
Stem and progenitor cells in biostructure of blood vessel walls
Krzysztof Korta 1 , Piotr Kupczyk 2 , Jan Skóra 1 , Artur Pupka 1 , Paweł Zejler 3 , Marcin Hołysz 4 , Mariusz Gajda 5 , Beata Nowakowska 2 , Piotr Barć 1 , Andrzej T. Dorobisz 1 , Tomasz Dawiskiba 1 , Piotr Szyber 1 , Julia Bar 4Streszczenie
Rozwój układów naczyniowego i hematopoetycznego w czasie organogenezy odbywa się jednocześnie. Podczas waskulogenezy, część komórek nie podlega procesowi różnicowania się, a pozostaje na bardzo wczesnym etapie „zakotwiczona” w strukturach tkankowych, określanych jako nisze tkankowe komórek macierzystych i progenitorowych. Obecność naczyń krwionośnych w obrębie niszy tkankowych jest cechą charakterystyczną, która pozwala na identyfikację niszy, a także zapewnia jej funkcjonowanie. Wchodzące w skład biostruktury ściany naczyń żylnych i tętniczych tunica: intima, media i adventitia przez długi czas były definiowane, jako mechaniczne bariery oddzielające światło naczynia od lokalnego środowiska tkankowego. Badania z zakresu biologii naczyń z ostatniego okresu wykazały, że ściana naczynia jest dynamiczną biostrukturą, która zawiera w komórki macierzyste, opisywane jako rezydentne/spoczynkowe macierzyste i progenitorowe komórki ścian naczyń krwionośnych (VW-SC/PC). Różne obszary naczynia zawierają heterogenne populacje komórek WV-SC/PC, a najważniejsze z nich opisywane są jako strefy: okołosródbłonkowa i waskulogenna. Liczne doświadczenia prowadzone in vitro i in vivo wskazują, że aktywność komórek macierzystych nie jest ograniczona wyłącznie do organogenezy, ale przejawia się w okresie postnatalnym, gdzie odpowiada za homeostazę, przebudowę oraz regenerację ściany naczynia. Uważa się również, że komórki WV-SC/PC mogą być zaangażowane w progresję i rozwój chorób naczyniowych np. tworzenie neointima.
W pracy podsumowano wiadomości o fenotypie komórek mezenchymalnych i progenitorowych, rozmieszczeniu i biologicznych właściwościach VW-SC/PC w tunica: intima, media i adventitia. Uważa się, że w niedalekiej perspektywie nisze z komórkami VW-SC/PC mogą się stać źródłem pozyskiwania komórek macierzystych i progenitorowych do wykorzystania w naczyniowej bioinżynierii tkankowej, jako alternatywa wobec tradycyjnych metod rewaskularyzacyjnych.
Summary
Development of vascular and hematopoietic systems during organogenesis occurs at the same time. During vasculogenesis, a small part of cells does not undergo complete differentiation but stays on this level, “anchored” in tissue structures described as stem cell niches. The presence of blood vessels within tissue stem cell niches is typical and led to identification of niches and ensures that they are functioning. The three-layer biostructure of vessel walls for artery and vein, tunica: intima, media and adventitia, for a long time was defined as a mechanical barrier between vessel light and the local tissue environment. Recent findings from vascular biology studies indicate that vessel walls are dynamic biostructures, which are equipped with stem and progenitor cells, described as vascular wall-resident stem cells/progenitor cells (VW-SC/PC). Distinct zones for vessel wall harbor heterogeneous subpopulations of VW-SC/ PC, which are described as “subendothelial or vasculogenic zones”. Recent evidence from in vitro and in vivo studies show that prenatal activity of stem and progenitor cells is not only limited to organogenesis but also exists in postnatal life, where it is responsible for vessel wall homeostasis, remodeling and regeneration. It is believed that VW-SC/PC could be engaged in progression of vascular disorders and development of neointima. We would like to summarize current knowledge about mesenchymal and progenitor stem cell phenotype with special attention to distribution and biological properties of VW-SC/PC in biostructures of intima, media and adventitia niches. It is postulated that in the near future, niches for VW-SC/PC could be a good source of stem and progenitor cells, especially in the context of vessel tissue bioengineering as a new alternative to traditional revascularization therapies.
Key words:vessel wall-stem cell/progenitor cell • tissue bioengineering • tunica: intima • media • adventitia • mesenchymal stem cell • stem cell niches • endothelial progenitor cell • pericytes • perivascular cell • angiogenesis • hemangioblast • tissue remodeling • neointima • vein • artery
Wykaz skrótów:
ACE – enzym konwertujący angiotensynę (angiotensin converting enzyme); AGM – aorta-gonad-mesonephros; BM-EPC – komórki progenitorowe sródbłonka naczyniowego ze szpiku kostnego (bone marrow – endothelial progenitor cells); C-EPC – cyrkulujące-komórki progenitorowe sródbłonka naczyniowego (circulating endothelial progenitor cells); CEACAM-1 – molekuła adhezji komórkowej antygenu karcynoembrionalnego 1 (carcinoembryonic antigen-related cell adhesion molecule 1); CFU – jednostki formowania kolonii (colony forming units); CXCL – C-X-C ligand; CXCR – C-X-C receptor; ECM – macierz międzykomórkowa (extra cellular matrix); ESC – embrionalne komórki macierzyste (embrionic stem cells); FACS – fluorescencyjny sorter komórkowy (fluorescence-activated cell sorting); FGF – czynnik wzrostu fibroblastów (fibroblast growth factor); FGFR – receptor czynnika wzrostu fibroblastów (fibroblast growth factor receptor); GFP – białko zielonej fluorescencji (green fluorescent protein); GM-CSF – granulocytowo-makrofagowy czynnik wzrostu (granulocyte-macrophage colony-stimulating factor); HAEC – ludzkie komórki sródbłonka aorty (human aortic endothelial cell); HDA1 – ludzki antygen skóry 1 (human dermal antigen 1); HLA – cząsteczki klasy I głównego kompleksu zgodnosci tkankowej człowieka (human leukocytes antigens class I molecules); HSC – komórki macierzyste hematopoezy (hematopoietic stem cells); HSCT – transplantacje komórek macierzystych hematopoezy (hematopoietic stem cell transplantation); HUVEC – ludzkie komórki sródbłonka żyły pępowinowej (human umbilical vein endothelial cells); ISCT – Międzynarodowe Towarzystwo Terapii Komórkowej (International Society For Cell Therapy); KGF – czynniki wzrostu keratynocytów (keratinocyte growth factor); MPP – białka metaloproteinaz (metaloproteinase proteins); MSC – mezenchymalne komórki macierzyste (mesenchymal stem cells); NG2 – neuralny antigen glejowy 2 (neural glial antigen 2); PDGF-B – czynnik wzrostu pochodzenia łożyskowego B (placenta-derived growth factor-B); PVC – komórki perywaskularne (perivascular cells); PBMC – komórki jednojądrzaste krwi obwodowej (peripheral blood mononuclear cells); SCID – ciężkie złożone niedobory odporności (severe combined immunodeficiency); SCL – komórki macierzyste białaczki (stem cell leukemia); SDF-1 – czynnik wzrostowy pochodzenia stromalnego 1 (stromal derived factor-1); Shh – sygnalizacja Shh (donic hedgehog), SMC – komórki mięsniówki gładkiej (smooth muscle cells); α-SMA -α-aktyna mięśni gładkich (α-smooth muscle actin); SMP – komórki progenitorowe mięśni gładkich (smooth muscle progenitors); UCB – krew pępowinowa (umbilical cord blood); UC-EPC – komórki progenitorowe sródbłonka naczyniowego z krwi pępowinowej (umbilical cord-endothelial progenitor cells); UEA-1 – aglutynina Ulex Europeaus 1 (Ulex Europaeus Agglutinin-1); VEGF – czynnik wzrostu sródbłonka naczyniowego (vascular endothelial growth factor); VESC – komórki sródbłonka żyły odpiszczelowej (vessel endothelial saphenous cell); VTB-E – naczyniowa bioinżynieria tkankowa (vascular tissue bioenginery); VW-MPSC – multipotencjalne komórki macierzyste ściany naczynia (vessel wal-multipotent stem cell); VW-SC/PC – komórki macierzyste/komórki progenitorowe ściany naczynia (vessel wall-stem cell/progenitor cell).
Wstęp
Organogeneza układu sercowo-naczyniowego i hematopoetycznego przebiega jednocześnie w okresie embrionalnym. W tym samym czasie w obrębie zachodzących przemian można wyodrębnić proces waskulogenezy, czyli formowania się struktur pierwotnego układu naczyniowego z wykorzystaniem komórek o charakterze macierzystym i progenitorowym. Oba układy: naczyniowy i hematopoetyczny w okresie embrionalnym formują się i różnicują ze wspólnej masy komórek pochodzenia mezodermalnego, która nazywana jest hemangioblastem. Definicja hemangioblastu jest niejednoznaczna. Dotychczasowe wyniki badań wskazują, że komórki z ekspresją markerów CD34+, CD309+, CD133+ mogą niewystarczająco odzwierciedlać właściwości hemangioblastu. W opinii niektórych autorów komórki z ekspresją CD34+, CD309+, CD133+ stanowią jedynie boczną populację, której właściwości biologiczne zbliżone są z linią hematopoetyczną [34,36]. Powyższe obserwacje wskazują, że istotne jest prowadzenie badań zmierzających do zdefiniowania markerów, które pozwoliłyby na ustalenie precyzyjnej granicy między linią naczyniową i hematopoetyczną, a także opisanie markerów typowych dla komórek macierzystych i progenitorowych.
W okresie prenatalnym wzmożona aktywność komórek macierzystych i progenitorowych prowadzi do powstania pierwszych, pierwotnych struktur naczyniowych. W kolejnych etapach procesu ontogenezy dochodzi do różnicowania komórek i nadawania im „funkcjonalnego przeznaczenia”.
W życiu postnatalnym jedynie niewielka liczba komórek w obrębie zróżnicowanych tkanek narządów stanowi tzw. pule spoczynkowych komórek macierzystych lub progenitorowych. Komórki te nie zostały poddane procesowi definitywnego różnicowania się, lecz zostały „zakotwiczone” w środowisku zróżnicowanych tkanek i narządów [29].
Przez długi czas panowało przekonanie, że powstawanie naczyń krwionośnych w okresie prenatalnym i związana z nim aktywność komórek macierzystych są zjawiskami ograniczonymi do etapu organogenezy [74,77,78]. Pierwsze informacje o zdolnościach regeneracyjnych naczyń krwionośnych w życiu postnatalnym pojawiły się w 1988 r., kiedy to Simionescu M. i Simionescu M.L.W. sformułowali hipotezę, według której w obrębie trójwarstwowej biostruktury naczynia krwionośnego: tunica intima, tunica media i tunica adventitia mogą się znajdować populacje komórek o zdolnościach regeneracyjnych [68,69,70]. Obecnie prawdopodobieństwo występowania komórek o potencjale regeneracyjnym w obrębie biostruktury naczynia krwionośnego jest udokumentowane [1,17,24,25,26,34,39,40,49,50,59,79,80,81,83,88]. Zaangażowanie układu krwionośnego w organogenezę [77,78] oraz zdolności regeneracyjne ściany naczynia wywołane np. warunkami hemodynamicznymi wynikającymi z przepływu krwi to tylko niektóre przykłady potwierdzające tezę, że ściana naczynia zarówno w okresie prenatalnym, jak i postnatalnym jest biostrukturą dynamiczną [2]. W obu przypadkach aktywność komórek macierzystych lub progenitorowych jest niezbędnym czynnikiem kształtującym i utrzymującym prawidłową homeostazę ściany naczynia.
Komórki o różnym potencjale i na różnym etapie różnicowania, opisane w biostrukturze naczyń, przedstawiane są jako rezydentne/spoczynkowe macierzyste i progenitorowe komórki ścian naczyń krwionośnych VW-SC/ PC [1,14,24,29,39,41,48,54,79,81,83,86,88]. Identyfikacja komórek progenitorowych dla śródbłonków naczyniowych we krwi obwodowej C-EPC [3], szpiku kostnym BM-EPC, krwi pępowinowej UC-EPC, a także odkrycie klastrów komórek macierzystych i progenitorowych w obrębie zróżnicowanych tkanek i narządów [29,73,79] stanowi wystarczający dowód, że mechanizmy, których aktywność pozwala na rozwój organizmu w okresie życia prenatalnego są również obecne i aktywne w życiu postnatalnym [88]. Komórki VW-SC/PC spoczywające w środowisku nisz komórkowych zróżnicowanych tkanek i narządów, w tym również w obrębie biostruktur naczyń żylnych i tętniczych, są najprawdopodobniej pierwszą bezpośrednią formą zaangażowaną w rekonstrukcje tkankowe w przypadku uszkodzenia lub zmian wywołanych chorobą, na bardzo wczesnych etapach jej rozwoju [80,86]. Należy podkreślić, że wyniki badań na modelach in vitro i in vivo wykazały, że VW-SC/PC mogą być również zaangażowane w rozwój chorób naczyniowych i nowotworowych [11].
Zastosowanie komórek macierzystych i progenitorowych do celów terapeutycznych jest przedmiotem zainteresowania specjalistów z zakresu transplantologii, angiologii i angiochirurgii oraz biologów i biotechnologów, którzy wspólnie podejmują się idei tworzenia tzw. naczyniowej bioinżynierii tkankowej VTB-E. Tradycyjne interwencje chirurgiczne oraz farmakoterapia u części pacjentów ze schorzeniami naczyniowymi zawodzą, zwłaszcza w terapii naczyń o średnicy poniżej 10 mm. Wykorzystanie VTB-E w celu rekonstrukcji naczyniowych może się okazać skutecznym narzędziem terapeutycznym, a także ograniczyć lub wyeliminować powikłania wynikające z tradycyjnego leczenia rewaskularyzacyjnego [46].
Waskulogeneza
Waskulogeneza, czyli proces formowania się i powstawania pierwotnego układu naczyniowego w wyniku agregacji komórek macierzystych podczas embriogenezy rozpoczyna się tuż po gastrulacji, kiedy pierwotne komórki tylnej i bocznej mezodermy rozpoczynają migrację względem pierwotnego pęcherzyka żółtkowego (yolk sac). Komórki mezodermy agregują i rozpoczynają tworzenie tzw. wysp krwiotwórczych (blood islands). Wyspy krwiotwórcze stanowią jednolitą masę komórek, które w kolejnym etapie podlegają procesowi różnicowania się. Komórki usytuowane w centrum wysp krwiotwórczych stają się zawiązkiem pierwotnego układu hematopoetycznego, natomiast komórki usytuowane obwodowo stają się źródłem komórek układu sercowo-naczyniowego, tzw. angioblastów. Angioblasty w obrębie wysp krwiotwórczych ulegają dalszej reorganizacji, co prowadzi do formowania się pierwszych tzw. prymitywnych splotów naczyniowych (vascular plexus) [13,41,75,85]. To na poziomie angioblastów i formowania się pierwotnych splotów naczyniowych dochodzi najprawdopodobniej do uruchomienia molekularnych i genetycznych mechanizmów odpowiedzialnych za powstanie trójwarstwowej biostruktury naczyń krwionośnych, a także definiowania śródbłonka żylnego i tętniczego [41,43,49,50].
W kolejnych etapach waskulogenezy migracja i różnicowanie angioblastów z różnych obszarów mezodermy doprowadzają do powstania małych i dużych struktur naczyniowych i limfatycznych, endokardium oraz aorty grzbietowej (dorsal aorta) [41,45,59,84]. Aorta grzbietowa znajduje się w obrębie regionu AGM, który rozwija się z mezodermalnej paraaortycznej otrzewnej trzewnej (splanchnopleura). Obszar AGM, w obrębie, którego znajduje się aorta płodowa, zawiązki genitaliów oraz pranercze (mesonephros) jest szczególnie interesującym obszarem z punktu widzenia rozwijającego się układu naczyniowego embrionu. Na wczesnym etapie waskulogenezy dochodzi tu do uruchomienia mechanizmów, w wyniku których ściana aorty przejmuje aktywność proliferacyjną stając się źródłem dla pierwotnych komórek hematopoezy [72,85] (ryc.1). Uruchomianie aktywności hematopoetycznej komórek ścian aorty płodowej koreluje z pojawieniem się ekspresji genu Runx- 1, który najprawdopodobniej inicjuje mechanizm tzw. przejścia śródbłonkowo-hematopoetycznego (endothelial hematopoietic transition) [38,59,75,76,77,78]. Aktywność hematopoetyczna biostruktur aorty płodowej jest przejściowa, gdyż w kolejnych etapach embriogenezy pierwotne komórki hematopoezy migrują i zasiedlają docelowe dla nich nisze w jamach szpikowych. Przejściowa aktywność proliferacyjna ściany aorty potwierdza wspólny i ściśle zależny rozwój układu sercowo-naczyniowego oraz hematopoetycznego, co jest ważnym dowodem na istnienie ich wspólnego przodka, hemangioblastu [13,41,54].

Ryc. 1. Schemat przedstawia masę komórek pochodzenia mezodermalnego (1), z których w wyniku procesów różnicowania się dochodzi do formowania pierwotnego układu sercowo-naczyniowego z komórek usytuowanych obwodowo oraz pierwotnego układu hematopoetycznego z komórek centralnych (2). W dalszym etapie wspólnego rozwoju obu układów dochodzi do zasiedlenia komórek i wystąpienie chwilowej aktywności hematopoetycznej przez ścianę aorty płodowej (3)
Hemangioblast
Organogeneza układu sercowo-naczyniowego oraz hematopoetycznego podczas życia embrionalnego stały się inspiracją do poszukiwań wspólnego „przodka”, bipotencjalnej komórki, tzw. hemangioblastu, który jest początkiem linii komórek hematopoetycznych oraz linii komórek prowadzącej do powstania zróżnicowanych komórek śródbłonków naczyniowych [13,41] (ryc.2). Badania eksperymentalne z wykorzystaniem embrionów danio pręgowanego, myszy, a także ludzkich embrionalnych komórek macierzystych ESC dowodzą obecności komórek o bipotencjalnym profilu również w okresie postnatalnym [6,12]. Obecnie wiadomo, że hemangioblast nie jest komórką przejściową, która występuje wyłącznie podczas embriogenezy [12,41,75,85]. Obecność hemangioblastu potwierdzono pod względem fenotypowym i molekularnym w komórkach i tkankach zróżnicowanych. W komórkach ESC wykazano kilka genów i markerów, które uczestniczą w regulacji i różnicowaniu właściwości komórki hemangioblastu: gen Runx-1, gen komórki macierzystej białaczki SCL, gen mezodermalny T oraz enzym konwertujący angiotensynę ACE/ CD143 [54,59,71].
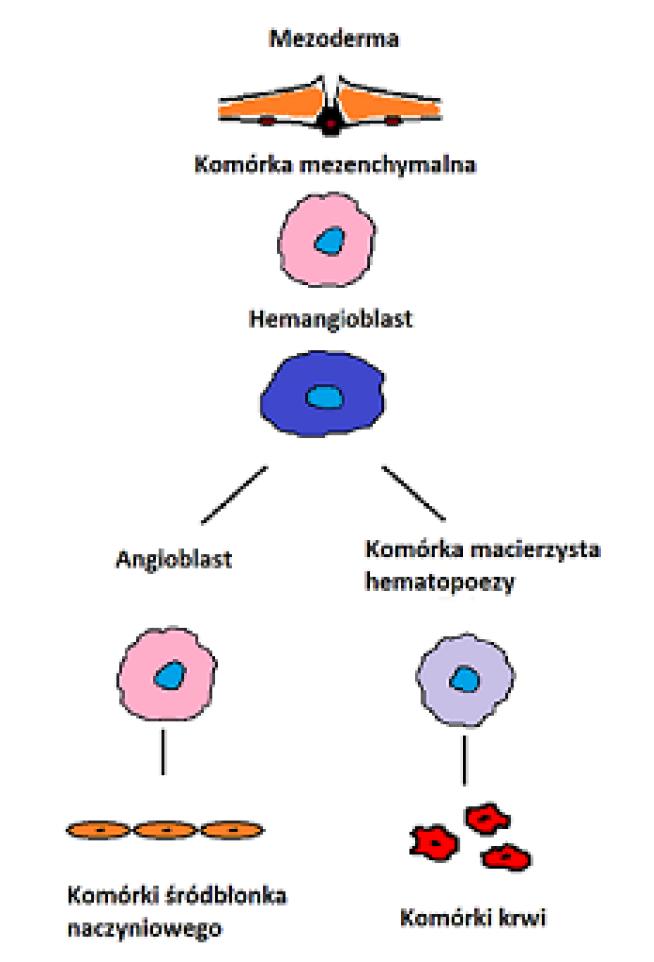
Ryc. 2. Różnicowanie się komórek pochodzenia mezodermalnego oraz wspólne pochodzenie i rozwój komórek układu naczyniowego i hematopoetycznego
Na podstawie profilu molekularnego i fenotypowania zidentyfikowano populację komórek bipotencjalnych w okresie postnatalnym. Ludzkie komórki o charakterze bipotencjalnym zostały wyizolowane z krwi pępowinowej UCB w oparciu o antygeny CD34+ oraz CD34+, CD309+, z komórek szpiku kostnego z ekspresją CD133+ oraz mobilizowanych komórek jednojądrzastych krwi obwodowej PBMC o fenotypie CD133+. Wszystkie wymienione populacje komórkowe poddane odpowiednim czynnikom różnicują się do komórek hematopoetycznych i komórek śródbłonków naczyniowych [11,55].
Mezenchymalne komórki macierzyste
Mezenchymalne komórki macierzyste (mesenchymal stem cell/MSC) i ich potencjał biologiczny opisano na początku lat 70 ub.w. w szpiku kostnym. Według Międzynarodowego Towarzystwa Terapii Komórkowej ISCT, MSC definiuje się, jako komórki z ekspresją SH-4/ CD73+, Thy-1/CD90+, SH-2/CD105+ [1,4,8,19]. Dopuszcza się, że w obrębie subpopulacji MSC może się pojawiać ekspresja innych markerów, takich jak: CD44+, c-kit/ CD117+, CD166+, Stro-1+, CD10+, CD13+, CD49+, CD146+, oraz CD271+ [4,23]. Uważa się, że obecność markera CD271, najlepiej definiuje MSC pochodzenia szpikowego, natomiast ekspresja CD146 może być pomocna w identyfikacji pozaszpikowych komórek MSC [19,21]. Wykazano również, że wczesne, niepoddane różnicowaniu MSC, wykazują ekspresję CD166. Klasyczna linia MSC nie powinna wykazywać ekspresji markerów związanych z linią hematopoetyczną: CD11b, CD14, CD19, CD79a, CD33, CD34, CD45, CD133, CD309, HLA-DR, czy też markera komórek śródbłonka CD31. Ilościowo MSC stanowią 0,01-0,0001% jednojądrzastych komórek szpiku kostnego, a ich poziom obniża się wraz z wiekiem [4,23]. Komórki MSC w warunkach in vitro są adherentne, poddane działaniu odpowiednich czynników wzrostowych różnicują się do linii osteoblastów, adipocytów oraz chondrocytów. Hodowla komórek MSC w warunkach in vitro jest ograniczona i po 40 pasażach obserwuje się spadek aktywności telomerazy, potencjału proliferacyjnego i tendencji do zmian nowotworowych [3,4,6]. MSC opisano i zidentyfikowano poza szpikiem kostnym w obrębie nisz tkankowych skóry, kości, mięśni, płuc, wątroby, naczyń krwionośnych, w krwi pępowinowej oraz krwi obwodowej [4,8,23,73].
Komórki progenitorowe śródbłonków naczyniowych (endothelial progenitor cell/EPC)
Termin angiogenezy został po raz pierwszy zastosowany przez Folkmana, który opisał go jako formowanie się nowych naczyń krwionośnych z już istniejących w czasie obserwacji unaczynionych guzów nowotworowych [27]. Od czasu wprowadzenia definicji na początku lat 70 ub.w. postęp wiedzy znacząco wzrósł. Przełomowym odkryciem rzucającym nowe spojrzenie na mechanizmy angiogenezy były opublikowane przez Asahara i wsp. wyniki badań z 1997 r. Opisane przez badaczy C-EPC były potwierdzeniem stawianej hipotezy, że proces neowaskularyzacji nie jest ograniczony do życia embrionalnego [3]. Wykazano, że komórki macierzyste pochodzące z krwi obwodowej i szpiku kostnego w określonych warunkach mogą się różnicować i jako C-EPC pełnić funkcje regeneracyjne [56]. Asahara i wsp. pierwsi udowodnili, że komórki macierzyste o fenotypie CD34+, CD309+ izolowane z krwi obwodowej oraz stymulowane czynnikiem wzrostu śródbłonka naczyniowego VEGF mogą w warunkach in vitro różnicować się do komórek śródbłonka naczyniowego. W modelach zwierzęcych obserwowano udział komórek CD34+, CD309+ w formowaniu nowych naczyń, czyli tzw. neoangiogenezy. Opisane przez grupę badawczą Asahary komórki C-EPC występują w niewielkiej liczbie i stanowią zaledwie 0,01-0,0001% jednojądrzastych komórek krwi obwodowej PBMC, w przeciwieństwie do ich odpowiedników w szpiku kostnym, tzw. BM-EPC, opisanych niedługo po C-EPC, które stanowią 1-5% komórek zrębu szpiku kostnego [42].
Dotychczasowe wyniki badań nie pozwalają na precyzyjną definicję EPC. Immunofenotypowanie z wykorzystaniem markerów powierzchniowych pozwala na wyodrębnienie subpopulacji EPC, których właściwości biologiczne sprawdza się w testach klonogennych [9,57]. Uważa się, że komórki zdolne do tworzenia nowych naczyń krwionośnych niezależnie od cech immunofenotypowych należy definiować jako komórki EPC [37,57,66]. Szeroki profil markerów stwierdzanych na powierzchni komórek zróżnicowanych, nieznajomość markerów typowych dla komórek progenitorowych, odkrycie EPC w szpiku kostnym, w krwi pępowinowej, krwi obwodowej oraz w obrębie struktur nisz tkankowych innych narządów, czyni je populacją wyjątkowo heterogenną. Zmienność fenotypowa komórek EPC koreluje z możliwościami biologicznymi, które przejawiają się tworzeniem wyspecjalizowanych populacji komórkowych o cechach: regeneracyjnych, syntetyzujących czynniki wzrostowe, mobilizujących inne populacje komórkowe do wspomagania lokalnej angiogenezy oraz będących źródłem dla zróżnicowanych komórek śródbłonka [9,55].
Analiza subpopulacji EPC możliwa jest również dzięki metodom hodowli komórkowych, które pozwalają na ocenę morfologii, zdolności różnicowania i potencjału proliferacyjnego. Testy klonogenne EPC wykazały obecność tzw. wczesnych komórek EPC, które charakteryzują się zdolnością formowania kolonii CFU do 10 dni w hodowlach oraz komórek EPC określanych jako późne-EPC, które wykazują zdolność CFU nawet po około 4 tygodniach hodowli. Wczesne EPC wykazują zmniejszoną ekspresję CD133+ w stosunku do hemangioblastu. Ponadto wykazują ekspresję CD309+, CD34+ oraz markerów charakterystycznych dla komórek śródbłonka np: CD31, CD62, CD144/VE-kadheryna, eNOS, vWF oraz komórek hematopoetycznych CD14, CD45. Całkowita utrata ekspresji CD133 określa późne komórki EPC, a pojawienie się CD144 wyznacza etap różnicowania się do komórek śródbłonków naczyniowych. Wczesne komórki EPC wykazują silniejszy potencjał integracyjny z naczyniami krwionośnymi w testach in vivo, a późne EPC cechuje zwiększona tendencja do tworzenia struktur przypominających naczynia krwionośne w testach in vitro [64]. W obrębie EPC potwierdzono również ekspresję innych markerów, takich jak receptor czynnika wzrostu fibroblastów FGFR, CD38, CD117/c-kit, CXCR4, vWF, CD144/VE-kadheryna, CD202b/Tie-2/TEK, UEA-1 [9,37,42,55,58,66,72,82].
Profil markerów powierzchniowych ludzkich i mysich EPC jest zbliżony, ale nie na tyle zgodny, aby model zwierzęcy odzwierciedlał zjawiska zachodzące w organizmie ludzkim, co stanowi dodatkowe utrudnienie w ocenie, oznaczaniu EPC oraz badaniu ich biologicznych właściwości [82]. Wciąż aktualnym pozostaje więc pytanie: jaki jest rzeczywisty immunofenotyp subpopulacji EPC, który wyróżnia komórki odpowiedzialne za konkretne efekty biologiczne [9,82]?
Nisze komórek macierzystych i progenitorowych w biostrukturze ścian naczyń krwionośnych
Pojęcie niszy tkankowych z komórkami o charakterze macierzystym/progenitorowym najlepiej opisano na podstawie mikrosiedliska szpiku kostnego. Nisze tkankowe ze spoczynkowymi komórkami macierzystymi i progenitorowymi w obrębie zróżnicowanego środowiska tkankowego stanowią struktury macierzy międzykomórkowej ECM: zawarte w niej metaloproteinazy oraz czynniki wzrostu, populacje komórek, które kontrolują i regulują właściwości biologiczne niszy. Kontrola komórek macierzystych/progenitorowych zakotwiczonych w środowisku niszy może się odbywać za pomocą receptorów sygnalizacji międzykomórkowej, które definiują stopień dywersyfikacji, potencjał różnicowania, proliferacji oraz mobilizacji [29,41,48,59]. Przedstawione wyżej przykłady środowiska i regulacji funkcjonowania niszy tkankowych są najczęściej prezentowanymi w podejmowanych próbach definicji niszy tkankowej. Definicja niszy tkankowej jest trudna do sprecyzowania, gdyż charakter, profil sposobów regulacji funkcjonowania niszy może zależeć również od lokalizacji tkankowej i narządowej [49]. Prowadzone są badania, które mają na celu wyjaśnienie komunikacji między mobilizacją komórek z jam szpikowych, a środowiskiem tkankowym np.: rola czynników wzrostowych i ich receptorów w procesach adhezji i deadhezji, chemotaksji i mobilizacji [9,28]. Czynnik pochodzenia stromalnego typu 1, SDF-1/CXCL12 i jego receptor CXCR4/CD184 są modelowym przykładem procesu oddziaływań i mobilizacji komórek macierzystych i progenitorowych ze środowiska szpiku kostnego do środowiska tkankowego [7,28]. Wykazano, że ekspresja SDF-1 w tunica media oraz tunica adventitia na zasadzie chemotaksji powoduje mobilizację komórek macierzystych i progenitorowych z jam szpikowych i promuje hiperplazję w układzie naczyniowym. Aktywację szlaku CXCR-4-SDF-1 zaobserwowano również w przypadku przebudowy ściany naczyń krwionośnych płuc u pacjentów z nadciśnieniem tętniczym [52]. Aktywacja szlaku SDF-1-CXCR4 może być spowodowana lokalnym niedotlenieniem, czy też urazami mechanicznymi komórek śródbłonka naczyniowego. W procesie mobilizacji komórek macierzystych i progenitorowych z jam szpikowych do niszy tkankowych mogą być zaangażowane również inne czynniki wzrostowe, np. granulocytowo-makrofagowy czynnik wzrostu GM-CSF, czynnik wzrostu fibroblastów FGF lub VEGF [7,9,15,28].
Istnienie nisz z komórkami macierzystymi i progenitorowymi na różnych poziomach zróżnicowania zaobserwowano i opisano w obrębie wielu narządów. Obecność komponentu naczyniowego wydaje się integralnym składnikiem nisz tkankowych. Ostatnie badania wskazują, że również biostruktury, z których zbudowane są naczynia krwionośne tunica: intima, media i adventitia mają komórki macierzyste/progeniterowe ścian naczyń (vascular wall-stem cell/progenitor cell) VW-SC/PC zakotwiczone w niszach tkankowych. Tilki i wsp. zaproponowali definicje VW-SC/PC, jako komórek, które poddane odpowiednim czynnikom różnicującym stają się źródłem komórek śródbłonka, komórek mięśniówki gładkiej SMC oraz fibroblastów, czyli głównych składników biostruktury naczynia [81]. Strefa okołośródbłonkowa na granicy tunica intima i media oraz waskulogenna (vasculogenic zone) zlokalizowane na granicy tunica media i adventitia to obszary występowania niszy komórek VW-SC/PC o dużym stopniu heterogenności (ryc.3). W obrębie biostruktur naczyniowych wykazano obecność niszy z komórkami progenitorowymi mięśni gładkich SMP, komórkami macierzystymi hematopoezy HSC, komórkami MSC, nerwowymi komórkami macierzystymi NSC, komórkami mezangium, komórkami linii rozrodczej oraz z komórkami macierzystymi odpowiedzialnymi za inicjowanie procesów nowotworowych [81].

Ryc. 3. Schemat trójwarstwowej biostruktury naczynia krwionośnego wraz z proponowanymi obszarami niszy ze spoczynkowymi komórkami macierzystymi i progenitorowymi: strefa okołośródbłonkowa oraz strefa waskulogenna. Uważa się, że wszystkie trzy biostruktury naczynia krwionośnego mogą zawierać niezróżnicowane komórki o charakterze macierzystymi lub progenitorowym
Komórki macierzyste/progenitorowe w tunica intima
Tunica intima stanowi wewnętrzną warstwę naczynia żylnego i tętniczego. Jest to pojedyncza warstwa komórek śródbłonka osadzona na błonie podstawnej i wsparta składnikami ECM, która bezpośrednio kontaktuje się ze światłem naczynia. Tunica intima uważana była przez długi czas za w pełni zróżnicowaną biostrukturę, mechaniczną barierę, która oddziela przepływającą krew od lokalnych tkanek. Obecnie tunica intima przedstawia się zupełnie inaczej.
Obserwacje Schwartza i Beneditta dostarczyły informacji o zdolności różnicowania i proliferacji komórek śródbłonków naczyń krwionośnych, migracji i regeneracji w chwili miejscowego uszkodzenia. Trzy lata później ta sama grupa wykazała, że komórki śródbłonka naczyń krwionośnych poddane bodźcom stymulującym lub imitującym zranienie ulegają wzmożonej aktywacji. Zaobserwowano, że w obrębie tunica intima występują klastry komórek o podwyższonym potencjale proliferacji i wzrostu, opisywane w literaturze jako: high turnover regions lub growth centers [63,64,65].
Ingram i wsp. uważają, że regeneracyjna aktywność tunica intima, pod wpływem czynnika uszkadzającego, jest rezultatem obecnych w niej klastrów VW-SC/PC. Grupa Ingrama wykazała, że komórki śródbłonka wywodzące się z ludzkiej żyły pępowinowej, HUVEC oraz ludzkiej aorty, HAEC wykazują właściwości komórek EPC. Komórki śródbłonka po izolacji z naczyń poddawano analizie za pomocą różnicujących testów proliferacyjnych i klonogennych, a następnie porównano z właściwościami EPC wyizolowanymi z krwi pępowinowej. Potencjał proliferacyjny pojedynczych komórek HUVEC, HAEC oraz EPC krwi pępowinowej sięgał odpowiednio 52, 53 i 55%. Komórki EPC krwi pępowinowej wykazywały natomiast znacznie większy potencjał tworzenia kolonii CFU (47%) w stosunku do HUVEC (28%) oraz HAEC (27%). Fenotypowanie śródbłonków wykazało profil markerów: CD31+, CD105+, CD141+, CD144+, CD146+, vWF oraz CD309+ typowy dla EPC. Komórki nie wykazywały natomiast ekspresji markerów hematopoetycznych: CD14–, CD45 – [34].
Grupa Fanga i wsp. dostarczyła kolejnych dowodów potwierdzających potencjał regeneracyjny obecny w tunica intima. Badacze wykazali, że intima ściany naczynia u myszy szczepu C57BL/6J zawiera klastry komórkowe, które opisali, jako dorosłe komórki macierzyste śródbłonków naczyniowych VESC. Komórki VESC zostały opisane przez grupę badawczą w śródbłonkach żylnych i tętniczych nerki, śledziony oraz płuc. Do celów badawczych wyizolowano śródbłonek ze ściany naczyń płuc myszy C57BL/6J. Izolacji dokonano na podstawie markerów Lin–, CD31+, CD105+, Sca-1+, CD117+. W celu wykluczenia komórek linii hematopoetycznej, komórki oczyszczono z komórek CD45+. Komórki po enzymatycznej izolacji poddano hodowli z dodatkiem VEGF. Badacze obserwowali jednostki CFU oraz ekspresję markerów śródbłonkowych: CD31+, CD105+, CD144+, vWF, ale brak ekspresji markerów hematopoetycznych, takich jak CD45–. Komórki o dużej aktywności CFU, zwłaszcza śródbłonki CD117+ wyznakowano barwnikiem fluorescencyjnym GFP. Komórki poddano analizie na obecność innych markerów śródbłonka. Wykazano obecność: CD144+, CD106+, CD309+, VEGFR-1, CD104+ oraz CD34+. Komórki po około 14 dniach hodowli znakowano GFP i przeszczepiono do szczepu dzikiego C57BL/6J-wt, bez GFP. Dodatkowo podzielono je na populację komórek CD117+ i CD117–. Okazało się, że komórki CD117+ były obecne we krwi, a nawet wykazywały zdolność inkorporacji do funkcjonujących już naczyń. Komórki CD117– nie wykazywały już takich właściwości. Na podstawie zgromadzonych wyników badacze postawili tezę, że w obrębie zróżnicowanych śródbłonków ściany naczynia znajduje się bardzo mała liczba komórek o charakterze komórek macierzystych i progenitorowych, tzw. VESC zdolnych do inicjowania angiogenezy oraz inkorporacji do funkcjonujących już naczyń w środowisku in vivo [26].
Grupa Fanga i wsp. w 2005 roku przedstawiła wyniki badań, w których prezentuje obecność komórek CD105+, CD31–, CD34–, CD11a–, CD11b–, CD45– izolowanych ze ściany aorty płodowej. Komórki w warunkach hodowli in vitro z dodatkiem VEGF różnicowały się do komórek śródbłonka. Pod wpływem medium różnicującego komórki po 3 tygodniach hodowli wykazały ekspresję CD31+, CD202b+ oraz vWF, co potwierdzono w analizie FACS oraz western-blotting. Podanie komórek do mysich modeli uszkodzenia skóry inicjowało procesy angiogenne, a same komórki obserwowano w naczyniach, co potwierdzono metodą immunohistochemiczną na obecność markerów śródbłonka: CD31, vWF oraz mięśniówki: α-SMC [25].
Powyższe przykłady dowodzą, że obserwowana na początku lat 70 ub.w. zdolność zróżnicowanych komórek tunica intima do regeneracji, proliferacji i migracji nie wynika wyłącznie z biologicznych właściwości śródbłonków. Okazuje się, że zróżnicowany śródbłonek zawiera niewielki odsetek komórek o charakterze macierzystym i progenitorowym, które stymulowane odpowiednim środowiskiem czynników wzrostowych wykazuje zdolności proliferacji i regeneracji w testach in vitro i in vivo.
Komórki macierzyste/progenitorowe w tunica media
Tunica media, zbudowana z komórek mięśni gładkich, SMC, stanowi o sile skurczowej i rozkurczowej naczynia, nadaje i reguluje ton przepływu krwi, uczestniczy w kontrolowaniu stabilności mechanicznej. W układzie tętniczym jest ona silniej rozwinięta niż w żylnym. Istnieje hipoteza, według której komórki SMC tunica media pod wpływem zmian naczyniowych wywołanych procesem chorobowym ulegają różnicowaniu z postaci spoczynkowej do aktywnej proliferacyjnej. Uważa się, że proces ten może wynikać z aktywności komórek progenitorowych mięśni gładkich PMC, które uczestniczą w procesie proliferacji i migracji komórek z tunica media do strefy okołośródbłonkowej tunica intima. Aktywność komórek PMC może odpowiadać za formowanie się zmian naczyniowych np. powstawanie neointimy [30,47,51].
Kolejne dowody potwierdzające istnienie VW-SC/PC pochodzą z eksperymentów wykonanych na segmentach ścian naczyń izolowanych z tętnicy łuku aorty, tętnicy piersiowej oraz udowej. Eksperymentalnie udowodniono, że komórki izolowane ze ścian wspomnianych naczyń wykazują ekspresję markerów Oct-4, Stro-1, Sca-1 oraz Notch-1, charakterystycznych dla MSC. Fenotypowanie komórek nie wykazało obecności markerów komórek hematopoetycznych oraz śródbłonków naczyniowych. W testach in vitro komórki poddane czynnikom różnicującym rozwijały się do linii komórek SMC, adipocytów oraz chondrocytów [47,51].
W cytowanej wcześniej pracy Fang i wsp. przedstawiają, że komórki CD309+, CD105+, CD34– izolowane ze ściany ludzkiej aorty płodowej oprócz potencjału proangiogennego i zdolności tworzenia dojrzałych komórek śródbłonków, mają możliwość różnicowania się do komórek SMC. Komórki po enzymatycznym trawieniu ściany naczynia i hodowli poddano fenotypowaniu. Analiza nie wykazała markerów charakterystycznych dla mięśniówki: α-SMA, MHC I oraz kalponiny. Natomiast hodowla komórek w medium różnicującym z dodatkiem PDGF-BB powodowała ekspresję markerów mięśniówki u 80% komórek. Komórki wykazywały ekspresję czynnika vWF oraz α-SMA, co nadawało im mieszany charakter mioangiogenny. Podanie komórek do mysiego modelu eksperymentalnego poprzez żyłę ogonową wzmagało proces neoangiogenezy i gojenia ran w skórze. Na podstawie tego zjawiska można stwierdzić, że poza właściwościami angiogennymi komórki doskonale radziły sobie z zasiedlaniem miejsc ich przeznaczenia [25].
Komórki izolowane z tunica intima żyły odpiszczelowej pacjentów z żylakami, na granicy tunica media i adventitia, czyli tzw. strefy waskulogennej zawierają komórki VW-SC/PC z ekspresją CD34+ oraz CD117+. Komórki poddane hodowli, a następnie fenotypowaniu wykazały ekspresję markerów MSC: CD44+, CD90–, CD105+, natomiast nie wykazano u nich ekspresji markerów linii hematopoetycznej. W testach klonogennych komórki tworzą CFU po 2-3 tygodniach hodowli, a poddane czynnikom różnicującym rozwijały się w linie osteoklastów, osteoblastów i chondroblastów. W środowisku różnicującym obserwowano tworzenie struktur naczyniopodobnych, a także ekspresję genu komórek MSC, Oct-4 [18,83].
Podsumowując, VW-SC/PC obecne w tunica media są komórkami, które mają właściwości proangiogenne, charakter komórek MSC, a także mają możliwość różnicowania się w komórki mięśni gładkich oraz inne populacje komórkowe. Zaobserwowano również, że w obrębie tunica media występują komórki, które nie wykazują cech linii hematopoetycznej [83].
Komórki macierzyste/progenitorowe w tunica adventitia
Tunica adventitia jest najbardziej zewnętrzną warstwą naczynia krwionośnego, bogatą w fibroblasty, kolagen i system okołonaczyniowych włókien nerwowych o nieuporządkowanej strukturze. Podobnie jak w przypadku pozostałych biostruktur naczynia, tunica adventitia uważana była za barierę oddzielającą naczynie od lokalnego środowiska tkankowego, w czym nie dopatrywano się szczególnego znaczenia [31]. Poznanie molekularnych i genetycznych mechanizmów odpowiedzialnych za jej formowanie pozwoliło odkryć jej dynamizm oraz zaangażowanie w różne procesy biologiczne, takie jak: udział w rekonstrukcji naczyniowej, pośrednictwo w sygnalizacji między komponentami naczynia, a lokalnym środowiskiem tkankowym, transport komórek układu immunologicznego do środowiska tkankowego, czy też kontrola i nadawanie tonu naczyniowego przez oddziaływanie na tunica media [31,32]. Bezpośredni kontakt tunica adventitia z lokalnym środowiskiem tkankowym powoduje, że sygnały o zmianach patologicznych zachodzących w świetle naczynia oraz w tunica intima i media mogą być przesyłane za pomocą jej komponentów do pozanaczyniowego środowiska tkankowego. Tunica adventitia może uczestniczyć w regeneracji lokalnego środowiska tkankowego na wczesnych etapach rozwoju choroby, a na poziomie bardziej zaawansowanym w tworzeniu neointimy [31,32,49,50].
Na podstawie obecnych doniesień uważa się, że tunica adventitia jest największym rezerwuarem VW-SC/PC. W 2004 r. Hu i wsp. opisali, że tunica adventitia aorty szczepu myszy Apo E – /- ma komórki z profilem Sca-1+, CD34+, CD117+, CD309+. Komórki te poddane działaniu czynnika PDGF-BB ulegają różnicowaniu do SMC w warunkach in vitro. Populacja fibroblastów Sca-1+, inkorporowana do eksperymentalnych przeszczepów naczyń myszy Rosa26 migrowała przez tunica adventitia do tunica media. Komórki traciły ekspresję Sca-1, a nabywały fenotyp SMC. Po 4 tygodniach od przeszczepu obserwowano ich obecność również w neointimie, gdzie promowały jej progresję [33,82].
Sainz i wsp. wyizolowali populację komórek progenitorowych z aorty myszy ApoE – / – o profilu markerów Sca-1+, CD117-/low, Lin–, CD34-/low. Komórki te poddane czynnikom, takim jak: VEGF lub PDGF-BB/TGF-β1 ulegały różnicowaniu do komórek śródbłonka oraz SMC, ale nie do HSC. W celu obserwacji zachowania komórek Sca-1+ w warunkach in vivo, komórki podawano myszom, u których fibroblasty ściany aorty nie wykazywały ekspresji Sca-1. Po 4 tygodniach od transplantacji obserwowano obecność komórek Sca-1+, których udział w formowaniu się neointimy stanowił do 30% [62].
Zaobserwowano również, że migracja komórek z tunica adventitia i ich udział w formowaniu neointimy może być regulowana poprzez sygnalizację sonic-hedgehog Shh, która zaangażowana jest w tworzenie pierwotnych struktur naczyniowych. Passman i wsp. opisali populacje z ekspresją Sca-1+, CD309+, CD34+, CD117–, CD140b+, zlokalizowane w subregionie adventitia. U myszy transgenicznych Shh-/- zaobserwowano znacznie mniej komórek Sca-1+, co oznacza, że sygnalizacja Shh może być zaangażowana w regulację procesu zasiedlania komórek Sca-1+ do tunica media, gdzie przyczyniły się do formowania neointimy [57].
Uważa się, że największy zasób VW-SC/PC znajduje się w strefie waskulogennej na granicy tunica media i adventitia, również w obrębie samej tunica adventitia. Zengin i wsp. po raz pierwszy wprowadzili termin tzw. strefy waskulogennej w oparciu o badania ludzkich ścian naczyń żylnych i tętniczych. Ze ściany naczynia poddanej enzymatycznemu trawieniu wyizolowano komórki CD34+ oraz wysortowano na podstawie markera CD105 w celu wykluczenia zróżnicowanych komórek śródbłonka. Następnie wykonano modyfikację tętniczego testu pierścieniowego na bazie kolagenu typu I w celu określenia potencjału angiogennego komórek. Zaobserwowano, że po 3-4 dniach hodowli dochodzi do formowania się pierwszych struktur o charakterze naczyniowym. Po około 14 dniach hodowli komórki wykazywały ekspresję antygenów CD34+, CD309+, CD202b+. Następnie obserwowano ekspresję CD144, białka okludyny, charakterystycznego dla ścisłych połączeń komórkowych oraz CD66a/CEACAM1 występującego na aktywowanych komórkach śródbłonków naczyniowych. Potencjał angiogenny komórek na różnych etapach testu potwierdzono poprzez hodowlę in vitro, gdzie z dodatkiem czynnika VEGF tworzyły system mikronaczyń po transplantacji do mysich modelów nowotworowych. Komórki także opisano w naczyniach żylnych i tętniczych z prostaty, śledziony, serca, mózgu oraz nerki [88].
Campagnolo i wsp. w żyle odpiszczelowej wykazują obecność komórek CD34+, CD31–, które opisują jako komórki progenitorowe wywodzące się z żyły odpiszczelowej SVP obecne w obrębie tunica adventitia i vasa vasorum. Po enzymatycznym trawieniu ściany naczynia, inkubacji z medium różnicującym wyhodowano wyselekcjonowane komórki, a następnie poddano analizie FACS. Komórki CD34+, CD31–, vWF w obecności medium nabywają fenotypu komórek mezenchymalnych/pericytów (ekspresja genu Sox-2), a w testach klonogennych wykazują silny potencjał proliferacyjny, a także zdolność wieloliniowego różnicowania się. Na powierzchni komórek SVP wykazano również jednoczesną ekspresję markerów komórek MSC , CD117+ oraz EPC, CD133+. Komórki w testach in vitro wykazują zdolność różnicowania się do komórek śródbłonków naczyniowych i SMC. SVP okazały się również źródłem czynników wzrostowych. Komórki SVP podane do mysich modeli transplantacyjnych wzmagają neowaskularyzację u zwierząt przez oddziaływanie na inne populacje komórek spoczynkowych niszy. Zaobserwowano, że podanie inhibitorów receptorów Tie-2 oraz PDGFR-β, które są odpowiedzialne również za udział w kapilarogenezie i kształtowaniu biostruktury naczynia koreluje ze znacznym obniżeniem potencjału migracyjnego i neowaskularyzacyjnego w porównaniu do grupy zwierząt, którym domięśniowe podanie komórek inicjowało neowaskularyzację w mysich modelach niedokrwiennych. Obecność komórek SVP w biostrukturze naczynia potwierdzano nawet 14 dni po przeprowadzonych transplantacjach [10].
Podobne rezultaty zaobserwował zespół Fanga, który izolował komórki CD309+, CD105+, CD34 – z ludzkiego materiału tętniczego. Komórki po izolacji hodowane w obecności VEGF oraz PDGF po inkorporacji do zwierzęcych modelów wzmagają progresje procesu neowaskularyzacji i różnicowanie się do komórek śródbłonka oraz SMC. Komórki różnicują się również do linii adipocytów, czy osteoblastów [25].
Corselli i wsp. na materiale ze stromalnej frakcji naczyniowej wykazali, ze tunica adventitia naczyń żylnych i tętniczych zawiera komórki CD34+, CD31–, CD146–, CD45–, które wykazują cechy MSC. Komórki nie wykazują ekspresji markerów komórek śródbłonka, perycytów oraz hematopoetycynych, ale mają markery klasycznych MSC: CD44+, CD73+, CD90+ i CD105+. Ich inkorporacja w zwierzęcych modelach doświadczalnych koreluje dodatnio ze wzmożoną aktywnością regeneracyjną naczyń [16,17].
Z kolei zespół Kleina opisuje w tętnicach ludzkich mezenchymalne, multipotencjalne komórki macierzyste ściany naczynia VW-MPSC z obszaru strefy waskulogennej oraz tunica adventitia. Komórki VW-MSCP wykazują ekspresję klasycznych markerów komórek MSC: CD44+, CD90+, CD74+ oraz brak markerów CD34–, CD45–. Komórki VW-MPSC mają zdolność różnicowania się do perycytów oraz SMC, co według badaczy może być rezerwuarem komórek np. do rekonstrukcji ściany naczynia. Identyfikacja VW-MPSC została dodatkowo potwierdzona poprzez barwienie immunohistochemiczne fragmentów ściany tętnicy na obecność α-SMA oraz kolokalizację z markerem MSC, CD73 oraz RGS5, markerem aktywowanych perycytów. W obrębie strefy waskulogennej zaobserwowano również ekspresję genu Sox-2 oraz CD90+, nestyny i CD44+. W kolejnym etapie wykonano tętnicze testy pierścieniowe, które po dwóch dniach wykazały obecność markera perycytów NG2+, CD90+, ale brak ekspresji markerów śródbłonkowych: CD34 i CD146. Podanie komórek CD44+ in vitro oraz in vivo do mysich modeli SCID wzmagało tworzenie struktur naczyniowych. Przypuszcza się, że komórki VW-MPSC CD44+ mogą być zaangażowane w proces rekonstrukcji i utrzymywania prawidłowej homeostazy ściany naczynia, co wynika np. ze zdolności różnicowania się do SMC i perycytów oraz wbudowywania się komórek do biostruktury naczynia [40].
Badania eksperymentalne na modelach zwierzęcych oraz wyniki prac klinicznych wskazują, że w obrębie tunica adventitia mogą się znajdować nisze tkankowe z populacjami komórek macierzystych/progenitorowych o silnym potencjale angiogennym. Interesujące jest to, że aktywność wspominanych komórek może być zaangażowana w proces tworzenia systemu mikroskopijnego unaczynienia, obserwowany u pacjentów z miażdżycą. Wzmożona neoangiogeneza oraz gęstość mikrounaczynienia w tunica adventita wydaje się korelować z postępem miażdżycy i formowaniem neointimy [53,84].
Nisze tkankowe tunica adventitia z komórkami VW-SC/ PC odgrywają ważną rolę w procesie prawidłowej homeostazy naczynia i w utrzymywaniu kontaktu z lokalnym środowiskiem tkankowym. Są źródłem komórek macierzystych i progenitorowych na różnych poziomach rozwoju, co potwierdzono poprzez opisanie heterogennej „strefy waskulogennej”. Wstępne informacje dowodzą, że tunica adventitia może być zaangażowana w rozwoju chorób naczyniowych, a jednym z powodów może być wzmożona aktywność proliferacyjna komórek VW-SC/ PC [31].
Perycyty
Perycyty/komórki ścienne/komórki Rougeta definiowane są najczęściej jako okołośródbłonkowe komórki, które opisano począwszy od mikronaczyń i kapilar naczyniowych, aż po duże naczynia krwionośne. Komórki te umiejscowione są wzdłuż komórek śródbłonka, z którymi dzielą wspólnie błonę podstawną oraz połączone są licznymi połączeniami międzykomórkowymi. W większych naczyniach mogą występować w obrębie tunica adventitia, a ich charakter morfologiczny i fizjologiczny jest swoisty narządowo [6]. Perycyty to źródło czynników wzrostowych, które regulują proces formowania „tuby naczyniowej”, nadają kształt i światło naczyniu, regulują jego ton, oddziaływają na komórki śródbłonka regulując ich proliferację i migrację. W dużych naczyniach perycyty spotykane są również w tunica media i tunica adventitia. Perycyty są źródłem czynników wzrostowych, które regulują angiogenezę oraz utrzymują homeostazę naczyniową i uczestniczą w syntezie składników ECM. Sekrecja przez perycyty czynnika wzrostu pochodzenia płytkowego typu B PDGF-B kontroluje proces tzw. kapilarogenezy, czyli tworzenia splotów naczyniowych przez wbudowywanie komórek śródbłonka do biostruktury naczynia. Synteza PDGF-B przez perycyty oddziałuje również na różnicowanie, proliferację i migrację EPC. Myszy transgeniczne pozbawione perycytów wykazują liczne anomalie naczyniowe, np. nieregularną i chaotyczną strukturę naczyniową, czy też zmiany w połączeniach międzykomórkowych [6,15]. Perycyty są komórkami zdecydowanie wyróżniającymi się fenotypowo na tle całej biostruktury naczynia, gdyż nie wykazują ekspresji markerów komórek śródbłonka i hematopoetycznych, takich jak CD31, CD34, CD144, czynnik vWF, UEA1 oraz CD45. Natomiast na ich powierzchni obserwuje się ekspresję CD146, NG2, PDGF-B, fosfatazę alkaliczną oraz ludzki antygen dermalny 1 HDA1 [16,22].
Biorąc pod uwagę zaangażowanie perycytów w kapilarogenezę, regulację śródbłonków i innych komórek biostruktury naczynia poprzez sekrecję czynników wzrostowych, utrzymywanie prawidłowej homeostazy ściany naczynia czyni je potencjalnym źródłem komórek o właściwościach regeneracyjnych. W literaturze naukowej perycyty oraz komórki blisko z nimi spokrewnione, wykazujące właściwości regeneracyjne określane są komórkami perywaskularnymi PVC lub nazywane są potomkami komórek MSC okresu postnatalnego [16,17].
Komórki PVC opisano oraz analizowano w naczyniach: mięśni szkieletowych, łożyska, skóry, mózgu, krwi pępowinowej, serca, miazgi zęba i szpiku kostnego [22]. Komórki poddane hodowli in vitro, a następnie fenotypowaniu wykazywały ekspresję markerów MSC: CD44+, CD90+, CD73+, CD105+ oraz właściwości miogenne [20,21,22]. PVC izolowane ze ścian naczyń, hodowane w warunkach in vitro ulegają również różnicowaniu do linii komórek osteoblastów, adipocytów, chondrocytów, a także mioblastów oraz EPC [14,16,17,19,20,21,22].
W mysim modelu doświadczalnym zaobserwowano, że podanie komórek PVC CD146+, CD34–, CD45–, CD56 izolowanych z mięśni o charakterze mioendotelialnym, przyczynia się do regeneracji miokardium oraz wzmaga neoangiogenezę u zwierząt [16]. Obecność PVC opisano również we krwi pępowinowej, w obrębie galarety Warthona. Komórki PVC izolowane z tego obszaru wykazują ekspresję markerów MSC, ale również ESC: Oct-4, SSEA- 4. Ich podanie w modelach eksperymentalnych powodowało migrację do miejsc uszkodzenia, sekrecję VEGF, czynnika wzrostu keratynocytów KGF oraz inicjowało procesy rekonstrukcyjne uszkodzonych mięśni, a także wzmagało proces neoangiogenezy [16,17].
Z kolei zespół badawczy Tiggesa przedstawia badania, z których wynika, że komórki PVC oraz MSC izolowane z tunica adventitia tętnicy udowej myszy mogą być zaangażowane w proces restenozy, który występuje po zabiegach angioplastyki tętnic. Mechanizm i zaangażowanie konkretnych populacji komórek w tym procesie jest wciąż nie do końca poznany. Badacze zaobserwowali wzmożoną aktywności komórek PVC CD146+, NG2+, PDGFβ-R+ w mysim modelu, w którym wywołano uszkodzenie ściany naczynia w porównaniu do myszy grupy kontrolnej. Część komórek wykazywała charakter MSC (CD29,CD90) i była obserwowana w neointima. Kultury in vitro oraz eksperymenty transplantacyjne in vivo wykazały, że komórki nie są pochodzenia szpikowego, lecz w wyniku wzmożonej proliferacji powstają w środowisku biostruktury tunica adventitia. Sytuacja ta jest podyktowana miejscową reakcją na uszkodzenie ściany naczynia [80].
Wydaje się, że połączenie fenotypu MSC, zdolność sekrecji wielu czynników wzrostowych, udział w homeostazie ściany naczynia czynią komórki PVC oraz same perycyty atrakcyjnym, potencjalnym źródłem komórek regeneracyjnych. Możliwości różnicowania się komórek PVC w komórki linii, takich jak osteoklasty, osteoblasty, adipocyty oraz komórki mięśni gładkich dodatkowo wzmacnia ich pozycję, jako atrakcyjnego czynnika regeneracyjnego.
Vasa vasorum, czyli unaczynienie ściany naczyń krwionośnych
Vasa vasorum to system mikroskopijnego unaczynienia występujący przede wszystkim w obrębie tunica adventitia dużych naczyń krwionośnych o średnicy powyżej 5 mm i grubości około 29 komórek. Przebieg vasa vasorum w obrębie ściany naczyń żylnych i tętniczych jest nieuporządkowany i może ulegać drobnym modyfikacjom w różnych naczyniach. Wyróżnia się vasa vasorum: interna, externa i vasa vasorae. Mikronaczynia vasa vasorum penetrują ścianę naczynia od tunica media przez tunica adventitia, w niektórych miejscach kontaktując się z lokalnym otoczeniem odpowiadając za kontakt tkankowy, stopień perfuzji naczyniowej, ale przede wszystkim za dostarczanie składników odżywczych i tlenu do najbardziej na zewnątrz usytuowanych komponentów tkankowych naczynia [61]. Istotność vasa vasorum w homeostazie i strukturze naczyń krwionośnych jest niekwestionowana, czego najlepszym dowodem są eksperymenty, w których usuwa się tę biostrukturę. Usunięcie vasa vasorum w modelach doświadczalnych zwierząt powoduje nekrozę tunica media, zwłaszcza jej środkową i zewnętrzną część, co wskazywałoby, że biostruktury naczynia umiejscowione najbardziej na zewnątrz zaopatrywane są przez vasa vasorum i komponenty tkankowe adventitia [49]. Przypuszcza się, że vasa vasorum może również uczestniczyć w procesie miażdżycowym, jednak informacje te wymagają dodatkowego potwierdzenia [5]. W obrębie vasa vasorum odnotowano aktywność komórek o charakterze macierzystym i progenitorowym. Z vasa vasorum żyły odpiszczelowej wyizolowano komórki z ekspresją markerów: CD34+, Sox2+, CD31-. W obrębie tej populacji wykazano również obecność komórek o charakterze MSC z ekspresja NG2+, PDGFRb+. Obie populacje poddawane czynnikom różnicującym wykazywały zdolności klonogenne oraz przejawiały cechy MSC, a inkorporowane do zwierzęcych modeli doświadczalnych inicjowały mechanizmy regeneracyjne [10,11].
Podsumowanie
Współczesne badania niszy z komórkami macierzystymi i progenitorowymi w obrębie zróżnicowanego środowiska tkankowego dostarczają informacji, z których wynika, że aktywność komórek macierzystych i progenitorowych w mniejszym stopniu występuje również w okresie życia postnatalnego. Biostruktury naczyń żylnych i tętniczych tunica: intima, media i adventitia są rezerwuarem VW-SC/PC, ponieważ czynniki środowiskowe, takie jak np. hemodynamika krwi wymuszają nieustanną kontrolę oraz regenerację ściany naczynia w chwili uszkodzenia. Granica między funkcją naprawczą, a udziałem w promowaniu zmian naczyniowych przez VW-SC/PC jest niejednoznaczna i trudna do sprecyzowania. Najprawdopodobniej aktywność naprawcza komórek VW-SC/PC przejawia się na bardzo wczesnych etapach powstawania choroby. Natomiast dane eksperymentalne z testów in vitro i na modelach zwierzęcych in vivo wykazują, że komórki te na etapie zaawansowanej choroby naczyniowej zamiast regenerować, mogą uczestniczyć w progresji choroby. Informacje te wskazują, że niezbędne jest dokładne poznanie biologicznych właściwości nisz z komórkami WV-SC/PC. VW-SC/ PC stanowią bogate źródło heterogennych populacji komórek, które poddane odpowiednim warunkom hodowlanym mogą się stać źródłem zróżnicowanych komórek biostruktury naczyniowej, a nawet komórek pochodzenia pozanaczyniowego. Cechy te predysponują do wykorzystania tych komórek w naczyniowej bioinżynierii tkankowej, dziedzinie, której możliwości terapeutyczne mogą być wykorzystywane w sytuacjach, w których tradycyjne metody chirurgii rewaskularyzacyjnej są niewystarczające.
Podsumowując, nisze komórek VW-SC/PC wydają się interesującym źródłem pozyskiwania komórek macierzystych do celów terapeutycznych, a ich izolacja, hodowla i różnicowanie w odpowiednim kierunku może się stać w niedługiej przyszłości alternatywą dla przeszczepów komórek macierzystych hematopoezy HSCT.
PIŚMIENNICTWO
[1] Abedin M., Tintut Y., Demer L.L.: Mesenchymal stem cells and the artery wall. Circ. Res., 2004; 95: 671-676
[PubMed] [Full Text HTML] [Full Text PDF]
[2] Anwar M.A., Shalhoub J., Lim C.S., Gohel M.S., Davies A.H.: The effect of pressure-induced mechanical stretch on vascular wall differential gene expression. J. Vasc. Res., 2012; 49: 463-478
[PubMed] [Full Text HTML] [Full Text PDF]
[3] Asahara T., Murohara T., Sullivan A., Silver M., van der Zee R., Li T., Witzenbichler B., Schatteman G., Isner J.M.: Isolation of putative progenitor endothelial cells for angiogenesis. Science, 1997; 275: 964-967
[PubMed]
[4] Bajek A., Olkowska J., Drewa T.: Mezenchymalne komórki macierzyste narzędziem terapeutycznym w regeneracji tkanek i narządów. Postępy Hig. Med. Dośw., 2011; 65: 124-132
[PubMed] [Full Text HTML] [Full Text PDF]
[5] Barger A.C., Beeuwkes R.3rd, Lainey L.L., Silverman K.J.: Hypothesis: vasa vasorum and neovascularization of human coronary arteries. A possible role of pathophsyilogy of atherosclerosis. N. Engl. J. Med., 1984; 310: 175-177
[PubMed]
[6] Bautch V.L.: Stem cells and the vasculature. Nat. Med., 2011; 17: 1437-1443
[PubMed]
[7] Bogunia-Kubik K., Gieryng A., Gebura K., Lange A.: Genetic variant of the G-CSF receptor gene is associated with lower mobilization potential and slower recovery of granulocytes after transplantation of autologous peripheral blood progenitor cells. Cytokine, 2012; 60: 463-467
[PubMed]
[8] Boxall S.A., Jones E.: Markers for characterization of bone marrow multipotential stromal cells. Stem Cell Int., 2012; 975871
[PubMed] [Full Text HTML] [Full Text PDF]
[9] Caiado F., Dias S.: Endothelial progenitor cells and integrins: adhesive needs. FibrogenesisTissue Repair, 2012; 5: 4
[PubMed] [Full Text HTML] [Full Text PDF]
[10] Campagnolo P., Cesselli D., Al Ha Zen A., Beltrami A.P., Krankel N., Katare R., Angelini G., Emanuel C., Madeddu P.: Human adult vena saphena contains perivascular, progenitor cells endowed with clonogenic and proangiogenic potential. Circulation, 2010; 121: 1735-1745
[PubMed] [Full Text HTML] [Full Text PDF]
[11] Campagnolo P., Wong M.M., Xu Q.: Progenitor cells in arteriosclerosis: good or bad guys? Antioxid. Redox. Signal, 2011; 15: 1013-1027
[PubMed]
[12] Cao N., Yao Z.X.: The hemangioblast: from concept to authentication. Anat. Rec., 2011; 294: 580-588
[PubMed] [Full Text HTML] [Full Text PDF]
[13] Chao H., Hirschi K.K.: Hemato-vascular origins of endothelial progenitor cells? Microvasc. Res., 2010; 79: 169-173
[PubMed] [Full Text HTML] [Full Text PDF]
[14] Chen C.W., Corselli M., Peault B., Huard J.: Human blood-vessel-derived stem cells for tissue repair and regeneration. J. Biomed. Biotechnol., 2012; 2012: 597439
[PubMed] [Full Text HTML] [Full Text PDF]
[15] Chen C.W., Montelatici E., Crisan M., Corselli M., Huard J., Lazzari L., Peault B.: Perivascular multi-lineage progenitor cells in human organs: regenerative units, cytokine sources or both? Cytokine Growth Factor Rev., 2009; 20: 429-434
[PubMed]
[16] Corselli M., Chen C.W., Crisan M., Lazzari L., Peault B.: Perivascular ancestors of adult multipotent stem cells. Arterioscler. Thromb. Vasc. Biol., 2010; 30: 1104-1109
[PubMed] [Full Text HTML] [Full Text PDF]
[17] Corselli M., Chen C.W., Sun B., Yap S., Rubin J.P., Peault B.: The tunica adventitia of human arteries and veins as a source of mesenchymal stem cells. Stem Cells Dev., 2011; 21: 1299-1308
[PubMed] [Full Text HTML] [Full Text PDF]
[18] Covas D.T., Piccinato C.E., Orellana M.D., Siufi J.L., Silva W.A.Jr., Proto-Siqueira R., Rizzatti E.G., Neder L., Silva A.R., Rocha V., Zago M.A.: Mesenchymal stem cells can be obtained from the human saphena vein. Exp. Cell. Res., 2005; 309: 340-344
[PubMed]
[19] Crisan M., Chen C.W., Corselli M., Andriolo G., Lazzari L., Peault B.: Perivascular multipotent progenitor cells in human organs. Ann. NY Acad. Sci., 2009; 1176: 118-123
[PubMed]
[20] Crisan M., Deasy B., Gavina M., Zheng B., Huard J., Lazzari L., Peault B.: Purification and long-term culture of multipotent progenitor cells affiliated with the walls of human blood vessels: myoendothelial cells and pericytes. Methods Cell Biol., 2008; 86: 295-309
[PubMed]
[21] Crisan M., Huard J., Zheng B., Sun B., Yap S., Logar A., Giacobino J.P., Casteilla L., Peault B.: Purification and culture of human blood vessel-associated progenitor cells. Curr. Prot. Stem Cell Biol., 2008; 2
[PubMed]
[22] Crisan M., Yap S., Casteilla L., Chen C.W., Corselli M., Park T.S., Andriolo G., Sun B., Zheng B., Zhang L., Norotte C., Teng P.N., Traas J., Schugar R., Deasy B.M., Badylak S., Buhring H.J., Giacobino J.P., Lazzari L., Huard J., Peault B.: A perivascular origin for mesenchymal stem cells in multiple human organs. Cell Stem Cell, 2008; 3: 301-313
[PubMed] [Full Text HTML] [Full Text PDF]
[23] Docheva D., Popov C., Mutschler W., Schieker M.: Human mesenchymal stem cells in contact with their environment: surface characteristics and the integrin system. J. Cell. Mol. Med., 2007; 11: 21-38
[PubMed]
[24] Ergun S., Tilki D., Klein D.: Vascular wall as a reservoir for different types of stem and progenitor cells. Antioxid. Redox. Signal., 2011; 15: 981-995
[PubMed]
[25] Fang B., Li Y., Song Y., Li N.: Isolation and characterization of multipotent progenitor cells from the human fetal aorta wall. Exp. Biol. Med., 2010; 235: 130-138
[PubMed]
[26] Fang S., Wei J., Pentinmikko N., Leinonen H., Salven P.: Generation of functional blood vessels from a single c-kit+ adult vascular endothelial stem cell. PLoS Biol., 2012; 10: e1001407
[PubMed] [Full Text HTML] [Full Text PDF]
[27] Folkman J.: Tumor angiogenesis: therapeutic implications. N. Engl. J. Med., 1971; 285: 1182-1186
[PubMed]
[28] Gieryng A., Bogunia-Kubik K.: Znaczenie interakcji między SDF-1 i CXCR4 w hematopoezie i mobilizacji macierzystych komórek hematopoetycznych do krwi obwodowej. Postępy Hig. Med. Dośw., 2007; 61: 369-383
[PubMed] [Full Text HTML] [Full Text PDF]
[29] Gomez-Gaviro M.V., Lovell-Badge R., Fernandez-Aviles F., Lara-Pezzi E.: The vascular stem cell niche. J. Cardiovasc. Transl. Res., 2012; 5: 618-630
[PubMed]
[30] Hirschi K.K., Majesky M.W.: Smooth muscle stem cells. Anat. Rec. A Discov. Mol. Cell Evol. Biol., 2004; 276: 22-33
[PubMed] [Full Text HTML] [Full Text PDF]
[31] Hoglund V.J., Dong X.R., Majesky M.W.: Neointima formation: a local affair. Arterioscle. Thromb. Vasc. Biol., 2010; 30: 1877-1879
[PubMed] [Full Text HTML] [Full Text PDF]
[32] Hoglund V.J., Majesky M.W.: Patterning the artery wall by lateral induction of Notch signaling. Circulation, 2012; 125: 212-215
[PubMed] [Full Text HTML] [Full Text PDF]
[33] Hu Y., Zhang Z., Torsney E., Afzal A.R., Davison F., Metzler B., Xu Q.: Abundant progenitor cells in the adventitia contribute to atherosclerosis of vein grafts in ApoE-deficient mice. J. Clin. Invest., 2004; 113: 1258-1265
[PubMed] [Full Text HTML] [Full Text PDF]
[34] Ingram D.A., Mead L.E., Moore D.B., Woodard W., Fenoglio A., Yoder M.C.: Vessel wall-derived endothelial cells rapidly proliferate because they contain a complete hierarchy of endothelial progenitor cells. Blood, 2005; 105: 2783-2786
[PubMed] [Full Text HTML] [Full Text PDF]
[35] Jaffredo T., Bollerot K., Sugiyama D., Gautier R., Drevon C.: Tracing the hemangioblast during embryogenesis: developmental relationships between endothelial and hematopoietic cells. Int. J. Dev. Biol., 2005; 49: 269-277
[PubMed] [Full Text PDF]
[36] Jaffredo T., Nottingham W., Liddiard K., Bollerot K., Pouget C., de Bruijn M.: From hemangioblast to hematopoietic stem cell: an endothelial connection? Exp. Hematol., 2005; 33: 1029-1040
[PubMed]
[37] Khakoo A.Y., Finkel T.: Endothelial progenitor cells. Annu. Rev. Med., 2005; 56: 79-101
[PubMed]
[38] Kissa K., Herbomel P.: Blood stem cells emerge from aortic endothelium by a novel type of cell transition. Nature, 2010; 464: 112-115
[PubMed] [Full Text HTML] [Full Text PDF]
[39] Klein D., Hohn H.P., Kleff V., Tilki D., Ergun S.: Vascular wall-resident stem cells. Histol. Histopathol., 2010; 25: 681-689
[PubMed]
[40] Klein D., Weisshardt P., Kleff V., Jastrow H., Jakob H.G., Ergun S.: Vascular wall-resident CD44+ multipotent stem cells give rise to pericytes and smooth muscle cells and contribute to new vessel maturation. PLoS One, 2011; 6: e20540
[PubMed] [Full Text HTML] [Full Text PDF]
[41] Kovacic J.C., Boehm M.: Resident vascular progenitor cells: an emerging role for non-terminally differentiated vessel-resident cells in vascular biology. Stem Cell Res., 2009; 2: 2-15
[PubMed] [Full Text HTML] [Full Text PDF]
[42] Krause D.S., Fackler M.J., Civin C.I., May W.S.: CD34: structure, biology, and clinical utility. Blood, 1996; 87: 1-13
[PubMed] [Full Text HTML] [Full Text PDF]
[43] Kume T.: Ligand-dependent Notch signaling in vascular formation. Adv. Exp. Med. Biol., 2012; 727: 210-222
[PubMed]
[44] Lander A.D., Kimble J., Clevers H., Fuchs E., Montarras D., Buckingham M., Calof A.L., Trumpp A., Oskarsson T.: What does the concept of the stem cell niche really mean today? BMC Biol., 2012; 10: 19
[PubMed] [Full Text HTML] [Full Text PDF]
[45] Leri A., Hosoda T., Kajstura J., Anversa P., Rota M.: Identification of a coronary stem cell in the human heart. J. Mol. Med., 2011; 89: 947-959
[PubMed] [Full Text HTML] [Full Text PDF]
[46] Lukasiewicz A., Drewa T., Molski S.: Advances in engineering of blood vessels. Pol. Merkur. Lek., 2007; 23: 439-442
[PubMed] [Full Text PDF]
[47] Majesky M.W.: Developmental basis of vascular smooth muscle diversity. Arterioscler. Thromb. Vasc. Biol., 2007; 27: 1248-1258
[PubMed] [Full Text HTML] [Full Text PDF]
[48] Majesky M.W.: Developmental biology in the vasculature–review series. Arterioscler. Thromb. Vasc. Biol., 2009; 29: 622
[PubMed] [Full Text HTML] [Full Text PDF]
[49] Majesky M.W., Dong X.R., Hoglund V., Daum G., Mahoney W.M.Jr.: The adventitia: a progenitor cell niche for the vessel wall. Cells Tissues Organs, 2012; 195: 73-81
[PubMed] [Full Text HTML] [Full Text PDF]
[50] Majesky M.W., Dong X.R., Hoglund V., Mahoney W.M.Jr., Daum G.: The adventitia: a dynamic interface containing resident progenitor cells. Arterioscler. Thromb. Vasc. Biol., 2011; 31: 1530-1539
[PubMed] [Full Text HTML] [Full Text PDF]
[51] Majesky M.W., Dong X.R., Regan J.N., Hoglund V.J.: Vascular smooth muscle progenitor cells: building and repairing blood vessels. Circ. Res., 2011; 108: 365-377
[PubMed] [Full Text HTML] [Full Text PDF]
[52] Montani D., Perros F., Gambaryan N., Girerd B., Dorfmuller P., Price L.C., Huertas A., Hammad H., Lambrecht B., Simonneau G., Launay J.M., Cohen-Kaminsky S., Humbert M.: C-kit-positive cells accumulate in remodeled vessels of idiopathic pulmonary arterial hypertension. Am. J. Respir. Crit. Care Med., 2011; 184: 116-123
[PubMed]
[53] Nykanen A.I., Krebs R., Saaristo A., Turunen P., Alitalo K., Yla-Herttuala S., Koskinen P.K., Lemstrom K.B.: Angiopoietin-1 protects against the development of cardiac allograft arteriosclerosis. Circulation, 2003; 107: 1308-1314
[PubMed] [Full Text HTML] [Full Text PDF]
[54] Pacilli A., Pasquinelli G.: Vascular wall resident progenitor cells: a review. Exp. Cell Res., 2009; 315: 901-914
[PubMed]
[55] Paprocka M., Krawczenko A., Dus D., Kantor A., Carreau A., Grillon C., Kieda C.: CD133 positive progenitor endothelial cell lines from human cord blood. Cytometry A, 2011; 79: 594-602
[PubMed] [Full Text HTML] [Full Text PDF]
[56] Parker R.C.: The development of organized vessels in cultures of blood cells. Science, 1933; 77: 544-546
[PubMed]
[57] Passman J.N., Dong X.R., Wu S.P., Maguire C.T., Hogan K.A., Bautch V.L., Majesky M.W.: A sonic hedgehog signaling domain in the arterial adventitia supports resident Sca1+ smooth muscle progenitor cells. Proc. Natl. Acad. Sci. USA, 2008; 105: 9349-9354
[PubMed] [Full Text HTML] [Full Text PDF]
[58] Peichev M., Naiyer A.J., Pereira D., Zhu Z., Lane W.J., Williams M., Oz M.C., Hicklin D.J., Witte L., Moore M.A., Rafii S.: Expression of VEGFR-2 and AC133 by circulating human CD34(+) cells identifies a population of functional endothelial precursors. Blood, 2000; 95: 952-958
[PubMed] [Full Text HTML] [Full Text PDF]
[59] Psaltis P.J., Harbuzariu A., Delacroix S., Holroyd E.W., Simari R.D.: Resident vascular progenitor cells – diverse origins, phenotype, and function. J. Cardiovasc. Transl. Res., 2011; 4: 161-176
[PubMed] [Full Text HTML] [Full Text PDF]
[60] Ranjan A.K., Kumar U., Hardikar A.A., Poddar P., Nair P.D., Hardikar A.A.: Human blood vessel-derived endothelial progenitors for endothelialization of small diameter vascular prosthesis. PLoS One, 2009; 4: e7718
[PubMed] [Full Text HTML] [Full Text PDF]
[61] Ritman E.L., Lerman A.: The dynamic vasa vasorum. Cardiovas.Res., 2007; 75: 649-658
[PubMed] [Full Text HTML] [Full Text PDF]
[62] Sainz J., Al Haj Zen A., Caligiuri G., Demerens C., Urbain D., Lemitre M., Lafont A.: Isolation of “side population” progenitor cells from healthy arteries of adult mice. Artherioscler. Thromb. Vasc. Biol., 2006; 26: 281-286
[PubMed] [Full Text HTML] [Full Text PDF]
[63] Schwartz S.M., Benditt E.P.: Cell replication in the aortic endothelium: a new method for study of the problem. Lab. Invest., 1973; 28: 699-707
[PubMed]
[64] Schwartz S.M., Benditt E.P.: Clustering of replicating cells in aortic endothelium. Proc. Natl. Acad. Sci. USA, 1976; 73: 651-653
[PubMed] [Full Text HTML] [Full Text PDF]
[65] Schwartz S.M., Stemerman M.B., Benditt E.P.: The aortic intima. II. Repair of the aortic lining after mechanical denudation. Am. J. Pathol., 1975; 81: 15-42
[PubMed] [Full Text HTML] [Full Text PDF]
[66] Sen S., McDonald S.P., Coates P.T., Bonder C.S.: Endothelial progenitor cells: novel biomarker and promising cell therapy for cardiovascular disease. Clin. Sci., 2011; 120: 263-283
[PubMed] [Full Text HTML] [Full Text PDF]
[67] Shi Q., Rafii S., Wu M.H., Wijelath E.S., Yu C., Ishida A., Fujita Y., Kothari S., Mohle R., Sauvage L.R., Moore M.A., Storb R.F., Hammond W.P.: Evidence for circulating bone marrow-derived endothelial cells. Blood, 1998; 92: 362-367
[PubMed] [Full Text HTML] [Full Text PDF]
[68] Simionescu M., Simionescu N., Palade G.E.: Morphometric data on the endothelium of blood capillaries. J. Cell. Biol., 1974; 60: 128-152
[PubMed] [Full Text HTML] [Full Text PDF]
[69] Simionescu M., Simionescu N., Palade G.E.: Segmental differentiations of cell junctions in the vascular endothelium. The microvasculature. J. Cell. Biol., 1975; 67: 863-865
[PubMed] [Full Text HTML] [Full Text PDF]
[70] Simionescu M., Simionescu N., Palade G.E.: Segmental differentiations of cell junctions in the vascular endothelium. Arteries and veins. J. Cell. Biol., 1976; 68: 705-723
[PubMed] [Full Text HTML] [Full Text PDF]
[71] Sinka L., Biasch K., Khazaal I., Peault B., Tavian M.: Angiotensin-converting enzyme (CD143) specifies emerging lympho-hematopoietic progenitors in the human embryo. Blood, 2012; 119: 3712-3723
[PubMed] [Full Text HTML] [Full Text PDF]
[72] Skora J., Biegus J., Pupka A., Barc P., Sikora J., Szyber P.: Molekularne podstawy angiogenezy. Postępy Hig. Med. Dośw., 2006; 60: 410-415
[PubMed] [Full Text HTML] [Full Text PDF]
[73] Staniszewska M., Sluczanowska-Glabowska S., Drukala J.: Stem cells and skin regeneration. Folia Histochem. Cytobiol., 2011; 49: 375-380
[PubMed]
[74] Tavian M., Biasch K., Sinka L., Vallet J., Peault B.: Embryonic origin of human hematopoiesis. Int. J. Dev. Biol., 2010; 54: 1061-1065
[PubMed] [Full Text PDF]
[75] Tavian M., Cortes F., Robin C., Schiavon V., Hallais M.F., Coulombel L., Charbord P., Labastie M.C., Peault B.: The hemangioblast, common precursor of endothelial and hematopoietic cells. Transfus. Clin. Biol., 2000; 7: 238-241
[PubMed]
[76] Tavian M., Coulombel L., Luton D., Clemente H.S., Dieterlen-Lievre F., Peault B.: Aorta-associated CD34+ hematopoietic cells in the early human embryo. Blood, 1996; 87: 67-72
[PubMed] [Full Text HTML] [Full Text PDF]
[77] Tavian M., Peault B.: Embryonic development of the human hematopoietic system. Int. J. Dev. Biol., 2005; 49: 243-250
[PubMed] [Full Text PDF]
[78] Tavian M., Peault B.: Analysis of hematopoietic development during human embryonic ontogenesis. Methods. Mol. Med., 2005; 105: 413-424
[PubMed]
[79] Tavian M., Zheng B., Oberlin E., Crisan M., Sun B., Huard J., Peault B.: The vascular wall as a source of stem cells. Ann. NY Acad. Sci., 2005; 1044: 41-50
[PubMed]
[80] Tigges U., Komatsu M., Stallcup W.B.: Adventitial pericyte progenitor/mesenchymal stem cells participate in the restenotic response to arterial injury. J. Vasc. Res., 2012; 50: 134-144
[PubMed]
[81] Tilki D., Hohn H.P., Ergun B., Rafii S., Ergun S.: Emerging biology of vascular wall progenitor cells in health and disease. Trends. Mol. Med., 2009; 15: 501-509
[PubMed]
[82] Timmermans F., Plum J., Yoder M.C., Ingram D.A., Vandekerckhove B., Case J.: Endothelial progenitor cells: identity defined? J. Cell. Mol. Med., 2009; 13: 87-102
[PubMed]
[83] Torsney E., Xu Q.: Resident vascular progenitor cells. J. Mol. Cell. Cardiol., 2011; 50: 304-311
[PubMed]
[84] Wnuczko K., Szczepanski M.: Endothelium–characteristics and functions. Pol. Merkur. Lek., 2007; 23: 60-65
[PubMed]
[85] Xiong J.W.: Molecular and developmental biology of the hemangioblast. Dev. Dyn., 2008; 237: 1218-1231
[PubMed] [Full Text HTML] [Full Text PDF]
[86] Yeager M.E., Frid M.G., Stenmark K.R.: Progenitor cells in pulmonary vascular remodeling. Pulm. Circ., 2011; 1: 3-16
[PubMed] [Full Text HTML]
[87] Zampetaki A., Kirton J.P., Xu Q.: Vascular repair by endothelial progenitor cells. Cardiovasc. Res., 2008; 78: 413-421
[PubMed] [Full Text HTML] [Full Text PDF]
[88] Zengin E., Chalajour F., Gehling U.M., Ito W.D., Treede H., Lauke H., Weil J., Reichenspurner H., Kilic N., Ergun S.: Vascular wall resident progenitor cells: a source for postnatal vasculogenesis. Development, 2006; 133: 1543-1551
[PubMed] [Full Text HTML] [Full Text PDF]
Autorzy deklarują brak potencjalnych konfliktów interesu.