamin E on peritoneal macrophages and T-kininogen level in rats. Int. J. Vitam. Nutr. Res., 1989; 59: 131-139
[PubMed]
[109] Schwetz B.A., Norris J.M., Sparschu G.L., Rowe U.K., Gehring P.J., Emerson J.L., Gerbig C.G.: Toxicology of chlorinated dibenzo-p-dioxins. Environ. Health Perspect., 1973; 5: 87-99
[PubMed] [Full Text HTML] [Full Text PDF]
[110] Siewiński M., Han S., Antonowicz J., Andrzejak R., Mikulewicz W., Saleh Y., Sebzda T., Kielan W., Zasławski R., Janocha A., Murawski M.: Perspektywy zastosowania dużych dawek witaminy E. Adv. Clin. Exp. Med., 2000; 9: 171-178
[111] Singh S.U., Casper R.F., Fritz P.C., Sukhu B., Ganss B., Girard B., Savouret J.F., Tenenbaum H.C.: Inhibition of dioxin effects on bone formation in vitro by a newly described aryl hydrocarbon receptor antagonist, resveratrol. J. Endocrinol., 2000; 167: 183-195
[PubMed] [Full Text PDF]
[112] Singh U., Devaraj S., Jialal I.: Vitamin E, oxidative stress, and inflammation. Annu. Rev. Nutr., 2005; 25: 151-174
[PubMed]
[113] Singh U., Jialal I.: Anti-inflammatory effects of alpha-tocopherol. Ann. N.Y. Acad. Sci., 2004; 1031: 195-203
[PubMed]
[114] Sonne C., Leifsson P.S., Dietz R., Born E.W., Letcher R.J., Hyldstrup L., Riget F.F., Kirkegaard M., Muir D.C.: Xenoendocrine pollutants may reduce size of sexual organs in East Greenland polar bears (Ursus maritimus). Environ. Sci. Technol., 2006; 40: 5668-5674
[PubMed]
[115] Sorg O., Zennegg M., Schmid P., Fedosyuk R., Valikhnovskyi R., Gaide O., Kniazevych V., Saurat J.H.: 2,3,7,8-tetrachlorodibenzo- p- dioxin (TCDD) poisonong In Victor Yushchenko; identyfication and measurement of TCDD metabolitem. Lancet, 2009; 374: 1179-1185
[PubMed]
[116] Starek A.: Ryzyko zdrowotne związane ze spalaniem odpadów komunalnych. Med. Pr., 2005; 56: 55-62
[117] Sterling J.B., Hanke C.W: Dioxin toxicity and chloracne in the Ukraine. J. Drugs Dermatol., 2005; 4: 148-150
[PubMed]
[118] Stohs S.J., Hassan M.Q., Murray W.J.: Effects of BHA, d-alpha-tocopherol and retinol acetate on TCDD-mediated changes in lipid peroxidation, glutathione peroxidase activity and survival. Xenobiotica, 1984; 14: 533-537
[PubMed]
[119] Tanaka G., Kanaji S., Hirano A., Arima K., Shinagawa A., Goda C., Yasunaga S., Ikizawa K., Yanagihara Y., Kubo M., Kuriyama-Fujii Y., Sugita Y., Inokuchi A., Izuhara K.: Induction and activation of the aryl hydrocarbon receptor by IL-4 in B cells. Int. Immunol., 2005; 17: 797-805
[PubMed] [Full Text HTML] [Full Text PDF]
[120] Tang K.M., Nace C.G.Jr., Lynes C.L., Maddaloni M.A., LaPosta D., Callahan K.C.: Characterization of background concentrations in upper Manhattan, New York apartments for select contaminants identified in World Trade Center dust. Environ. Sci. Technol., 2004; 38: 6482-6490
[PubMed]
[121] Tang K.M., Nace C.G.Jr., Maddaloni M.A., Harkay J.D., Graham D.R.Jr., Koustas R.N., Rotola J.D., LaPosta D., Callahan K.C.: Evaluation of vacuum and wet-wipe methods for removal of World Trade Center dust from indoor environments. J. Air Waste Manag. Assoc., 2004; 54: 1293-1298
[PubMed]
[122] Tian Y., Ke S., Thomas T., Meeker R.J., Gallo M.A.: Transcriptional suppression of estrogen receptor gene expression by 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD). J. Steroid Biochem. Molec. Biol., 1998; 67: 17-24
[PubMed]
[123] Tracey W.R., Nakane M., Kuk J., Budzik G., Klinghofer V., Harris R., Carter G.: The nitric oxide synthase inhibitor, l-N-monomethylarginine, reduces carrageenan-induced pleurisy in the rat. J. Pharmacol. Exp. Ther., 1995; 273: 1295-1299
[PubMed]
[124] Trickler D., Shklar G.: Prevention by vitamin E of experimental carcinogenesis. J. Natl. Cancer Inst., 1987; 78: 165-169
[PubMed]
[125] Tsukamoto H., Rippe R., Niemela O., Lin M.: Roles of oxidative stress in activation of Kupffer and Ito cells in liver fibrogenesis. J. Gastroenterol. Hepatol., 1995; 10: S50-S53
[PubMed]
[126] Tulloch J.A., Sood M.K.: Vitamin E deficiency in Uganda. Am. J. Clin. Nutr., 1967; 20: 884-887
[PubMed] [Full Text PDF]
[127] Turnbull A.V, Rivier C.L.: Regulation of the hypothalamic-pituitary-adrenal axis by cytokines: actions and mechanisms of action. Physiol. Rev., 1999; 79: 1-71
[PubMed] [Full Text HTML] [Full Text PDF]
[128] Urbanowicz W.: Rola wątroby w odpowiedzi ostrej fazy. Post. Nauk. Med. 2000, 13, 46-49
[129] Utsunomiya I., Nagai S., Oh-ishi S.: Sequential appearance of IL-1 and IL-6 activities in rat carrageenin-induced pleurisy. J. Immunol., 1991; 147: 1803-1809
[PubMed]
[130] Verreault J., Norstrom R.J., Ramsay M.A., Mulvihill M., Letcher R.J.: Composition of chlorinated hydrocarbon contaminants among major adipose tissue depots of polar bears (Ursus maritimus) from the Canadian high Arctic. Sci. Total. Environ., 2006; 370: 580-587
[PubMed]
[131] Viluksela M., Bager Y., Tuomisto J.T., Scheu G., Unkila M., Pohjanvirta R., Flodstrom S., Kosma V.M., Maki-Paakkanen J., Vartiainen T., Klimm C., Schramm K.W., Warngard L., Tuomisto J.: Liver tumor-promoting activity of 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD) in TCDD-sensitive and TCDD-resistant rat strains. Cancer Res., 2000; 60: 6911-6920
[PubMed] [Full Text HTML] [Full Text PDF]
[132] Vogel C., Schuhmacher U.S., Degen G.H., Bolt H.M., Pineau T., Abel J.: Modulation of prostaglandin H synthase-2 mRNA expression by 2,3,7,8-tetrachlorodibenzo-p-dioxin in mice. Arch. Biochem. Biophys., 1998; 351: 265-271
[PubMed]
[133] Vos J.G., Moore J.A., Zinkl J.G.: Effect of 2,3,7,8-tetrachlorodibenzo-p-dioxin on the immune system of laboratory animals. Environ. Health. Perspect., 1973; 5: 149-162
[PubMed] [Full Text HTML] [Full Text PDF]
[134] Wang S., Sun N.N., Zhang J., Watson R.R., Witten M.L.: Immunomodulatory effects of high-dose alpha-tocopherol acetate on mice subjected to sidestream cigarette smoke. Toxicology, 2002; 175: 235-245
[PubMed]
[135] Weglicki W.B., Stafford R.E., Freedman A.M., Cassidy M.M., Phillips T.M.: Modulation of cytokines and myocardial lesions by vitamin E and chloroquine in a Mg-deficient rat model. Am. J. Physiol., 1993; 264: C723-C726
[PubMed]
[136] Weiser H.: Die Wirkungen des Vitamins E auf den Haemolysevorgang. W: von Kress H. K.U. Blum: Vitamine A, E und K, klinische und physiologisch-chemische Probleme. Stuttgart- New York: Schattauer F. K. – Verlag 1969; 309-319
[137] Wrbitzky R., Beyer B., Thoma H., Flatau B., Hennig M., Weber A., Angerer J., Lehnert G.: Internal exposure to polychlorinated dibenzo-p-dioxins and polychlorinated dibenzofurans (PCDDs/PCDFs) of Bavarian chimney sweeps. Arch. Environ. Contim. Toxicol., 2001; 40: 136-140
[PubMed]
[138] Yan L., Wang S., Rafferty S.P., Wesley R.A., Danner R.L.: Endogenously produced nitric oxide increases tumor necrosis factor-α production in transfected human U937 cells. Blood, 1997; 90: 1160-1167
[PubMed] [Full Text HTML] [Full Text PDF]
[139] Yoon C.Y., Park M., Kim B.H., Park J.Y., Park M.S., Jeong Y.K., Kwon H., Jung H.K., Kang H., Lee Y.S., Lee B.J.: Gene expression profile by 2,3,7,8-tetrachlorodibenzo-p-dioxin in the liver of wild-type (AhR+/+) and aryl hydrocarbon receptor-deficient (AhR-/-) mice. J. Vet. Med. Sci., 2006; 68: 663-668
[PubMed] [Full Text PDF]
[140] Zeman K.: Współczesne poglądy na rolę granulocytów obojętnochłonnych (neutrofilów) w procesach zapalnych. Patofizjologiczne podstawy udziału neutrofilów w zapaleniu. Immunol. Pol., 1993; 18: 3-21
Autorzy deklarują brak potencjalnych konfliktów interesów.