
Proteins of human milk involved in immunological processes
Jolanta Lis 1 , Magdalena Orczyk-Pawiłowicz 1 , Iwona Kątnik-Prastowska 1Streszczenie
Mleko matki zawiera wiele cząsteczek (m.in. białka, węglowodany, lipidy, związki nieorganiczne), które dostarczają noworodkom podstawowych składników budulcowych w pierwszym okresie ich życia. Skład jakościowy związków obecnych w mleku jest podobny u każdej zdrowej matki, jednak ich stężenie zmienia się wraz z okresami laktacji. W pierwszych dniach po porodzie przez komórki nabłonkowe gruczołu sutkowego wytwarzana jest siara (colostrum), między 5-15 dniem laktacji mleko niedojrzałe, a powyżej 15 dnia mleko dojrzałe. Naturalne mleko ludzkie, oprócz składników odżywczych, jest również źródłem cząsteczek biologicznie aktywnych, wśród których znajdują się białka bezpośrednio i pośrednio uczestniczące w procesie odporności, tj. immunoglobuliny, czynniki wzrostu, cytokiny, białka ostrej fazy, białka o właściwościach przeciw- bakteryjnych oraz przeciwwirusowych. Bioaktywne cząsteczki sprawiają, że ludzkie mleko jest niezwykle specyficzną wydzieliną, która odgrywa istotną rolę w prawidłowym rozwoju noworodka. Cząsteczki o działaniu bioaktywnym obecne w mleku matki nie tylko wspomagają działanie niedojrzałego układu odpornościowego noworodka, ale także chronią go przed rozwojem infekcji. W pracy przedstawiono aktualny stan wiedzy na temat szlaków zaangażowanych w powstawanie mleka, proteomu ludzkiego mleka, zawartości niektórych składników mleka w zależności od okresu laktacji, scharakteryzowano białka uczestniczące w odporności nieswoistej i swoistej, białka uczestniczące w komunikacji międzykomórkowej, niektóre czynniki wzrostu, białka uczestniczące w immunomodulacji oraz białka zaangażowane w proces zapalny.
Słowa kluczowe:mleko ludzkie • proteom mleka ludzkiego • bioaktywne białka • immunoglobuliny • cytokiny • białka ostrej fazy
Summary
Human milk contains a lot of components (i.e. proteins, carbohydrates, lipids, inorganic elements) which provide basic nutrients for infants during the first period of their lives. Qualitative composition of milk components of healthy mothers is similar, but their levels change during lactation stages. Colostrum is the fluid secreted during the first days postpartum by mammary epithelial cells. Colostrum is replaced by transitional milk during 5-15 days postpartum and from 15 days postpartum mature milk is produced. Human milk, apart from nutritional components, is a source of biologically active molecules, i.e. immunoglobulins, growth factors, cytokines, acute phase proteins, antiviral and antibacterial proteins. Such components of human milk are responsible for specific biological activities of human milk. This secretion plays an important role in growth and development of newborns. Bioactive molecules present in the milk support the immature immune system of the newborn and also protect against the development of infection. In this article we describe the pathways involved in the production and secretion of human milk, the state of knowledge on the proteome of human milk, and the contents of components of milk during lactation. Moreover, some growth factors and proteins involved in innate and specific immunity, intercellular communication, immunomodulation, and inflammatory processes have been characterized.
Key words:human milk • human milk proteome • bioactive proteins • immunoglobulins • cytokines • acute phase proteins
Wykaz skrótów:
AGP – α1-kwaśna glikoproteina (α1-acid glycoprotein); BDNF – mózgowy czynnik neurotroficzny (brain-derived neurotrophic factor); CRP – białko C- reaktywne (C-reactive protein); EGF – czynnik wzrostu naskórka (epidermal growth factor); FGF-7 – czynnik wzrostu fibroblastów (FGF-7 – fibroblast growth factor 7); G-CSF – czynnik stymulujący tworzenie kolonii granulocytów (granulocyte-colony stimulating factor); GDNF – czynnik neurotroficzny pochodzący z linii komórek gleju (glial cell-line derived neurotrophic factor); GM-CSF – czynnik stymulujący tworzenie kolonii granulocytów i makrofagów (granulocyte-macrophage colony-stimulating factor); IFN-β – interferon β (interferon β); IFN-γ – interferon γ(interferon γ); IGF – insulinopodobny czynnik wzrostu (insulin-like growth factor); IL – interleukina (interleukin); LF – laktoferyna (laktoferrin); MBP – białko wiążące mannozę (mannose bindinig protein); NRP-1 – neuropilina 1 (neuropilin-1); pIgR – receptor dla polimerycznych form immunoglobulin (polymeric immunoglobulin receptor); S-IgA – wydzielni-cze IgA (secretory IgA); TGF-β – transformujący czynnik wzrostu β (transforming growth factor β); TNF-α- czynnik martwicy nowotworu α (tumor necrosis factor α); TNF-RI – receptor 1 TNF (TNF I receptor); VCAM – cząsteczka adhezji komórkowej naczyń (vascular cell adhesion molecule); VEGF – czynnik wzrostu śródbłonka naczyń (vascular endothelial growth factor).
Wstęp
Mleko ssaków, poza składnikami odżywczymi, zawiera wiele innych cząsteczek, które mogą pełnić różnorodne funkcje biologiczne, m.in. ułatwiają trawienie oraz przyswajanie składników odżywczych zawartych w mleku, biorą udział w procesach odpornościowych, stymulują prawidłowy rozwój dziecka, chronią przed infekcjami wywoływanymi przez wirusy oraz bakterie, uczestniczą w procesie dojrzewania przewodu pokarmowego, a w przypadku zachorowania wspomagają powrót do zdrowia [41,42,87,96,97,98,110].
Zgodnie z zaleceniami Amerykańskiej Akademii Pediatrii [4] oraz Europejskiego Stowarzyszenia Neonatologii i Perinatologii [117] kobiety, które nie mogą karmić naturalnie, jako mleko drugiego wyboru powinny stosować mleko pochodzące z banku mleka. Ze względu na zawartość bioaktywnych składników, karmienie noworodków (w tym wcześniaków) oraz niemowląt mlekiem naturalnym efektywnie wspomaga działanie niedojrzałego układu odpornościowego dziecka, chroniąc przed patogenami kolonizującymi przewód pokarmowy oraz układ oddechowy, zmniejszając częstość występowania infekcji dróg oddechowych, sepsy, martwiczego zapalenia jelita oraz biegunek [59,69].
W opracowaniu przedstawiono aktualny stan wiedzy o białkach obecnych w mleku pod kątem ich bezpośredniego i pośredniego zaangażowania w procesy immunologiczne, z pominięciem roli bioaktywnych peptydów mleka, które doczekały się licznych opracowań przez innych badaczy [35,57,86].
Szlaki metaboliczne zaangażowane w powstawanie mleka
Mleko jest wydzieliną powstającą w wyniku miejscowej syntezy przez komórki nabłonkowe gruczołu sutkowego oraz przesączem rozpuszczalnych składników krwi i płynu śródmiąższowego, które zawiera wiele białek o różnorodnych funkcjach. W powstawanie mleka ludzkiego zaangażowanych jest kilka szlaków, dzięki czemu możliwe jest uzyskanie niezwykle złożonej pod względem składu wydzieliny. Cztery, spośród pięciu szlaków, angażują komórki nabłonkowe gruczołu sutkowego i obejmują: syntezę białek, ich transport wewnątrz komórek nabłonkowych, a następnie egzocytozę (I), powstawanie i wydzielanie lipidów (II), transport przez błonę (III) oraz transcytozę (IV). Piąta z kolei ścieżka jest szlakiem okołokomórkowym (ryc.1) [81].

Ryc. 1. Schemat procesu wytwarzania i wydzielania mleka przez komórki nabłonkowe czynnego gruczołu sutkowego. Liczbami rzymskimi przedstawiono poszczególne szlaki zaangażowane w proces wytwarzania mleka: I – egzocytoza, II – wydzielanie trójacylogliceroli, III – transport związków niskocząsteczkowych , IV – transcytoza, V – szlak okołokomórkowy; BP – błona podstawna, RE – retikulum endoplazmatyczne, PW – pęcherzyk wydzielniczy. Rycina własna na podstawie [80]
W wyniku egzocytozy wydzielana jest większość składników mleka. Na rybosomach komórek pęcherzykowych gruczołu sutkowego (alveolar cell from lactating mammary gland) są syntetyzowane białka, które następnie przenoszone są do retikulum endoplazmatycznego, stąd do aparatu Golgiego, a po zapakowaniu do pęcherzyków wydzielniczych są wydzielane poza komórkę (ryc.1, droga I). W ten sposób uwalniane są: m.in. laktoferyna, α-laktoalbumina, kwaśne białko serwatki oraz kazeina, która do mleka transportowana jest w postaci miceli kazeinowych zawierających jony wapnia [18]. Pęcherzyki wydzielnicze są również źródłem cytrynianu wapnia, fosforanu oraz glukozy.
Wydzielanie trójacylogliceroli do mleka odbywa się poprzez szczytowe części komórek wydzielniczych (ryc.1, droga II). Trójacyloglicerole, powstałe z kwasów tłuszczowych i glicerolu w retikulum endoplazmatycznym, łączą się tworząc kuleczki tłuszczu, które po zbliżeniu się do szczytowych części komórki są wydzielane do mleka razem z otaczającym je fragmentem błony komórki gruczołu sutkowego. Błona otaczająca wydzielone kuleczki tłuszczu jest źródłem fosfolipidów oraz cholesterolu, a także zapobiega łączeniu się poszczególnych kuleczek ze sobą.
W szczytowych fragmentach komórek wydzielniczych odbywa się transport przez błonę małych cząsteczek, takich jak woda, kationy sodowe i potasowe oraz chlorki i niektóre monosacharydy (ryc.1, droga III).
W procesie tzw. transcytozy wytwarzane są m.in. wydzielnicze IgA (S-IgA – secretory IgA). W procesie tym zsyntetyzowane przez plazmocyty IgA są wiązane przez odpowiednie receptory na jednym biegunie komórek nabłonkowych, a następnie w postaci pęcherzyków wewnątrzcytoplazmatycznych transportowane są na ich drugi koniec. Po połączeniu się pęcherzyka z błoną komórki transportowane S-IgA są uwalniane do mleka (ryc. 1, droga IV). W podobny sposób mogą z osocza do mleka przedostawać się inne białka, hormony oraz czynniki wzrostu.
Szlak okołokomórkowy umożliwia bezpośrednią wymianę składników między przestrzenią śródmiąższową, osoczem, a mlekiem. Droga ta odgrywa istotną rolę w pierwszych dniach po porodzie, bowiem wówczas do mleka w ten sposób mogą przedostawać się niektóre składniki osoczowe, głównie albumina oraz immunoglobuliny. W 2-3 tygodniu laktacji dochodzi do uszczelnienia przestrzeni międzykomórkowej, co prowadzi do zamknięcia szlaku (ryc.1, droga V) [81].
Rodzaje mleka w zależności od okresu laktacji
Zapotrzebowanie noworodków na różnego rodzaju składniki w pierwszym miesiącu ich życia ulega zmianom. Jednocześnie zmienia się skład mleka matki. Bezpośrednio po porodzie wytwarzana jest siara (colostrum), w okresie 5-15 dni po porodzie mleko niedojrzałe, a powyżej 15 dnia laktacji mleko dojrzałe [9]. Światowa Organizacja Zdrowia [19] podaje, że siara wydzielana jest 0-5 dnia po porodzie, mleko niedojrzałe 6-14 dnia, a mleko dojrzałe między 15-30 dniem laktacji. Przejście siary w mleko niedojrzałe, a następnie w mleko dojrzałe u każdej kobiety jest procesem dość indywidualnym, stąd drobne rozbieżności w określeniu przez badaczy czasu powstawania mleka niedojrzałego i dojrzałego, niemniej w każdym przypadku, stężenia podstawowych składników zawartych w mleku pozostają na charakterystycznym dla danego etapu laktacji poziomie. Wydzieliny pochodzące z odmiennych okresów laktacji różnią się wartością energetyczną oraz zawartością składników bioaktywnych [9,11,30,43,88,122]. Siara jest mniej kaloryczna, zawiera więcej białka oraz mniej sacharydów niż dojrzałe mleko. W mleku niedojrzałym z kolei jest więcej białka, a mniej tłuszczu i laktozy niż w mleku dojrzałym (tab. 1) [37].
Tabela 1. Zawartość głównych składników mleka ludzkiego w zależności od okresu laktacji
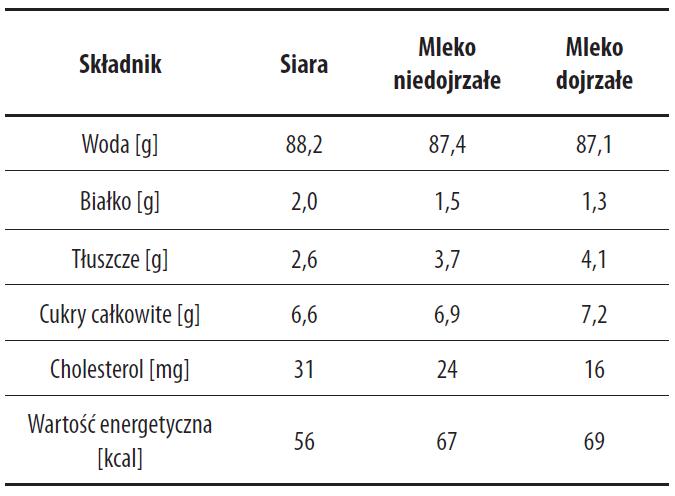
Dane w g na 100 g suchej masy wg [36].
Porównanie zawartości wybranych składników mleka ludzkiego i krowiego
Białka mleka ludzkiego, podobnie jak białka mleka krowiego, dzieli się na białka kazeinowe (białka nierozpuszczalne) oraz białka serwatkowe (białka obecne w mleku w postaci rozpuszczalnej) [111,124]. Proporcje białek kazeinowych do serwatkowych w mleku krów oraz ludzi znacznie się różnią. Odmienne jest też zapotrzebowanie noworodków i cieląt na składniki nieorganiczne i organiczne. Według van Neerven i wsp. [124] mleko krowie zawiera aż 26,4 g/L białek kazeinowych oraz jedynie 5,24 g/L białek serwatkowych. W dojrzałym mleku ludzkim całkowite stężenie kazein wynosi 7 g/L, z kolei białek serwatkowych 7,25 g/L [123]. Proporcja między zawartością białek kazeinowych i białek serwatkowych w mleku ludzkim ulega zmianie w czasie laktacji. W siarze stosunek białek kazeinowych do białek serwatkowych wynosi 20:80, następnie w mleku przejściowym 40:60, a w mleku dojrzałym 50:50 [74]. Istotne różnice obserwowane są także między zawartością tłuszczów, cukrów, witamin oraz składników mineralnych. Mleko ludzkie w porównaniu do mleka krowiego zawiera więcej α-laktoalbuminy, laktoferyny, lizozymu, przeciwciał klasy IgA, laktozy, wolnych oligosacharydów, kwasów tłuszczowych, a także witamin A oraz C. Ponadto mleko ludzkie nie zawiera laktoglobuliny β i kazeiny αS1 (tabela 2).
Tabela 2. Porównanie zawartości niektórych składników w dojrzałym mleku ludzkim i krowim

Na podstawie [123]
Charakterystyka białek mleka ludzkiego
Mleko ludzkie było i jest przedmiotem badań mających na celu szczegółowe ustalenie profilu białkowego [28,43,44,72,88]. D’Alessandro i wsp. [28] dokonali podsumowania analiz proteomicznych mleka wykonanych do 2010 r. i podali, że spośród 285 zidentyfikowanych białek mleka ~67% znajdowało się w przestrzeni międzykomórkowej. W przestrzeni tej obecne były nie tylko białka będące głównie źródłem składników odżywczych (białka kazeinowe, α-laktoalbumina), ale także białka pełniące funkcje biologiczne. Białka mleka zostały podzielone w zależności od pełnionej funkcji. Zidentyfikowano m.in. białka uczestniczące w:
• transporcie składników odżywczych oraz w metabolizmie lipidów, np.: α-albumina (transport witaminy E do mózgu), albumina (transport lipidów i związków niskocząsteczkowych), apolipoproteiny A-I, A-IV, C-I, E oraz białka błonowe uczestniczące w wiązaniu i transporcie lipidów;
• stymulacji proliferacji i różnicowania limfocytów, granulocytów, makrofagów, np.: czynnik wzrostu fibroblastów (FGF-7- fibroblast growth factor 7), erytropoetyna
• procesie angiogenezy np.: neuropilina 1 (NRP-1 – neuropilin 1), czynnik wzrostu śródbłonka naczyń (VEGF);
• odpowiedzi immunologicznej np.: immunoglobuliny, interleukiny, α-laktoalbumina (źródło peptydów o właściwościach przeciwbakteryjnych), czynnik stymulujący tworzenie kolonii makrofagów (M-CSF – macrophage colony-stimulating factor);
• dojrzewaniu układu nerwowego np.: interleukina 6 (IL- 6), interferon β (IFN-β) oraz mózgowy czynnik neurotroficzny (BDNF);
• adhezji i przekazywaniu sygnałów między limfocytami a śródbłonkiem w tym: cząsteczka adhezji komórkowej naczyń (VCAM).
Molinari i wsp. [88] dwa lata później, dzięki zaawansowanej analizie proteomicznej (połączenie technik elektroforetycznych, chromatografii cieczowej, programu ProteinPilot oraz bazy danych Swiss-Prot), zidentyfikowali 415 białek obecnych w odtłuszczonym mleku między 7-28 dniem laktacji. Spośród wszystkich zidentyfikowanych białek, aż 261 wykryto po raz pierwszy [88]. Zidentyfikowane białka podzielono pod względem pełnionych funkcji biologicznych (ryc. 2A) oraz wewnątrz- lub zewnątrzkomórkowej lokalizacji (ryc. 2B).
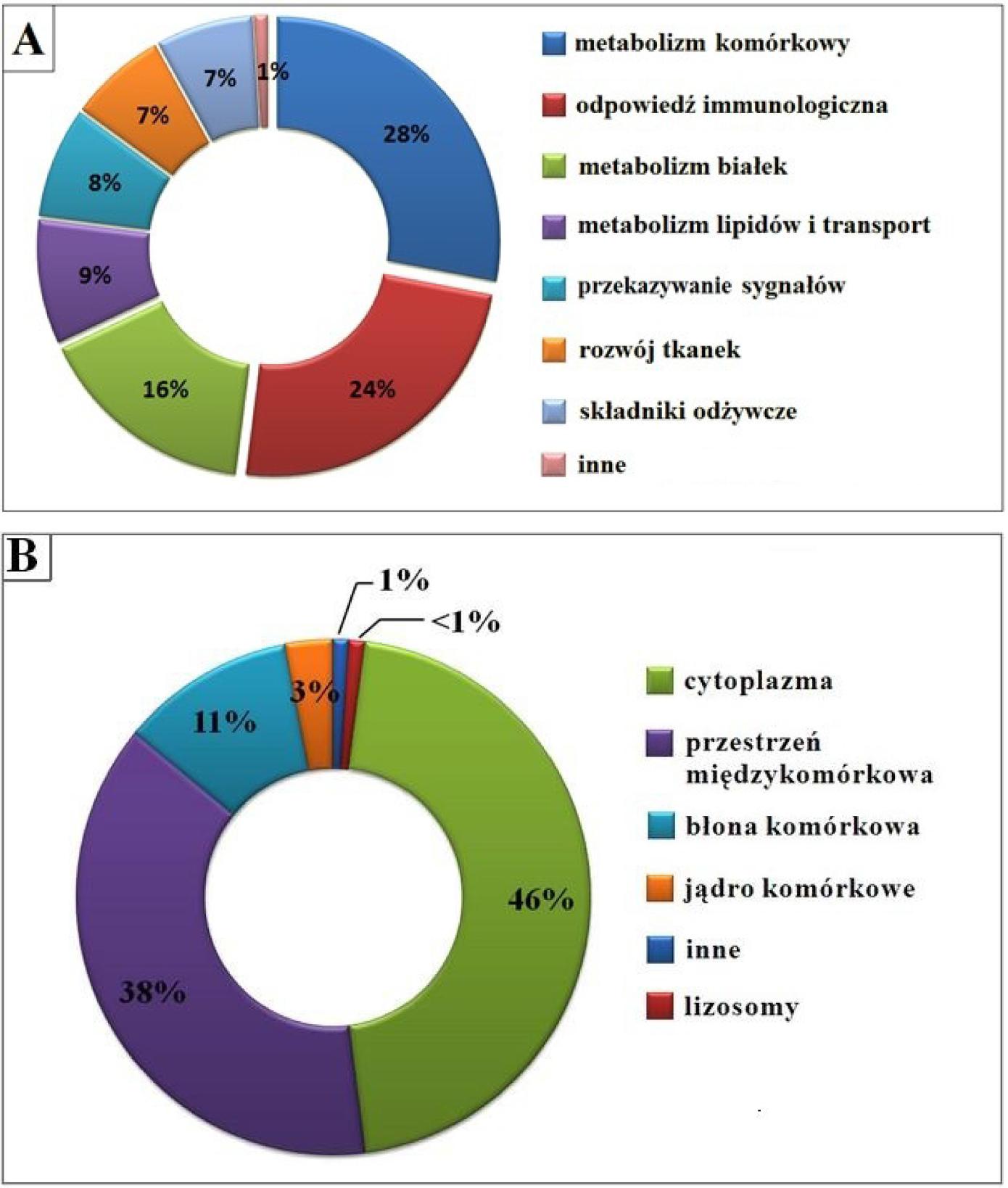
Ryc. 2. Podział białek mleka ludzkiego w zależności od pełnionej funkcji (A) oraz komórkowej lokalizacji (B). Rycina własna na podstawie [88]
Gao i wsp. [44] badając proteom mleka ludzkiego zidentyfikowali 976 białek, spośród których ekspresja 152 w sposób istotny zmieniała się w mleku niedojrzałym i dojrzałym. Dodatkowo wykazano, że immunoglobuliny S-IgA oraz IgM występują w porównaniu z IgG w wyższym stężeniu w mleku niedojrzałym. W mleku dojrzałym stężenie IgG jest wyższe niż S-IgA oraz IgM. Autorzy [44] sugerują, że zmieniające się stężenia S-IgA, IgM oraz IgG w mleku matki wpływają na zmianę mechanizmów immunologicznej ochrony dziecka podczas przejścia z etapu noworodka do niemowlęcia.
W dalszej części pracy scharakteryzowano białka kazeinowe oraz serwatkowe, do których zalicza się m.in.: białka uczestniczące w odporności swoistej oraz nieswoistej, białka uczestniczące w komunikacji międzykomórkowej, białka ostrej fazy, białka uczestniczące w regulacji równowagi energetycznej oraz peptydy o właściwościach bifidogennych.
Białka kazeinowe
Kazeiny są białkami mleka, które są syntetyzowane, a następnie wydzielane przez komórki nabłonkowe gruczołu sutkowego. U ludzi występuje: kazeina-β, kazeina-κ oraz kazeina-αs1 [76,88,111]. Kazeina jest nie tylko źródłem aminokwasów, ale zapewnia także odpowiednią ilość fosforanów oraz wapnia, niezbędnych w procesie mineralizacji kości.
Kazeina poza pełnieniem funkcji odżywczych może również pełnić funkcje immunomodulujące. Kazeina-αs1 jest m.in. źródłem peptydów o właściwościach przeciwbakteryjnych oraz immunostymulujących, które przyczyniają się do wzrostu ekspresji i wydzielania czynnika stymulującego tworzenie kolonii granulocytów i makrofagów (GM-CSF) [3,28,125].
Kazeina-β jest białkiem wysoce ufosforylowanym. Grupa fosforanowa przyłącza się do grupy hydroksylowej reszt seryny i treoniny umiejscowionych przy N-końcu łańcucha peptydowego. Region ten zdolny jest do kompleksowania jonów Ca2+. Powstające z kazeiny podczas trawienia fosfopeptydy sprzyjają poprawie wchłaniania jonów wapniowych i innych dwuwartościowych jonów (m.in. cynku) przez przewód pokarmowy dziecka.
Kazeina-κ jest sjaloglikoproteiną zdolną do hamowania adhezji bakterii do błony śluzowej żołądka. Z tego powodu karmienie dzieci naturalnym pokarmem zapewnia znaczną ochronę przed zakażeniem przez Helicobacter pylori [74], dla których ligandem są usjalowane determinanty.
Białka serwatkowe
Mleko ssaków, oprócz nierozpuszczalnych białek kazeinowych, zawiera białka serwatkowe, które są białkami globularnymi obecnymi w mleku w postaci rozpuszczonej. Wiele z białek serwatkowych jest bezpośrednio lub pośrednio zaangażowanych w procesy immunologiczne: immunoglobuliny, laktoferyna, lizozym, cytokiny, czynniki wzrostu, zapewniając noworodkom, z niedojrzałym jeszcze układem odpornościowym, efektywną ochronę przed infekcjami, a w przypadku zachorowania szybki powrót do zdrowia [74,111]. W tabeli 3 przedstawiono stężenia wybranych białek zaangażowanych w procesy odpornościowe w zależności od okresu laktacji. Karmienie noworodków mlekiem ludzkim chroni przed infekcjami i stanami zapalnymi, zwłaszcza siara, która jest bogata w czynniki zapewniające ochronę immunologiczną noworodkowi. Ogromna ilość oraz wielofunkcyjność tych składników nadal jest przedmiotem intensywnych badań.
Tabela 3. Stężenie wybranych białek mleka zaangażowanych w procesy odpornościowe u noworodków oraz niemowląt w zależności od okresu laktacji

Białka mleka uczestniczące w odporności nieswoistej
Płód w łonie matki, podczas prawidłowo przebiegającej ciąży, rozwija się w środowisku sterylnym. Dopiero podczas narodzin dziecko po raz pierwszy kontaktuje się z mikroorganizmami. Układ odpornościowy, zarówno noworodków jak i niemowląt, nie jest układem w pełni dojrzałym i w pierwszych miesiącach życia główną rolę w walce z mikroorganizmami odgrywa odporność wrodzona, zwana również nieswoistą. Do najważniejszych zidentyfikowanych białek mleka uczestniczących w reakcjach obrony nieswoistej należą: lizozym, laktoferyna, laktoperoksydaza oraz składniki dopełniacza [74,93,111].
Laktoferyna
Laktoferyna (LF) obecna jest w mleku różnych gatunków ssaków, m.in.: krów, świń, myszy oraz u człowieka [111]. Ludzka laktoferyna jest jednołańcuchowym polipeptydem, zawierającym 691 reszt aminokwasowych [61]. Stężenie laktoferyny w siarze osiąga wartość 6-8 g/L, w następnych etapach laktacji ulega obniżeniu do 2-4 g/L. Wysokie stężenie laktoferyny w siarze, a także jej obecność w mleku niedojrzałym i dojrzałym, sugeruje znaczący udział tego białka w ochronie noworodka przed patogenami, które kolonizują przewód pokarmowy noworodka [6,111].
Przeciwbakteryjne działanie laktoferyny związane jest m.in. z możliwością wiązania jonów żelaza, które jest wymagane do wzrostu niektórych bakterii i grzybów. Ponadto, laktoferyna poprzez wiązanie się do fimbrialnych adhezyn bakterii uniemożliwia adhezję patogenów do nabłonkowych komórek jelita gospodarza. Blokowanie etapu adhezji bakterii na powierzchni komórek, zapobiega przedostaniu się ich do wnętrza komórki i dalszym etapom infekcji [6]. Antywirusowe działanie laktoferyny wynika przede wszystkim z tego, że białko to wiąże się z receptorami błonowymi komórki gospodarza zbudowanymi z glikozaminoglikanów, które są miejscem interakcji z wieloma wirusami. Jak dotąd potwierdzono działanie laktoferyny m.in. przeciw wirusowi opryszczki typu 1 i 2, ludzkiemu wirusowi cytomegalii, ludzkiemu wirusowi niedoboru odporności (HIV), wirusowi brodawczaka ludzkiego (HPV), rotawirusom, enterowirusom, adenowirusom, wirusowi grypy, wirusowi paragrypy, wirusowi zapalenia wątroby typu C (HCV) oraz wirusowi zapalenia wątroby typu B (HBV) [6,70,96].
Laktoferyna, oprócz antymikrobiologicznego działania, pełni również wiele innych niezwykle istotnych dla rozwoju niemowlęcia funkcji, m.in. stymuluje wzrost tkanek jelita, chroni przed translokacją bakterii jelitowych do układu krążenia noworodka, a także chroni jelita i inne tkanki przed stresem oksydacyjnym [6]. Laktoferyna może również w sposób bezpośredni wpływać na układ odpornościowy gospodarza poprzez stymulację syntezy nieswoistych przeciwciał klas IgA i IgG (w jelicie) oraz aktywację limfocytów B, T oraz NK (w jelicie, śledzionie i krwi obwodowej) [6]. Esaguy i wsp. [38] wykazali, że laktoferyna może także wpływać na proliferację limfocytów. Jest to związane najprawdopodobniej z działaniem LF na obecne w grasicy prekursorowe komórki T, przez co nabywają one fenotypu komórek pomocniczych Th CD4+CD8– [129]. Natomiast badania Ashorna i wsp. [7] oraz Richie i wsp. [104] wykazały hamujące działanie LF w tym procesie. Mechanizm stymulacji komórek T do proliferacji może także wynikać z indukcji przez to białko kinazy indukowanej miogenem [31]. Zimecki i wsp. [130], na podstawie badań przeprowadzonych na myszach, sugerują ponadto promowanie dojrzewania limfocytów B przez laktoferynę.
Laktoferyna może ponadto obniżać wytwarzanie niektórych cytokin prozapalnych, takich jak czynnik martwicy nowotworu α (TNF-α), IL-1α i IL-6 [52,70,78] oraz zwiększać stężenia przeciwzapalnej IL-10 [2].
Laktoperoksydaza
Laktoperoksydaza obecna jest w wydzielinach wytwarzanych przez gruczoły sutkowe oraz ślinowe. W siarze kobiet aktywność laktoperoksydazy jest stosunkowa niska (~5%) w porównaniu do siary krów. Białko obecne jest również w ślinie niemowląt w stężeniu, które w warunkach in vitro skutecznie hamuje wzrost mikroorganizmów [50]. Laktoperoksydaza jest enzymem, który w obecności nadtlenku wodoru katalizuje reakcję utlenienia tiocyjanków (SCN) do związków o właściwościach przeciwbakteryjnych. Szczególnie aktywne działanie bakteriostatyczne laktoperoksydaza wykazuje wobec paciorkowców [97].
Alfa-laktoalbumina
Alfa-laktoalbumina jest hydrofilowym białkiem zbudowanym z 123 aminokwasów, które u większości ssaków, w tym również u człowieka, wykazuje wysoki stopień homologii. W organizmie człowieka α-laktoalbumina, poza funkcją odżywczą, pełni również inne funkcje m.in. uczestniczy w biosyntezie laktozy, ułatwia (przez wiązanie jonów wapnia oraz cynku) wchłanianie niezbędnych minerałów oraz jest źródłem aminokwasów, a także peptydów o właściwościach przeciwbakteryjnych [76,107]. Możliwość wiązania jonów dwuwartościowych przez α-laktoalbuminę może również pośrednio wpływać na system odpornościowy noworodków oraz niemowląt, bowiem białko to, wiążąc jony wapnia, obniża ich poziom wymagany do pełnej aktywności składników układu dopełniacza [94].
Lizozym
Lizozym jest enzymem o masie cząsteczkowej 15 kDa, który powszechnie występuje w tkankach, narządach, osoczu i innych płynach ustrojowych, a także w wydzielinach ludzkiego organizmu, takich jak ślina, mleko, łzy, wydzieliny surowiczo-śluzowe. Stężenie lizozymu w ludzkim mleku w pierwszych 3 tygodniach laktacji najwyższe jest w siarze (0,37 g/L). W kolejnych dniach laktacji stężenie lizozymu maleje, a około 28 dnia zaczyna rosnąć, osiągając w 57-84 dniu wartość 0,89 g/L [89,97]. Według Reddy’ego i wsp. [102] i Goldmana [48] w mleku kobiet, które urodziły dziecko przed terminem, stężenie lizozymu osiąga wyższą wartość w porównaniu ze stężeniem lizozymu w siarze kobiet, które urodziły w terminie.
Lizozym katalizuje reakcję hydrolizy wiązania β1,4- glikozydowego między kwasem N-acetylomuraminowym a N-acetyloglukozoaminą, co prowadzi do uszkodzenia ściany komórkowej bakterii Gram-dodatnich i niektórych bakterii Gram-ujemnych, dzięki czemu wykazuje działanie bakteriostatyczne, zmniejszając kolonizację przewodu pokarmowego noworodków przez bakterie [97,111].
Lizozym wykazuje działanie wspomagające podczas odpowiedzi immunologicznej (adjuvant effect of lysozyme). W wyniku hydrolizy peptydoglikanu ściany komórkowej bakterii z rodzaju Mycobacterium przez lizozym uwalniany jest tzw. dipeptyd muramylowy. Dipeptyd ten wzmacnia efekt antyinfekcyjny powodując m.in. aktywację komórek prezentujących antygen oraz zwiększenie proliferacji limfocytów T [25,51,97].
Białka układu dopełniacza oraz jego inhibitory
W immunologicznej ochronie noworodków i niemowląt szczególnie istotną rolę może odgrywać droga alternatywna aktywacji układu dopełniacza. Droga ta uruchamiana jest niemal natychmiast po wniknięciu drobnoustroju do organizmu i w przeciwieństwie do drogi klasycznej nie jest konieczna biosynteza przeciwciał [66,80,94].
Noworodki przychodzące na świat między 38 a 42 tygodniem ciąży mają słabo rozwinięty układ dopełniacza. Związane jest to przede wszystkim z nieobecnością w dostatecznym stężeniu niektórych składowych układu dopełniacza. Jeszcze niższe stężenie niektórych składników dopełniacza obserwowane jest w przypadku noworodków urodzonych przed terminem, dlatego też wcześniaki są szczególnie podatne na infekcje spowodowane przez mikroorganizmy [80].
Mleko ludzkie zawiera niektóre białka układu dopełniacza, które efektywnie wspomagają układ odpornościowy noworodka, chroniąc go tym samym przed patogenami kolonizującymi jego przewód pokarmowy oraz układ oddechowy [59,69]. Dotąd w mleku ludzkim wykryto następujące białka układu dopełniacza: C2, C3, C4a, C4b, C5, C9 oraz czynnik B [1,87,90].
Morceli i wsp. [90] wykazali, że stężenie składnika C3 w mleku kobiet z hiperglikemią jest wyższe niż w mleku kobiet z prawidłowym stężeniem glukozy we krwi. Warto podkreślić, że wartości stężeń składnika C3 w surowicy kobiet zdrowych oraz z hiperglikemią były zbliżone. Z kolei stężenie składnika C4 w surowicy kobiet z hiperglikemią było wyższe w porównaniu z surowicą kobiet zdrowych, podczas gdy stężenia tego białka w mleku obu grup kobiet były porównywalne (tab. 4). Trégoat i wsp. [120] analizując mleko pochodzące od kobiet, które urodziły przed terminem wykazali obecność składników C3 oraz C4 układu dopełniacza w stężeniach wyższych niż w mleku pochodzącym od kobiet, które urodziły o czasie.
Tabela 4. Stężenie składników dopełniacza C3 oraz C4 w surowicy oraz mleku matek z prawidłowym stężeniem glukozy oraz z hiperglikemią

Dane wg [89]
Ogundele [93,94] zidentyfikował w ludzkim mleku białka pełniące funkcje inhibitorów układu dopełniacza. Autor [94] uważa, że mniejsza aktywność składnika C3 w mleku dojrzałym jest następstwem wysokiego poziomu inhibitorów układu dopełniacza w tym mleku. Jako związki mogące pełnić rolę inhibitorów układu dopełniacza, zostały wskazane: laktoferyna, lizozym, α-laktoalbumina, rozpuszczalne białko CD59 (tab. 5). Hamujący wpływ niektórych z wymienionych białek na składniki dopełniacza wynika przede wszystkim z ich zdolności do wiązania jonów dwuwartościowych, co obniża ich stężenie konieczne do pełnej aktywności białek układu dopełniacza. Obecność inhibitorów w mleku matki najprawdopodobniej zapobiega indukowaniu reakcji zapalnych powstających podczas uszkodzeń niedojrzałego przewodu pokarmowego noworodka, a także zapobiega rozwojowi zapalenia w gruczole sutkowym matki [94].
Tabela 5. Wybrane inhibitory układu dopełniacza obecne w mleku ludzkim

Dane wg [93]
Białko wiążące mannozę
Białko wiążące mannozę (MBP, nazywane również lektyną wiążącą mannozę – MBL) jest cząsteczką uczestniczącą w lektynowej drodze aktywacji układu dopełniacza. MBP po związaniu liganda cukrowego (mannoza>fukoza>N-acetyloglukozoamina), obecnego na powierzchni patogenu inicjuje kaskadę reakcji prowadzącą zazwyczaj do lizy drobnoustroju [66,118].
Badania przeprowadzone przez Trégoata i wsp. [118] wykazały obecność białka wiążącego mannozę w ludzkim mleku. Stężenie MBP w mleku maleje wraz z czasem laktacji i wynosi dla siary 0,55 mg/L, mleka niedojrzałego 0,18 mg/L i dla mleka dojrzałego 0,17 mg/L. Ze względu na to, że stężenie MBP największe jest w siarze przypuszcza się, że białko to odgrywa kluczową rolę w ograniczaniu kolonizacji tkanek organizmu noworodka przez mikroorganizmy w pierwszych dobach jego życia [118].
Białka uczestniczące w odporności swoistej
Obrona nieswoista noworodków przed infekcją może się okazać niewystarczająca, szczególnie z powodu niedostatecznie rozwiniętego układu dopełniacza. Wtedy ważną rolę odgrywa obrona swoista, w której bezpośrednio biorą udział swoiste immunoglobuliny. Wartości stężeń immunoglobulin klas IgG oraz IgM w osoczu noworodka według Kardara i wsp. [62] wynoszą odpowiednio 57,35 oraz 30,5% stężeń, w jakich występują one w organizmie zdrowych dorosłych osób. Przeciwciała klasy IgG obecne w surowicy noworodków pochodzą od matki, bowiem przeciwciała te, jako jedyne dzięki obecności receptora dla IgG są transportowane przez łożysko [27,44]. Od chwili narodzin do około 6 miesiąca życia stężenie przeciwciał IgG oraz IgM maleje. Do czasu, kiedy organizm niemowlęcia nie będzie zdolny sam wytwarzać przeciwciał, ważną rolę we wspomaganiu niedojrzałego układu odpornościowego w walce z różnego rodzaju patogenami odgrywają przeciwciała oraz inne składniki, biorące udział w mechanizmach odporności swoistej dostarczane wraz z mlekiem matki.
Immunoglobuliny
Niemowlęta rodzą się z niedojrzałym systemem odpornościowym i korzystają z obecnych w mleku matki przeciwciał do obrony przed patogenami. W mleku ludzkim dominującą klasę stanowią wydzielnicze IgA (80-90% wszystkich immunoglobulin) oraz IgM i IgG [3,15]. Wydzielnicze IgA powstają podczas transportu dimerów IgA przez komórki nabłonkowe gruczołu sutkowego do przewodów wyprowadzających gruczołu z udziałem receptora polimerycznych form immunoglobulin (pIgR). Stężenie S-IgA wynosi około 12 g/L w początkowym okresie laktacji, w etapach późniejszych ulega blisko dziesięciokrotnemu zmniejszeniu [74]. Stężenie pozostałych immunoglobulin w mleku wynosi: ~ 0,6 g/L dla IgM i ~ 0,1 g/L dla IgG. Chirico i wsp. [24] wykazali, że stężenie przeciwciał IgA w mleku matek, które urodziły przedwcześnie jest wyższe w porównaniu do mleka kobiet, które urodziły w terminie. Ponadto, Duchén i Björkstén [33] wykazali obecność IgE w 34 z 39 analizowanych próbkach siary kobiet zdrowych i z atopią.
Wydzielnicze IgA w mleku są swoiste wobec mikroflory układu pokarmowego oraz dróg oddechowych matki. Podstawowe funkcje biologiczne S-IgA obejmują: (1) neutralizację wirusów, w tym neutralizację wewnątrzkomórkową oraz hamowanie ich transcytozy, (2) hamowanie adhezji bakterii patogennych z gatunków Escherichia coli, Streptococcus pneumoniae, Haemophilus influenzae, Clostridium difficile do błon śluzowych gospodarza, (3) aglutynację wirusów i bakterii. Kompleksy S-IgA-antygen bakteryjny są rozpoznawane i przetwarzane przez komórki dendrytyczne jelita, prowadząc do usunięcia antygenu [49,60]. Ponadto, IgA dzięki obecności oligosacharydów stanowią istotny łącznik między odpornością nabytą i wrodzoną. Cząsteczki S-IgA zawierają N-glikany, a w przypadku podklasy IgA1, także O-glikany, których glikotopy są rozpoznawane i wiązane przez bakteryjne adhezyny, których interakcja zapoczątkowuje drogę lektynową aktywacji dopełniacza [24,27].
Franca i wsp. [42] porównywali stężenie immunoglobulin w siarze, osoczu matki oraz krwi pępowinowej u matek zdrowych i z hiperglikemią. Badania wykazały, że w siarze matek z hiperglikemią stężenie przeciwciał klas IgG oraz IgA było niższe w porównaniu do siary matek z prawidłowym stężeniem glukozy, natomiast stężenie IgM było porównywalne w obu grupach (tab. 6). Stężenie IgG, IgA i IgM w osoczu matek z hiperglikemią było niższe w porównaniu do grupy kontrolnej. We krwi pępowinowej stężenie IgG było wyższe u matek z hiperglikemią, natomiast wartości stężeń IgA i IgM były porównywalne. Autorzy [42] sugerują, że obniżona synteza przeciwciał u matek z hiperglikemią, może być następstwem zachodzących zmian w metabolizmie węglowodanów, lipidów oraz białek spowodowanych podwyższonym stężeniem glukozy. Ponadto, wyższe stężenie glukozy w osoczu ciężarnych może ułatwiać transfer IgG przez łożysko i wpływać na obserwowany wzrost stężenia IgG w krwi pępowinowej.
Tabela 6. Stężenie IgA, IgG oraz IgM w siarze, krwi pępowinowej oraz osoczu matek z prawidłowym stężeniem glukozy oraz z hiperglikemią
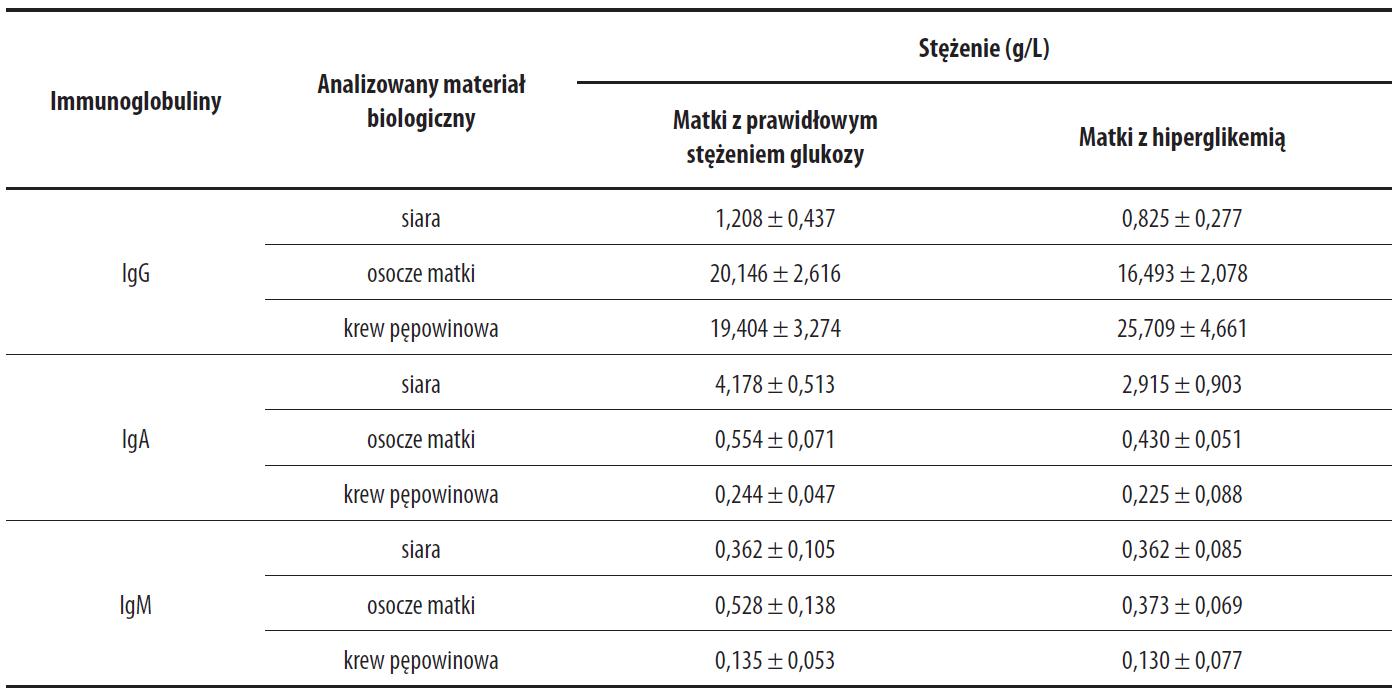
Dane wg [42]
Białka uczestniczące w komunikacji między komórkami
Cytokiny
Cytokiny obecne w mleku ludzkim, określane mianem lokalnych hormonów układu odpornościowego działającymi w sposób autokrynny i parakrynny, odgrywają znaczącą rolę w procesach immunomodulacji i immunoochrony (są mediatorami reakcji zapalnych i immunologicznych). Większość cytokin, które występują w niskim stężeniu w osoczu noworodków, w szczególności wcześniaków, występują w mleku ludzkim w znaczących ilościach. Należą do nich interleukiny: IL-1β, IL-2, -6, -8, -10, -12, -18, IFN-γ, TNFα, TGF-β oraz cytokiny hematopoetyczne. Cytokiny mleka ludzkiego mogą przekraczać barierę jelitową i wpływać na aktywność układu odpornościowego noworodka [3,24,45,68,91,123].
Najliczniejszą rodzinę cytokin mleka ludzkiego stanowi grupa TGF-β, która składa się z trzech izoform, wśród których dominuje TGF-β2 [99]. Transformujący czynnik wzrostu β jest zaliczany do przeciwzapalnych cytokin, wytwarzanych przez regulatorowe komórki B i T oraz komórki nabłonka gruczołu sutkowego. TGF-β hamuje wzrost różnych komórek, w tym nabłonka, śródbłonka oraz komórek hematopoetycznych [63]. Obecny w mleku TGF-β reguluje procesy zapalne, gojenie ran oraz bierze udział w zapobieganiu rozwoju chorób o podłożu alergicznym. TGF-β występuje mleku w postaci nieaktywnej, która ulega aktywacji w obecności niskiego pH w żołądku noworodka [91].
Cytokiny prozapalne: TNF-α, IL-6, IL-8 i IFN-γ są obecne w mleku ludzkim w stosunkowo niskim stężeniu. Poziom tych cytokin zależy od tygodnia porodu i ulega obniżeniu w czasie laktacji [123]. Dodatkowo, rozpuszczalne receptory dla TNF, które neutralizują TNF-α, są również obecne w mleku, tak więc TNF-α występuje w mleku głównie w postaci związanej, a nie w aktywnej postaci wolnej [17]. W mleku ludzkim stężenie TNF-α koreluje pozytywnie z poziomami innych cytokin prozapalnych [83]. Rola cytokin prozapalnych obecnych w mleku ludzkim jest przedmiotem intensywnych badań. Wiadomo, że są one zaangażowane w rekrutację neutrofilów oraz wpływają na rozwój komórek jelita [79]. Podwyższony poziom IL-6 i IL-8 obserwuje się w zapaleniu sutka [55,83]. Do cytokin z grupy regulacyjnych obecnych w mleku ludzkim zaliczamy także interleukiny IL-10 i IL-7. Interleukina 7, przechodzi przez ścianę jelita noworodka i wpływa na rozwój grasicy [8].
Czynnik stymulujący tworzenie kolonii granulocytów (G-CSF), jest zaliczany do cytokin hematopoetycznych wpływających na procesy różnicowania komórek szlaku krwiotworzenia. G-CSF obecny w mleku ludzkim ma korzystny wpływ na rozwój jelita oraz w terapii sepsy u noworodków [47]. G-CSF działa na powierzchnię jelita noworodków, gdzie zwiększa liczbę kosmków, głębokość krypt i proliferację enterocytów [46].
Czynniki wzrostu
Mleko ludzkie zawiera liczne czynniki wzrostu, które mają wpływ na działanie układu pokarmowego, naczyniowego, nerwowego i hormonalnego noworodka [12,23,34,64,71,77,85,106].
Czynnik wzrostu naskórka (EGF) uczestniczy w dojrzewaniu układu pokarmowego i naprawie zranień jelita. EGF jest obecny w mleku ludzkim. Stężenie EGF jest najwyższe na początku laktacji, a następnie ulega obniżeniu. W siarze jest 2000-krotnie wyższe, a w mleku dojrzałym 100-krotnie wyższe niż w surowicy matki. Ponadto, stężenie jest wyższe w mleku matek, które urodziły przedwcześnie [34]. EGF jest odporny na niskie pH i enzymy trawienne obecne w żołądku noworodka, co pozwala na jego przejście przez żołądek do jelita, w którym pobudza enterocyty do zwiększonej syntezy DNA, podziału komórek, absorpcji wody i glukozy oraz syntezy białek [23]. EGF jest niezbędny w procesach dojrzewania i gojenia zranień błony śluzowej jelita. Opisano kilka mechanizmów ochronnego działania EGF w jelicie niemowląt, w tym hamowanie programowanej śmierci komórek oraz modulację zmian indukowanych przez TNF-α [64].
Czynnik neurotroficzny pochodzący z mózgu (BDNF) oraz czynnik neurotroficzny pochodzący z linii komórek gleju (GDNF) stymulują wzrost i rozwój niedojrzałego jelitowego układu nerwowego noworodka [71,106]. Czynniki BDNF oraz GDNF są obecne w mleku ludzkim do 90 dnia laktacji [40]. Obecny w mleku BDNF wpływa na prawidłową perystaltykę jelit, która często jest zaburzona u noworodków urodzonych przedwcześnie [13], a GDNF zwiększa przeżycie neuronów [40].
W mleku ludzkim obecne są dwa insulinopodobne czynniki wzrostu (IGF) IGF-1 i IGF-2, a także białka wiążące IGF oraz proteazy IGF-swoiste [12,101]. Najwyższe stężenie czynników obserwuje się na początku laktacji, a następnie ich stężenie ulega obniżeniu [85]. Z wyjątkiem białka wiążącego IGF-2, którego stężenie w mleku matek wcześniaków jest wyższe, nie ma istotnych różnic w stężeniu IGF-1 i IGF-2 w zależności od terminu porodu [12]. IGF-1 wpływa na przeżywalność enterocytów po uszkodzeniu jelita na skutek stresu oksydacyjnego [36]. Niemowlęta karmione piersią mają wyższe stężenie IGF-1 w surowicy, w porównaniu do niemowląt karmionych sztucznie. IGF-1 może być w aktywnej postaci pobierany przez jelita i transportowany do krwi. Jego funkcja nie została jeszcze dokładnie poznana, ale dojelitowe podanie IGF-1 w stężeniu fizjologicznym stymuluje erytropoezę oraz podwyższa hematokryt u noworodków szczura [67].
Czynnik wzrostu śródbłonka naczyń (VEGF) bierze udział w procesie powstawania naczyń i regulacji angiogenezy. Regulacja odbywa się przede wszystkim przez względną ekspresję VEGF i jego antagonistów. Stężenie VEGF w mleku ludzkim jest najwyższe w siarze, aczkolwiek niższe w mleku matek wcześniaków w porównaniu do mleka matek, które urodziły w terminie [77]. Uważa się, że negatywna regulacja VEGF prowadzi do nieprawidłowego unaczynienia siatkówki a w konsekwencji do retinopatii wcześniaków. Reynolds [103] oraz DiBiasie [32] sugerują, że ludzkie mleko, dzięki zawartości VEGF, może pomóc zmniejszyć skalę problemu retinopatii wcześniaków.
Osteopontyna jako białko uczestniczące w immunomodulacji
Osteopontyna jest glikoproteiną z grupy cytokin, wydzielaną m.in. przez pobudzone limfocyty T i makrofagi, która jest obecna w tkankach, osoczu, moczu, żółci oraz w mleku. W najwyższym stężeniu wynoszącym 1,5 g/L osteopontyna występuje w mleku w pierwszych dniach po porodzie, a następnie jej stężenie spada do 0,9 g/L [110]. Osteopontyna zawiera 34 fosforylowane reszty seryny oraz dwie fosfotreoniny, co nadaje temu białku silnie anionowy charakter. Przypuszcza się, że osteopontyna przez oddziaływania elektrostatyczne może uczestniczyć w transporcie białek o właściwościach przeciwbakteryjnych i immunomodulących do miejsca ich przeznaczenia. Dodatkowo, wysoce anionowy charakter osteopontyny najprawdopodobniej jest odpowiedzialny za tworzenie rozpuszczalnych kompleksów tego białka z jonami wapnia, co zapobiega krystalizacji, a następnie precypitacji wapnia w mleku. Osteopontyna może także stymulować subpopulację limfocytów Th1 poprzez indukowanie wydzielania IL-2 z jednoczesnym hamowaniem wytwarzania IL-10, co może się przyczyniać do niszczenia wewnątrzkomórkowych patogenów, głównie wirusów oraz niektórych bakterii. Stymulowanie subpopulacji limfocytów Th1 przez osteopontynę może mieć związek z chorobami o podłożu autoimmunologicznym [110].
Białka uczestniczące w procesie zapalnym
W mleku ludzkim wykazano obecność białek ostrej fazy, w tym białka C-reaktywnego, α1-antytrypsyny, α1– antychymotrypsyny, α1-kwaśnej glikoproteiny (AGP), haptoglobiny, ceruloplazminy, fibrynogenu, prokalcytoniny oraz składników dopełniacza C3 i C4 [72,75].
Prokalcytonina
Prokalcytonina, białko o masie cząsteczkowej 13 kDa kodowane przez gen CALC-1 na chromosomie 11, jest wytwarzana przez różnego rodzaju komórki organizmu człowieka w infekcjach bakteryjnych, ogólnoustrojowych zapaleniach, zaburzeniach czynności narządów oraz tkanek. Prokalcytonina jest użytecznym markerem zapalenia wykorzystywanym powszechnie w diagnostyce medycznej [20,116,127]. Z prokalcytoniny, w wyniku swoistej reakcji proteolitycznej, powstają trzy mniejsze peptydy: N-końcowy peptyd – N-prokalcytonina, kalcytonina oraz C-końcowy peptyd – katakalcyna [121].
W surowicy zdrowych osób prokalcytonina występuje zazwyczaj w stężeniu poniżej 50 ng/L, jej poziom gwałtownie wzrasta w ciągu 3-6 godzin po zakażeniu i rośnie wraz z jego rozwojem. Badania przeprowadzone przez Strucka i wsp. [116] na mleku oraz surowicy pobranych od kobiet w pierwszych 10 dniach po porodzie wykazały, że stężenie prokalcytoniny w mleku było ponad 100 razy wyższe niż w surowicy. Najwyższe stężenie prokalcytoniny było w pierwszym oraz drugim dniu laktacji i wynosiły one odpowiednio: 2,31 µg/L i 2,44 µg/L. W dziesiątym dniu laktacji, stężenie prokalcytoniny w mleku matki wynosiło 0,75 µg/L. Znacząca różnica między stężeniem prokalcytoniny w surowicy matek w porównaniu z mlekiem prawdopodobnie wynika z miejscowej syntezy przez komórki gruczołu sutkowego kobiet po porodzie. Struck i wsp. [116] uważają, że prokalcytonina w mleku kobiet karmiących może się przyczyniać do aktywacji rozwijającego się u noworodków układu odpornościowego.
Białko C-reaktywne
Białko C-reaktywne (CRP) jest jednym z białek ostrej fazy, które syntetyzowane jest w wątrobie w odpowiedzi na IL-6 uwalnianą przez makrofagi oraz adipocyty [39]. Stężenie białka C-reaktywnego w surowicy zdrowych ludzi zazwyczaj nie przekracza wartości 0,01 g/L. Wzrost stężenia białka CRP do wartości 0,01-0,04 g/L stwierdza się u kobiet będących w zaawansowanej ciąży, a także podczas łagodnego zapalenia oraz infekcji wirusowych. Zapalenie bakteryjne powoduje wzrost stężenia CRP do wartości 0,04-0,2 g/L, a ciężkie zapalenie bakteryjne oraz oparzenia powyżej 0,2 g/L [26].
Fetherston i wsp. [39] wykazali, że stężenie białka C-reaktywnego w surowicy kobiet, których dzieci przyszły na świat w wyniku cesarskiego cięcia jest wyższe niż w surowicy kobiet, które urodziły w sposób naturalny. Niskie stężenie białka C-reaktywnego stwierdzono również w mleku kobiet, których dzieci przyszły na świat w wyniku porodu wspomaganego próżnociągiem, a także w wyniku nagłego cięcia cesarskiego (ryc. 3A i 3B). Stężenie białka CRP wzrasta w płynach ustrojowych podczas infekcji i podobnie jak prokalcytonina, jest markerem diagnostycznym rozwoju zakażeń oraz sepsy. Oznaczenie stężenia CRP zarówno we krwi jak i w mleku jest przyjętym markerem diagnostycznym zapalenia, szczególnie istotnym dla kobiet karmiących, ponieważ rozwijający się proces zapalny w organizmie matki wpływa na skład produkowanego mleka [39]. W przeciwieństwie jednak do prokalcytoniny nie udokumentowano proporcjonalnego wzrostu stężenia białka C-reaktywnego w stosunku do postępującego zakażenia [20].
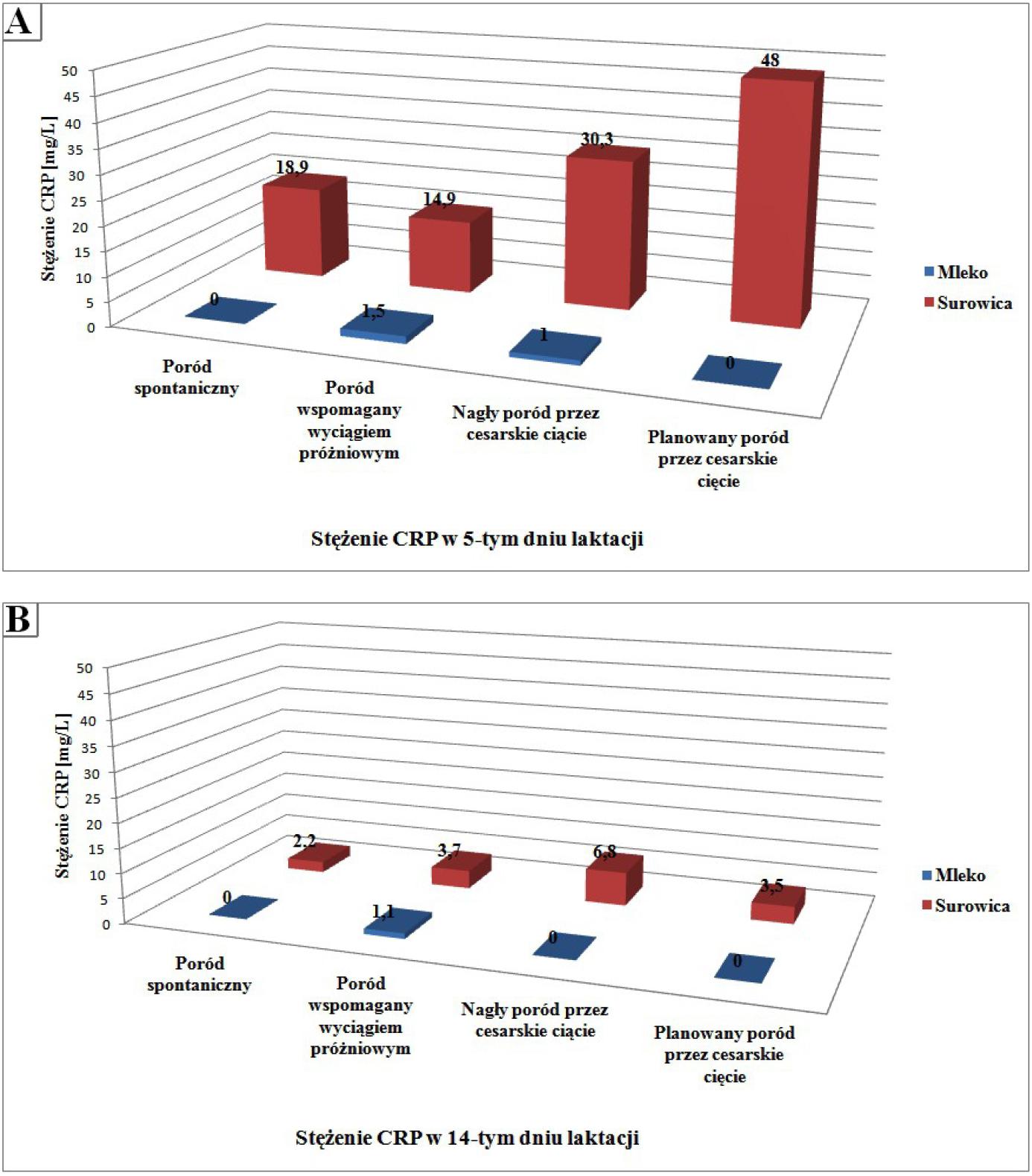
Ryc. 3. Stężenie białka C-reaktywnego w surowicy oraz w mleku ludzkim w zależności od okresu laktacji i sposobu rozwiązania ciąży. Dane dotyczą 5 (A) oraz 14 (B) dnia laktacji. Rycina własna na podstawie [38]
Alfa1-kwaśna glikoproteina
Alfa-1-kwaśna glikoproteina (AGP), białko ostrej fazy, jest produkowana głównie przez komórki wątroby, ale synteza miejscowa w innych tkankach jest również możliwa. AGP obecne w mleku pochodzi częściowo z przesączu osocza matki oraz jest wytwarzana miejscowo przez komórki nabłonkowe gruczołu sutkowego [22,112]. Stężenie AGP w siarze jest ponad 30-krotnie niższe niż w osoczu matek karmiących i ulega obniżeniu w pierwszych sześciu tygodniach laktacji. Sugeruje się, że obecne w mleku ludzkim sjalowane i fukozylowane glikotopy AGP, podobnie do wolnych oligosacharydów mleka, mogą pełnić rolę rozpuszczalnych inhibitorów dla bakteryjnych receptorów lektynowych, utrudniając adhezję i wiązanie patogenów do komórek nabłonkowych jamy ustnej noworodka [95].
Białka uczestniczące w regulacji równowagi energetycznej
W mleku ludzkim wykazano obecność białek, którym przypisuje się rolę regulacyjną i zalicza się do hormonów polipeptydowych uczestniczących w kontroli spożycia żywności i regulacji masy ciała. Należą do nich: grelina, adiponektyna, rezystyna i leptyna. Hormony znajdujące się w mleku ludzkim prawdopodobnie pochodzą z transferu z osocza matki, a częściowo są syntetyzowane miejscowo i wydzielane przez komórki nabłonkowe gruczołu sutkowego [9,65,109,113]. Leptyna i grelina, hormony zaangażowane w regulację równowagi energetycznej organizmu, działają jako przekaźniki informacji między adipocytami tkanki tłuszczowej, przewodem pokarmowym i mózgiem i w ten sposób uczestniczą w kontroli spożycia żywności i regulacji masy. Wykazano obecność receptorów dla leptyny na komórkach nabłonkowych żołądka i jelita cienkiego człowieka, co sugeruje, że leptyna może przechodzić z mleka matki do krwi niemowląt [10]. Natomiast adiponektyna i rezystyna, hormony wydzielane głównie przez adipocyty, pełnią ważną rolę w regulacji metabolizmu węglowodanów i lipidów oraz wrażliwości na insulinę. Stężenie rezystyny obecnej w mleku ludzkim koreluje dodatnio ze stężeniem leptyny oraz estradiolu, progesteronu, prolaktyny, tyroksyny, trójjodotyroniny, kortyzolu i CRP [108]. Stężenie hormonów w mleku ludzkim jest ściśle powiązane z okresem laktacji, dla rezystyny, adiponektyny i leptyny maleje wraz z laktacją, natomiast dla greliny [56] obserwuje się wzrost stężenia. Dodatkowo wykazano wyższe stężenie leptyny w mleku matek, które urodziły w terminie, w porównaniu do mleka matek, które urodziły przedwcześnie. Obecność w mleku ludzkim hormonów, takich jak leptyna, grelina, adiponektyna i rezystyna, uczestniczących w regulacji równowagi energetycznej, sugeruje, że mleko matki może być źródłem ważnych związków w rozwoju metabolicznym dziecka. Uważa się, że hormony te mogą być zaangażowane w regulację wzrostu i rozwoju w okresie noworodkowym i niemowlęcym, ochronę niemowląt przed otyłością oraz mogą mieć wpływ na długoterminowe efekty programowania metabolicznego [108,109,126].
Peptydy o właściwoścach bifidogennych
Mleko ludzkie, ze względu na obecność w nim laktoferyny, a także wydzielniczej immunoglobuliny S-IgA stymuluje również tworzenie korzystnej dla organizmu noworodka mikroflory. Badania przeprowadzone in vitro przez Liepke’go i wsp. [73] wykazały obecność w wyżej wymienionych białkach, peptydów o właściwościach bifidogennych, które wykazywały około 100 razy wyższą aktywność niż N-acetyloglukozoamina, która również pozytywnie wpływa na rozwój korzystnej dla niemowlęcia mikroflory [73].
Podsumowanie
Mleko ludzkie jest wydzieliną, której skład jest wyjątkowy i idealnie dostosowany do potrzeb noworodka pod względem składników odżywczych i bioaktywnych. Obecność w mleku ludzkim cząsteczek uczestniczących w odporności swoistej i nieswoistej, białek uczestniczących w komunikacji międzykomórkowej i regulacji równowagi energetycznej warunkuje prawidłowy rozwój niemowląt oraz wspomaga niedojrzały układ odpornościowy noworodka zapewniając tym samym skuteczną ochronę immunologiczną, a w razie zachorowania powrót do zdrowia. Z tego powodu mleko matki jest nie tylko naturalnym pokarmem zalecanym przez Amerykańską Akademię Pediatrii oraz Europejskie Stowarzyszenie Neonatologii i Perinatologii, ale również pierwszym naturalnym lekiem wykorzystywanym w opiece postnatalnej.
PIŚMIENNICTWO
[1] Abdulla E.M., Zaidi F.E., Zaidi A.: Immune factors in breast milk: a study and review. Pak. J. Med. Sci., 2005; 21: 178-186
[2] Adlerova L., Bartoskova A., Faldyna M.: Lactoferrin: a review. Vet. Med., 2008; 53: 457-468
[3] Agarwal S.K.W., Davis S., Gangur V.: Immune markers in breast milk and fetal and maternal body fluids: a systematic review of perinatal concentrations. J. Hum. Lact., 2011; 27: 171-186
[PubMed]
[4] American Academy of Pediatrics Policy on Breastfeeding and the Use of Human Milk (12.01.2013)
http://www2.aap.org/breastfeeding/policyOnBreastfeedingAndUseOfHumanMilk.html
[5] Araújo E.D., Gonçalves A.K., Cornetta Mda C., Cunha H., Cardoso M.L., Morais S.S., Giraldo P.C.: Evaluation of the secretory immunoglobulin A levels in the colostrum and milk of mothers of term and pre-term newborns. Braz. J. Infect. Dis., 2005; 9: 357-362
[PubMed] [Full Text HTML] [Full Text PDF]
[6] Artym J., Zimecki M.: Rola laktoferryny w prawidłowym rozwoju noworodka. Postępy Hig. Med. Dośw., 2005; 59: 421-432
[PubMed] [Full Text HTML] [Full Text PDF]
[7] Ashorn R.G., Eskola J., Tuohimaa P.J., Krohn K.J.: Effect of the progesterone-inducible proteins human lactoferrin and chicken avidin on lymphocyte proliferation. Hum. Reprod., 1986; 1: 149-151
[PubMed] [Full Text HTML] [Full Text PDF]
[8] Aspinall R., Prentice A.M., Ngom P.T.: Interleukin 7 from maternal milk crosses the intestinal barrier and modulates T-cell development in offspring. PLoS One, 2011; 6: e20812
[PubMed] [Full Text HTML] [Full Text PDF]
[9] Aydin S., Aydin S., Ozkan Y., Kumru S.: Ghrelin is present in human colostrum, transitional and mature milk. Peptides, 2006; 27: 878-882
[PubMed]
[10] Barrenetxe J., Villaro A.C., Guembe L., Pascual I., Munoz-Navas M., Barber A., Lostao M.P.: Distribution of the long leptin receptor isoform in brush border, basolateral membrane, and cytoplasm of enterocytes. Gut, 2002; 50: 797-802
[PubMed] [Full Text HTML] [Full Text PDF]
[11] Berenhauser A.C., Pinheiro do Prado A.C., da Silva R.C., Gioielli L.A., Block J.M.: Fatty acid composition in preterm and term breast milk. Int. J. Food Sci. Nutr., 2012; 63: 318-325
[PubMed]
[12] Blum J.W., Baumrucker C.R.: Colostral and milk insulin-like growth factors and related substances: mammary gland and neonatal (intestinal and systemic) targets. Domest. Anim. Endocrinol., 2002; 23: 101-110
[PubMed]
[13] Boesmans W., Gomes P., Janssens J., Tack J., Vanden Berghe P.: Brain-derived neurotrophic factor amplifies neurotransmitter responses and promotes synaptic communication in the enteric nervous system. Gut, 2008; 57: 314-322
[PubMed]
[14] Böttcher M.F., Jenmalm M.C., Garofalo R.P., Björkstén B.: Cytokines in breast milk from allergic and nonallergic mothers. Pediatr. Res., 2000; 47: 157-162
[PubMed]
[15] Brandtzaeg P.: Mucosal immunity: integration between mother and the breastfed infant. Vaccine, 2003; 21: 3382-3388
[PubMed]
[16] Bryan D.L., Hawkes J.S., Gibson RA.: Interleukin-12 in human milk. Pediatr. Res., 1999; 45: 858-859
[PubMed]
[17] Buescher E.S., McWilliams-Koeppen P.: Soluble tumor necrosis factor-α (TNF-α) receptors in human colostrum and milk bind to TNF-α and neutralize TNF-α bioactivity. Pediatr. Res., 1998; 44: 37-42
[PubMed]
[18] Burgoyne R.D., Duncan J.S.: Secretion of milk proteins. J. Mammary Gland Biol. Neoplasia, 1998; 3: 275-286
[PubMed]
[19] Butte N.F., Lopez-Alarcon M.G., Garza C.: Nutrient adequacy of exclusive breastfeeding for the term infant during the first six months of life. World Health Organization. Geneva 2002. http://www.who.int/nutrition/publications/infantfeeding/nut_adequacy_of_exc_bfeeding_eng.pdf (12.01.2012)
[20] Castelli G.P., Pognani C., Meisner M., Stuani A., Bellomi D., Sgarbi L.: Procalcitonin and C-reactive protein during systemic inflammatory response syndrome, sepsis and organ dysfunction. Crit. Care, 2004; 8: 234-242
[PubMed] [Full Text HTML] [Full Text PDF]
[21] Castellote C., Casillas R., Ramírez-Santana C., Pérez-Cano F.J., Castell M., Moretones M.G., López-Sabater M.C., Franch A.: Premature delivery influences the immunological composition of colostrum and transitional and mature human milk. J. Nutr., 2011; 141: 1181-1187
[PubMed] [Full Text HTML] [Full Text PDF]
[22] Ceciliani F., Pocacqua V.: The acute phase protein alpha1-acid glycoprotein: a model for altered glycosylation during diseases. Curr. Protein Pept. Sci., 2007; 8: 91-108
[PubMed]
[23] Chang C.J., Chao J.C.: Effect of human milk and epidermal growth factor on growth of human intestinal caco-2 cells. J. Pediatr. Gastroenterol. Nutr., 2002; 34: 394-401
[PubMed]
[24] Chirico G., Marzollo R., Cortinovis S., Fonte C., Gasparoni A.: Antiinfective properties of human milk. J. Nutr., 2008; 138: 1801S-1806S
[PubMed] [Full Text HTML] [Full Text PDF]
[25] Chodaczek G.: Adiuwanty jako czynniki podnoszące skuteczność szczepionek. Postępy Hig. Med. Dośw., 2004; 58: 47-59
[PubMed] [Full Text PDF]
[26] Clyne B., Olshaker J.S.: The C-reactive protein. J. Emerg. Med., 1999; 17: 1019-1025
[PubMed]
[27] Czyżewska-Buczyńska A., Lewandowicz-Uszyńska A., Jankowski A.: IgA istotny element układu odporności – wybrane zagadnienia. Postępy Hig. Med. Dośw., 2007; 61: 38-47
[PubMed] [Full Text HTML] [Full Text PDF]
[28] D’Alessandro A., Scaloni A., Zolla L.: Human milk proteins: an interactomics and updated functional overview. J. Proteome Res., 2010; 9: 3339-3373
[PubMed]
[29] Davies M.C., Arinolan G., Sanusin R., Osotimehin B.: Immunoglobulin classes and nutritional factors in plasma and breast milk of lactating mothers in Nigeria. Iran J. Immunol., 2006; 3: 181-186
[PubMed]
[30] De Leoz M.L., Gaerlan S.C., Strum J.S., Dimapasoc L.M., Mirmiran M., Tancredi D.J., Smilowitz J.T., Kalanetra K.M., Mills D.A., German J.B., Lebrilla C.B., Underwood M.A.: Lacto-N-tetraose, fucosylation, and secretor status are highly variable in human milk oligosaccharides from women delivering preterm. J. Proteome Res., 2012; 11: 4662-4672
[PubMed]
[31] Dhennin-Duthille I., Masson M., Damiens E., Fillebeen C., Spik G., Mazurier J.: Lactoferrin upregulates the expression of CD4 antigen through the stimulation of the mitogen-activated protein kinase in the human lymphoblastic T Jurkat cell line. J. Cell Biochem., 2000; 79: 583-593
[PubMed]
[32] DiBiasie A.: Evidence-based review of retinopathy of prematurity prevention in VLBW and ELBW infants. Neonatal Netw., 2006; 25: 393-403
[PubMed]
[33] Duchén K., Björkstén B.: Total IgE levels in human colostrum. Pediatr. Allergy Immunol., 1996; 7: 44-47
[PubMed]
[34] Dvorak B., Fituch C.C., Williams C.S., Hurst N.M., Schanler R.J.: Concentrations of epidermal growth factor and transforming growth factor-alpha in preterm milk. Adv. Exp. Med. Biol., 2004; 554: 407-409
[PubMed]
[35] Dziuba M., Dziuba B., Iwaniak A.: Milk proteins as precursors of bioactive peptides. Acta Sci. Pol. Technol. Aliment., 2009; 8: 71-90
[36] Elmlinger M.W., Hochhaus F., Loui A., Frommer K.W., Obladen M., Ranke M.B.: Insulin-like growth factors and binding proteins in early milk from mothers of preterm and term infants. Horm. Res., 2007; 68: 124-131
[PubMed]
[37] Emmett P.M., Rogers I.S.: Properties of human milk and their relationship with maternal nutrition. Early Hum. Dev., 1997; 49: S7-S28
[PubMed]
[38] Esaguy N., Aguas A.P., Vilanova M., Silva M.T.: Activation of human neutrophils by phorbol ester decreases the cytoplasm compactness and the lactoferrin content of the granulocytes. J. Leukoc. Biol., 1991; 50: 444-452
[PubMed] [Full Text HTML] [Full Text PDF]
[39] Fetherston C.M., Wells J.I., Hartmann P.E.: Severity of mastitis symptoms as a predictor of C-reactive protein in milk and blood during lactation. Breastfeed. Med., 2006; 1: 127-135
[PubMed]
[40] Fichter M., Klotz M., Hirschberg D.L., Waldura B., Schofer O., Ehnert S., Schwarz L.K., Ginneken C.V., Schäfer K.H.: Breast milk contains relevant neurotrophic factors and cytokines for enteric nervous system development. Mol. Nutr. Food Res., 2011; 55: 1592-1596
[PubMed]
[41] Field C.J.: The immunological components of human milk and their effect on immune development in infants. J. Nutr., 2005; 135: 1-4
[PubMed] [Full Text HTML] [Full Text PDF]
[42] França E.L., Calderon Ide M., Vieira E.L., Morceli G., Honorio-França A.C.: Transfer of maternal immunity to newborns of diabetic mothers. Clin. Dev. Immunol., 2012; 2012: 928187
[PubMed] [Full Text HTML] [Full Text PDF]
[43] França E.L., Nicomedes T.R., Calderon I.M., Honorio-França A.C.: Time-dependent alterations of soluble and cellular components in human milk. Biol. Rhythm Res., 2010; 5: 333-347
[44] Gao X., McMahon R.J., Woo J.G., Davidson B.S., Morrow A.L., Zhang Q.: Temporal changes in milk proteomes reveal developing milk functions. J. Proteome Res., 2012; 11: 3897-3907
[PubMed]
[45] Garofalo R.: Cytokines in human milk. J. Pediatr., 2010; 156: 36-40
[46] Gersting J.A., Christensen R.D., Calhoun D.A.: Effects of enterally administering granulocyte colony-stimulating factor to suckling mice. Pediatr. Res., 2004; 55: 802-806
[PubMed]
[47] Gilmore W., McKelvey-Martin V., Rutherford S., Strain J.J., Loane P., Kell M., Millar S.: Human milk contains granulocyte colony stimulating factor. Eur. J. Clin. Nutr., 1994; 48: 222-224
[PubMed]
[48] Goldman A.S.: Modulation of the gastrointestinal tract of infants by human milk. Interfaces and interactions. An evolutionary perspective. J. Nutr., 2000; 130: 426-431
[PubMed] [Full Text HTML] [Full Text PDF]
[49] Goldman A.S., Ogra P.L.: Anti-infectious and infectious agents in human milk. W: Ogra P.L., Mestecky J., Lamm M.E., Strober W., Bienenstock J., McGhee J.R.: Mucosal Immunology. Academic Press, San Diego 1999; 1511-1521
[50] Gothefors L., Marklund S.: Lactoperoxidase activity in human milk and in saliva of newborn infants. Infect. Immun., 1975; 11: 1210-1215
[PubMed] [Full Text HTML] [Full Text PDF]
[51] Hamosh M.: Protective function of proteins and lipids in human milk. Biol. Neonate, 1998; 74: 163-176
[PubMed]
[52] Haversen L., Ohlsson B.G., Hahn-Zoric M., Hanson L.A., Mattsby-Baltzer I.: Lactoferrin down-regulates the LPS-induced cytokine production in monocytic cells via NF-κB. Cell. Immunol., 2002; 220: 83-95
[PubMed]
[53] Hawkes J.S., Bryan D.L., James M.J., Gibson R.A.: Cytokines (IL-1beta, IL-6, TNF-alpha, TGF-beta1, and TGF-beta2) and prostaglandin E2 in human milk during the first three months postpartum. Pediatr. Res., 1999; 46: 194-199
[PubMed]
[54] Hirai Y., Kawakata N., Satoh K., Ikeda Y., Hisayasu S., Orimo H., Yoshino Y.: Concentrations of lactoferrin and iron in human milk at different stages of lactation. J. Nutr. Sci. Vitaminol. (Tokyo), 1990; 36: 531-544
[PubMed]
[55] Hunt K.M., Williams J.E., Shafii B., Hunt M.K., Behre R., Ting R., McGuire M.K., McGuire M.A.: Mastitis is associated with increased free fatty acids, somatic cell count, and interleukin-8 concentrations in human milk. Breastfeed. Med., 2013; 8: 105-110
[PubMed]
[56] Ilcol Y.O., Hizli B.: Active and total ghrelin concentrations increase in breast milk during lactation. Acta Pediatr., 2007; 96: 1632-1639
[PubMed]
[57] Iwaniak A., Minkiewicz P.: Biologically active peptides derived from proteins- a review. Pol. J. Food Nutr. Sci., 2008; 58: 289-294
[58] Jatsyk G.V., Kuvaeva I.B., Gribakin S.G.: Immunological protection of the neonatal gastrointestinal tract: the importance of breast feeding. Acta Paediatr. Scand., 1985; 74: 246-249
[PubMed]
[59] Jegier B.J., Meier P., Engstrom J.L., McBride T.: The initial maternal cost of providing 100ml of human milk for very low birth weight infants in the neonatal intensive care unit. Breastfeed. Med., 2010; 5: 71-77
[PubMed] [Full Text HTML] [Full Text PDF]
[60] Kadaoui K.A., Corthésy B.: Secretory IgA mediates bacterial translocation to dendritic cells in mouse Peyer’s patches with restriction to mucosal compartment. J. Immunol., 2007; 179: 7751-7757
[PubMed] [Full Text HTML] [Full Text PDF]
[61] Kanyshkova T.G., Buneva V.N., Nevinsky G.A.: Lactoferrin and its biological functions. Biochemistry, 2001; 1: 1-7
[PubMed]
[62] Kardar G., Oraei M., Shahsavani M., Namdar Z., Kazemisefat G., Haghi Ashtiani M., Shams S., Pourpak Z., Moin M.: Reference intervals for serum immunoglobulins IgG, IgA, IgM and complements C3 and C4 in Iranian healthy children. Iran J. Public Health, 2012; 41: 59-63
[PubMed] [Full Text HTML] [Full Text PDF]
[63] Kehrl J.H., Roberts A.B., Wakefield L.M., Jakowlew S., Sporn M.B., Fauci A.S.: Transforming growth factor beta is an important immunomodulatory protein for human B lymphocytes. J. Immunol., 1986; 137: 3855-3860
[PubMed]
[64] Khailova L., Dvorak K., Arganbright K.M., Williams C.S., Halpern M.D., Dvorak B.: Changes in hepatic cell junctions structure during experimental necrotizing enterocolitis: effect of EGF treatment. Pediatr. Res., 2009; 66: 140-144
[PubMed] [Full Text HTML] [Full Text PDF]
[65] Kierson J.A., Dimatteo D.M., Locke R.G., Mackley A.B., Spear M.L.: Ghrelin and cholecystokinin in term and preterm human breast milk. Acta Paediatr., 2006; 95: 991-995
[PubMed]
[66] Klaska I., Nowak J.Z.: Rola układu dopełniacza w fizjologii i patologii. Postępy Hig. Med. Dośw., 2007; 61: 167-177
[PubMed] [Full Text HTML] [Full Text PDF]
[67] Kling P.J., Taing K.M., Dvorak B., Woodward S.S., Philipps A.F.: Insulin-like growth factor-I stimulates erythropoiesis when administered enterally. Growth Factors, 2006; 24: 218-223
[PubMed]
[68] Kverka M., Burianova J., Lodinova-Zadnikova R., Kocourkova I., Cinova J., Tuckova L., Tlaskalova-Hogenova H.: Cytokine profiling in human colostrum and milk by protein array. Clin. Chem., 2007; 53: 955-962
[PubMed] [Full Text HTML] [Full Text PDF]
[69] Lam E.Y., Kecskes Z., Abdel-Latif M.E.: Breast milk banking: Current opinion and practice in Australian neonatal intensive care units. J. Paediatr. Child Health, 2012; 48: 833-839
[PubMed]
[70] Legrand D., Mazurier J.: A critical review of the roles of host lactoferrin in immunity. Biometals, 2010; 23: 365-376
[PubMed]
[71] Li R., Xia W., Zhang Z., Wu K.: S100B protein, brain-derived neurotrophic factor, and glial cell line-derived neurotrophic factor in human milk. PLoS One, 2011; 6: e21663
[PubMed] [Full Text HTML] [Full Text PDF]
[72] Liao Y., Alvarado R., Phinney B., Lönnerdal B.: Proteomic characterization of human milk whey proteins during a twelve-month lactation period. J. Proteome Res., 2011; 10: 1746-1754
[PubMed]
[73] Liepke C., Adermann K., Raida M., Mägert H.J., Forssmann W.G., Zucht H.D.: Human milk provides peptides highly stimulating the growth of bifidobacteria. Eur. J. Biochem., 2002; 269: 712-718
[PubMed] [Full Text HTML] [Full Text PDF]
[74] Lönnerdal B.: Nutritional and physiologic significance of human milk proteins. Am. J. Clin. Nutr., 2003; 77: 1537-1543
[PubMed] [Full Text HTML] [Full Text PDF]
[75] Lönnerdal B.: Human milk proteins: Key components for the biological activity of human milk. Adv. Exp. Med. Biol., 2004; 554: 11-25
[PubMed]
[76] Lönnerdal B., Lien E.L.: Nutritional and physiologic significance of alpha-lactalbumin in infants. Nutr. Rev., 2003; 61: 295-305
[PubMed]
[77] Loui A., Eilers E., Strauss E., Pohl-Schickinger A., Obladen M., Koehne P.: Vascular endothelial growth factor (VEGF) and soluble VEGF Receptor 1 (Sflt-1) levels in early and mature human milk from mothers of preterm versus term infants. J. Hum. Lact., 2012; 28: 522-528
[PubMed]
[78] Machnicki M., Zimecki M., Zagulski T.: Lactoferrin regulates the release of tumour necrosis factor alpha and interleukin 6 in vivo. Int. J. Exp. Pathol., 1993; 74: 433-439
[PubMed] [Full Text HTML] [Full Text PDF]
[79] Maheshwari A., Lu W., Lacson A., Barleycorn A.A., Nolan S., Christensen R.D., Calhoun D.A.: Effects of Interleukin-8 on the developing human intestine. Cytokine, 2002; 20: 256-267
[PubMed]
[80] McGreal E.P., Hearne K., Spiller O.B.: Off to a slow start: Under-development of the complement system in term newborns is more substantial following premature birth. Immunobiology, 2012; 217: 176-186
[PubMed]
[81] McManaman J.L., Neville M.C.: Mammary physiology and milk secretion. Adv. Drug Deliv. Rev., 2003; 55: 629-641
[PubMed]
[82] Mehra R., Marnila P., Korhonen H.: Milk immunoglobulins for health promotion. Int. Dairy J., 2006; 16: 1262-1271
[83] Meki A.R., Saleem T.H., Al-Ghazali M.H., Sayed A.A.: Interleukins -6, -8 and -10 and tumor necrosis factor alpha and its soluble receptor I in human milk at different periods of lactation. Nutr. Res., 2003; 23: 845-855
[84] Mickleson K.N., Moriarty K.M.: Immunoglobulin levels in human colostrum and milk. J. Pediatr. Gastroenterol. Nutr., 1982; 1: 381-384
[PubMed]
[85] Milsom S.R., Blum W.F., Gunn A.J.: Temporal changes in insulin-like growth factors I and II and in insulin-like growth factor binding proteins 1, 2, and 3 in human milk. Horm. Res., 2008; 69: 307-311
[PubMed]
[86] Minkiewicz P., Dziuba J., Darewicz M., Iwaniak A., Dziuba M., Nałęcz D.: Food peptidomics. Food Technol. Biotechnol., 2008; 46: 1-10
[87] Mizuno K., Hatsuno M., Aikawa K., Takeichi H., Himi T., Kaneko A., Kodaira K., Takahashi H., Itabashi K.: Mastitis is associated with IL-6 levels and milk fat globule size in breast milk. J. Hum. Lact., 2012; 28: 529-534
[88] Molinari C.E., Casadio Y.S., Hartmann B.T., Livj A., Bringans S., Arthur P.G., Hartmann P.E.: Proteome mapping of human skim milk proteins in term and preterm milk. J. Proteome Res., 2012; 11: 1696-1714
[PubMed]
[89] Montagne P., Cuilliere M.L., Mole C., Bene M.C., Faure G.: Changes in lactoferrin and lysozyme levels in human milk during the first twelve weeks of lactation. Adv. Exp. Med. Biol., 2001; 501: 241-247
[PubMed]
[90] Morceli G., França E.L., Magalhaes V.B., Damasceno D.C., Calderon I.M., Honorio-França A.C.: Diabetes induced immunological and biochemical changes in human colostrum. Acta Paediatr., 2011; 100: 550-556
[PubMed]
[91] Nakamura Y., Miyata M., Ando T., Shimokawa N., Ohnuma Y., Katoh R., Ogawa H., Okumura K., Nakao A.: The latent form of transforming growth factor-β administered orally is activated by gastric acid in mice. J. Nutr., 2009; 139: 1463-1468
[PubMed] [Full Text HTML] [Full Text PDF]
[92] Ogawa J., Sasahara A., Yoshida T., Sira M.M., Futatani T., Kanegane H., Miyawaki T.: Role of transforming growth factor-beta in breast milk for initiation of IgA production in newborn infants. Early Hum. Dev., 2004; 77: 67-75
[PubMed]
[93] Ogundele M.O.: Anti-complement activities of human breast-milk. Inflamm. Res., 1999; 48: 437-445
[PubMed]
[94] Ogundele M.O.: Inhibitors of complement activity in human breast-milk: a proposed hypothesis of their physiological significance. Mediators Inflamm., 1999; 8: 69-75
[PubMed] [Full Text HTML] [Full Text PDF]
[95] Orczyk-Pawiłowicz M., Hirnle L., Kątnik-Prastowska I.: Human milk α1-acid glycoprotein lectin-reactivity during transition from colostrum to mature milk. W: 16th ISRHML: Breastfeeding and the use of human milk. Science & practice 2012. Program and abstracts book (14.01.2013)
https://www.isrhml.org/wp-content/uploads/2012/11/Abstract-book-ISRHML-website.pdf
[96] Pan Y., Lee A., Wan J., Coventry M.J., Michalski W.P., Shiell B., Roginski H.: Antiviral properties of milk proteins and peptides. Int. Dairy J., 2006; 16: 1252-1261
[97] Pawlus B., Kordek A., Łoniewska B.: Enzymy mleka kobiecego – najnowsze wiadomości. Przegl. Pediatr., 2005; 35: 168-170
[Abstract]
[98] Pawlus B., Kordek A., Łoniewska B.: Podstawowe składniki mleka kobiecego – najnowsze wiadomości. Medycyna Rodzinna, 2004; 5: 213-216
[Full Text HTML]
[99] Penttila I.A.: Milk-derived transforming growth factor-β and the infant immune response. J. Pediatr., 2010; 156: 21-25
[100] Prokesová L., Lodinová-Zádníková R., Zizka J., Kocourková I., Novotná O., Petrásková P., Sterzl I.: Cytokine levels in healthy and allergic mothers and their children during the first year of life. Pediatr. Allergy Immunol., 2006; 17: 175-183
[PubMed]
[101] Prosser C.G.: Insulin-like growth factors in milk and mammary gland. J. Mammary Gland Biol. Neoplasia, 1996; 1: 297-306
[PubMed]
[102] Reddy V., Bhaskaram C., Raghuramulu N., Jagadeesan V.: Antimicrobial factors in human milk. Acta Paediatr. Scand., 1977; 66: 229-232
[PubMed]
[103] Reynolds J.D.: The management of retinopathy of prematurity. Paediatr. Drugs, 2001; 3: 263-272
[PubMed]
[104] Richie E.R., Hilliard J.K., Gilmore R., Gillespie D.J.: Human milk-derived lactoferrin inhibits mitogen and alloantigen induced human lymphocyte proliferation. J. Reprod. Immunol., 1987; 12: 137-148
[PubMed]
[105] Rigotti E., Piacentini G.L., Ress M., Pigozzi R., Boner A.L., Peroni D.G.: Transforming growth factor-β1 and interleukin-10 in breast milk and development of atopic diseases in infants. Clin. Exp. Allergy, 2006; 36: 614-618
[PubMed]
[106] Rodrigues D., Li A., Nair D., Blennerhassett M.G.: Glial cell line-derived neurotrophic factor is a key neurotrophin in the postnatal enteric nervous system. Neurogastroenterol. Motil., 2011; 23: e44-e56
[PubMed]
[107] Sandström O., Lönnerdal B., Graverholt G., Hernell O.: Effects of α-lactalbumin-enriched formula containing different concentrations of glycomacropeptide on infant nutrition. Am. J. Clin. Nutr., 2008; 87: 921-928
[PubMed] [Full Text HTML] [Full Text PDF]
[108] Savino F., Liguori S.A., Petrucci E., Lupica M.M., Fissore M.F., Oggero R., Silvestro L.: Evaluation of leptin in breast milk, lactating mothers and their infants. Eur. J. Clin. Nutr., 2010; 64: 972-977
[PubMed]
[109] Savino F., Lupica M.M., Benetti S., Petrucci E., Liguori S.A., Cordero Di Montezemolo L.: Adiponectin in breast milk: relation to serum adiponectin concentration in lactating mothers and their infants. Acta Paediatr., 2012; 101: 1058-1062
[PubMed]
[110] Schack L., Lange A., Kelsen J., Agnholt J., Christensen B., Petersen T.E., Sorensen E.S.: Considerable variation in the concentration of osteopontin in human milk, bovine milk and infant formulas. J. Dairy Sci., 2009; 92: 5378-5385
[PubMed]
[111] Séverin S., Wenshui X.: Milk biologically active components as nutraceuticals. Crit. Rev. Food Sci. Nutr., 2005; 45: 645-656
[PubMed]
[112] Shiyan S.D., Bovin N.V.: Carbohydrate composition and immunomodulatory activity of different glycoforms of alpha1-acid glycoprotein. Glycoconj. J., 1997; 14: 631-638
[PubMed]
[113] Smith-Kirwin S.M., O’Connor D.M., De Johnston J., Lancey E.D., Hassink S.G., Funanage V.L.: Leptin expression in human mammary epithelial cells and breast milk. J. Clin. Endocrinol. Metab., 1998; 83: 1810-1813
[PubMed] [Full Text HTML] [Full Text PDF]
[114] Stelwagen K., Carpenter E., Haigh B., Hodgkinson A., Wheeler T.T.: Immune components of bovine colostrum and milk. J. Anim. Sci., 2009; 87: 3-9
[PubMed] [Full Text HTML] [Full Text PDF]
[115] Srivastava M.D., Brouhard B., Saneto R., Groh-Wargo S., Kubit J.: Cytokines in human milk. Res. Commun. Mol. Pathol. Pharmacol., 1996; 93: 263-287
[PubMed]
[116] Struck J., De Almeida P., Bergmann A., Morgenthaler N.G.: High concentration of procalcitonin but not calcitonin in normal human milk. Horm. Metab. Res., 2002; 34: 460-465
[PubMed]
[117] The Union of European Neonatal & Perinatal Societies (UENPS) (10.01.2013)
http://www.uenps.com
[118] Trégoat V., Montagne P., Béné M.C., Faure G.: Changes in the mannan binding lectin (MBL) concentration in human milk during lactation. J. Clin. Lab. Anal., 2002; 16: 304-307
[PubMed]
[119] Trégoat V., Montagne P., Béné M.C., Faure G.: Increases of IgA milk concentrations correlate with IgA2 increment. J. Clin. Lab. Anal., 2001; 15: 55-58
[PubMed]
[120] Trégoat V., Montagne P., Cuilliere M.L., Béné M.C., Faure G.C.: Sequential C3 and C4 levels in human milk in relation to prematurity and parity. Clin. Chem. Lab. Med., 2000; 38: 609-613
[PubMed]
[121] Trelińska J., Bodalski J.: Rola prokalcytoniny w przebiegu reakcji zapalnej. Przegl. Pediatr., 2007; 37: 314-317
[Abstract]
[122] Uchiyama S., Sekiguchi K., Akaishi M., Anan A., Maeda T., Izumi T.: Characterization and chronological changes of preterm human milk gangliosides. Nutrition, 2011; 27: 998-1001
[PubMed]
[123] Ustundag B., Yilmaz E., Dogan Y., Akarsu S., Canatan H., Halifeoglu I., Cikim G., Aygun A.D.: Levels of cytokines (IL-1beta, IL-2, IL-6, IL-8, TNF-alpha) and trace elements (Zn, Cu) in breast milk from mothers of preterm and term infants. Mediators Inflamm., 2005; 2005: 331-336
[PubMed] [Full Text HTML] [Full Text PDF]
[124] van Neerven R.J., Knol E.F., Heck J.M., Savelkoul H.F.: Which factors in raw cow’s milk contribute to protection against allergies? J. Allergy Clin. Immunol., 2012; 130: 853-858
[PubMed]
[125] Vordenbäumen S., Braukmann A., Petermann K., Scharf A., Bleck E., von Mikecz A., Jose J., Schneider M.: Casein α s1 is expressed by human monocytes and upregulates the production of GM-CSF via p38 MAPK. J. Immunol., 2011; 186: 592-601
[PubMed] [Full Text HTML] [Full Text PDF]
[126] Weyermann M., Beermann C., Brenner H., Rothenbacher D.: Adiponectin and leptin in maternal serum, cord blood, and breast milk. Clin. Chem., 2006; 52: 2095-2102
[PubMed] [Full Text HTML] [Full Text PDF]
[127] Wrodycki W.: Przydatność oznaczania stężenia prokalcytoniny (PCT) w surowicy krwi u chorych diagnozowanych w oddziałach obserwacyjno-zakaźnych. Przegl. Epidemiol., 2003; 57: 211-219
[Full Text PDF]
[128] Yilmaz H.L., Saygili-Yilmaz E.S., Gunesacar R.: Interleukin-10 and -12 in human milk at 3 stages of lactation: a longitudinal study. Adv. Ther., 2007; 24: 603-610
[PubMed]
[129] Zimecki M., Mazurier J., Machnicki M., Wieczorek Z., Montreuil J., Spik G.: Immunostimulatory activity of lactotransferrin and maturation of CD4–CD8– murine thymocytes. Immunol. Lett., 1991; 30: 119-123
[PubMed]
[130] Zimecki M., Mazurier J., Spik G., Kapp J.A.: Human lactoferrin induces phenotypic and functional changes in murine splenic B cells. Immunology, 1995; 86: 122-127
[PubMed] [Full Text HTML] [Full Text PDF]
Autorzy deklarują brak potencjalnych konfliktów interesów.