
The biological activity of quaternary ammonium salts (QASs)
Ewa Obłąk 1 , Andrzej Gamian 2Streszczenie
Czwartorzędowe sole amoniowe (CSA), zwłaszcza te o charakterze kationowych surfaktantów są stosowane jako antybakteryjne i antygrzybowe dezynfektanty. CSA działają również na wirusy z otoczką lipidową, włączając HIV (human immunodeficiency virus) i HBV (hepatitis B virus), ale nie działają na wirusy bez otoczki lipidowej. Związki te mają szerokie zastosowanie w gospodarstwie domowym (szampony, odżywki do włosów), w rolnictwie (fungicydy, pestycydy, insektycydy), w ochronie zdrowia (leki) oraz w przemyśle (biocydy, środki antykorozyjne).
Stosowanie czwartorzędowych soli amoniowych na szeroką skalę, jako dezynfektantów doprowadziło do rozwoju oporności mikroorganizmów na te związki. Staphylococcus aureus zawiera plazmid niosący geny (qacA i qacB) kodujące oporność na czwartorzędowe związki amoniowe i akryflawinę. Białka błonowe QacA i QacB (tzw. pompy efflux) nadają wieloraką oporność przez usuwanie związków z komórki.
Słowa kluczowe:czwartorzędowe sole amoniowe • oporność • surfaktanty • Saccharomyces cerevisiae
Summary
Quaternary ammonium salts (QASs), especially those of cationic surfactant character, are applied as antibacterial and antifungal disinfectants. QASs affect lipid-enveloped viruses, including human immunodeficiency virus (HIV) and hepatitis B virus (HBV), but not non-enveloped viruses. These compounds are extensively used in domestic (as ingredients of shampoos, hair conditioners), agricultural (as fungicides, pesticides, insecticides), healthcare (as medications), and industrial applications (as biocides, fabric softeners, corrosion inhibitors). The extensive use of quaternary ammonium disinfectants in recent years has led to the development of resistance in microorganisms to these drugs. Thus Staphylococcus aureus strains contain the plasmid-carrying genes qacA and qacB encoding resistance to quaternary ammonium compounds and acriflavine. The membrane proteins QacA and QacB confer multidrug resistance by exporting the compound by the proton motive force which is generated by the transmembrane electrochemical proton gradient.
Key words:quaternary ammonium salt • resistant • surfactant • Saccharomyces cerevisiae
1. Czwartorzędowe sole amoniowe (CSAsa)
Czwartorzędowe sole amoniowe (CSA) wykazują działanie bakteriobójcze i grzybobójcze, są stosowane, na szeroką skalę, jako środki dezynfekcyjne. Długotrwałe ich stosowanie spowodowało powstanie oporności na te związki wśród szczepów bakteryjnych, jak również grzybów. Bezpośrednią przyczyną oporności mikroorganizmów na różnego rodzaju związki jest obecność i ekspresja odpowiednich genów w komórce o lokalizacji chromosomalnej bądź plazmidowej [23,50].
Aby zwalczyć problem oporności wśród mikroorganizmów prowadzone są intensywne badania nad syntezą nowych skutecznie działających dezynfektantów. Racjonalne szukanie środków biologicznie czynnych polega na takim projektowaniu struktury związku, aby oddziaływał on z wybranym receptorem biologicznym. Struktury komórkowe najbardziej eksponowane na działanie związków chemicznych to błony komórkowe, a zwłaszcza plazmalemma.
Szczególnie silnie oddziaływają z błoną substancje o charakterze amfifilowym. Do tej grupy związków można zaliczyć czwartorzędowe sole amoniowe. CSA są to pochodne jonu amoniowego. Biologicznie czynne CSA stanowią grupę związków amfifilowych o ogólnym wzorze (ryc. 1), gdzie podstawnik R4 stanowi długi łańcuch alkilowy o różnej liczbie atomów węgla, a R1, R2, R3 to m.in. grupy alkilowe: nasycone, nienasycone, rozgałęzione lub nierozgałęzione, cykliczne lub acykliczne [64]. Dodatnio naładowany atom azotu wraz z grupami alkilowymi stanowi hydrofilową część związku, natomiast łańcuch alkilowy o różnej liczbie atomów węgla jest jego częścią hydrofobową.

Ryc. 1. Struktura chemiczna czwartorzędowych soli amoniowych (CSA); R4 – długi hydrofobowy łańcuch alkilowy o różnej liczbie atomów węgla, R1 R2 R3 – grupy alkilowe, X– – anion chlorkowy lub bromkowy
Amfifilowość umożliwia tym związkom wchodzenie w reakcje z lipidową częścią błony komórkowej i przez zakłócanie jej funkcji, CSA mogą pośrednio wpływać na aktywność enzymów, które biorą udział w transporcie przez dwuwarstwę lipidową [5,10]. Czwartorzędowe sole amoniowe, jako kationowe surfaktanty, są szeroko wykorzystywane w wielu dziedzinach przemysłu (chemicznym, kosmetycznym, drzewnym – jako środki do płukania, szampony, odżywki do włosów, biocydy), rolnictwie, leśnictwie (pestycydy, fungicydy, insektycydy), medycynie (środki dezynfekcyjne, leki) [32,44,73,89].
Czwartorzędowe sole amoniowe ze względu na budowę strukturalną, możemy podzielić na kilka klas [64]: pochodne alifatyczne (np. glicynobetaina, charanina, antopleuryna); pochodne pirydyny (np. homaryna, trigonellina); pochodne pyrrolidynowe (np. faskoplazyna, stachydryna); pochodne indolu (np. bromgramina); pochodne imidazolu (np. zooanemonina); pochodne puryny (np. herbipolina); pochodne benzenu (np. kandycyna).
1.1. Czwartorzędowe sole amoniowe naturalnie występujące w przyrodzie
Występujące w przyrodzie czwartorzędowe sole amoniowe (CSA) tworzą klasę ponad 100 poznanych metabolitów [64]. Charakteryzują się one szerokim zakresem występowania: od bakterii, poprzez glony, grzyby, rośliny, bezkręgowce aż do kręgowców, a także różnorodnością ról, jakie spełniają. Przypuszcza się, że organizmy syntetyzują te substancje w celu lepszego przystosowania się do warunków środowiska np. zasolenia, nagłych zmian temperatury. I tak np. u bakterii Escherichia coli, Salmonella Typhimurium i Klebsiella pneumoniae wykryto obecność glicynobetainy, która pozwala im przetrwać w środowisku, kiedy wzrasta jego zasolenie [9,87,90]. Natomiast u symbionta brodawek korzeniowych, Rhizobium meliloti, glicynobetaina ma dwie funkcje: przy małej osmolarności środowiska zużywana jest jako pożywienie, natomiast gdy wzrasta stres osmotyczny, pełni funkcje ochronne [48,82]. Obecność glicynobetainy stwierdzono u większości bezkręgowców, gdzie oprócz tolerancji osmotycznej, chroni przed niską temperaturą. Czwartorzędowe sole ammoniowe biorą także udział w transporcie i metabolizmie lipidów – i tak karnityna (ryc. 2) ma zdolność przenoszenia kwasów tłuszczowych przez wewnętrzną błonę mitochondrialną oraz wpływa na cytoplazmatyczną koncentrację acetylo- CoA [3,77].

Ryc. 2. Struktura chemiczna karnityny
Niektóre z CSA są źródłem węgla, azotu np. butyrobetaina u Agrobacterium sp. [52]. Ponadto stanowią one rezerwuar grup metylowych, takich jak homaryna u skorupiaków [51]. Homaryna również bierze udział w adaptacji osmotycznej morskich glonów, hamuje metamorfozę niektórych bezkręgowców ze stanu larwalnego do postaci dorosłej [13]. Czwartorzędowe sole amoniowe (np. trigonellina) redukują stężenie cholesterolu we krwi szczurów [1] i hamują rozwój leukemii [2]. Jeszcze inne CSA (bromogramina) mogą hamować mitotyczne podziały komórkowe [76] lub spełniać funkcje antybiotyku (charamina) [4]. Niektóre z nich są inhibitorami Na+ i K+-ATP-azy [3]. Wreszcie są wśród nich też substancje (faskaplazyna) wykazujące właściwości bakteriobójcze i cytotoksyczne [70]. Powyższe przykłady są dowodem, że czwartorzędowe sole amoniowe o różnej swoistej budowie strukturalnej pełnią bardzo różnorodne funkcje fizjologiczne w wielu procesach żywych organizmów.
1.2. Własności syntetycznych czwartorzędowych soli amoniowych
Duża różnorodność budowy czwartorzędowych soli amoniowych (CSA) i pełnionych przez nie funkcji w żywych organizmach, przyczyniła się do intensywnych badań nad syntetyzowaniem nowych tego typu związków o specyficznej, ściśle określonej budowie chemicznej, w nadziei ich wykorzystania w różnych dziedzinach. Już Jacobs, Heidelberger i Amoss [64] uzyskali wiele różnych CSA o właściwościach bakteriobójczych. Jednak silne bakteriobójcze działanie czwartorzędowych soli amoniowych o długim hydrofobowym łańcuchu alkilowym ujawnił Domagk [14].
Amfifilowy charakter CSA przyczynił się do badań nad działaniem tych związków na modelowe błony lipidowe [6,38,39] i błony biologiczne [36]. Badania te wykonywane na sztucznych błonach lipidowych wykazały, że aktywność CSA zależy od długości hydrofobowego łańcucha alkilowego, który penetruje wnętrze hydrofobowe błony oraz od rozmiarów części polarnej cząsteczki [38]. Wykazano, że czwartorzędowe sole amoniowe o budowie amfifilowej zwiększają płynność hydrofobowego wnętrza błony [84]. Wyniki tych badań zostały potwierdzone na błonach biologicznych. Wykazano, że długość łańcucha alkilowego ma ścisły związek z poziomem aktywności czwartorzędowych soli amoniowych. Te CSA, które mają 10–12 atomów węgla w łańcuchu alkilowym, wykazują najwyższą aktywność. Aktywność ich spada wraz ze wzrostem długości łańcucha alkilowego powyżej 12 atomów węgla (14–16), a także poniżej 10 atomów węgla [57]. Bakteriobójczy wpływ tych związków zależy również od ich liofilowości – sole o obniżonej skłonności do rozpuszczania się w tłuszczach charakteryzują się obniżoną aktywnością, głównie wobec bakterii Gram-dodatnich [80].
W niskich stężeniach CSA występują w postaci monomerów natomiast, gdy ich stężenie wzrasta tworzą micele [38]. Zarówno monomery, jak i formy micelarne mają zdolność oddziaływania z błoną [71,72], jednak w lipidową dwuwarstwę wbudowują się wyłącznie monomery [45]. Duży wpływ na ich aktywność może mieć też pH, temperatura oraz stężenie innych jonów [46,53].
Stwierdzono również, że wraz ze wzrostem stężenia związków występowało zwiększenie efektu ich działania nie tylko na błony modelowe, ale również na obiekty biologiczne. Małe stężenia CSA działają bardziej swoiście na określone domeny białka. Helenius i Simons [28] sugerują, że przyłączenie się substancji o budowie amfifilowej do swoistych miejsc wywołuje w białku zmiany konformacyjne i może odsłonić nowe miejsca o powinowactwie do tych związków. Natomiast duże stężenia tych substancji mogą powodować denaturację białka [40] lub też cząsteczki białka mogą być oddzielane i włączone do miceli detergentów, co zaburza pierwotne ułożenie białek w błonie. CSA wykazują więc dużą reaktywność względem naturalnych biologicznych struktur błonowych; mogą wpływać na płynność dwuwarstwy fosfolipidowej, zaburzać swoiste oddziaływania hydrofobowe i elektrostatyczne między białkami błonowymi i lipidami oraz mogą zmieniać symetrię ułożenia lipidów [8]. Działając na enzymy błonowe związki te mogą powodować ich inhibicję przez wypłukiwanie lipidu, którego obecność jest niezbędna do prawidłowego funkcjonowania enzymu lub go inaktywować rozrywając słabe wiązania między podjednostkami enzymów oligomerycznych, takimi jak ATP-aza mitochondrialna czy ATP-aza wakuolarna. Udowodniono, że całkowita delipidacja H+-ATP-azy błony plazmatycznej u Saccharomyces cerevisiae, doprowadza do inaktywacji tego enzymu [17,78]. Do prawidłowego funkcjonowania ATP-aza typu P wymaga obecności lipidów zawierających ładunek ujemny w części hydrofilowej cząsteczki oraz nieuszkodzonych łańcuchów kwasów tłuszczowych w jej części hydrofobowej (warunki te spełniają fosfatydyloinozytol (PI) i fosfatydyloglicerol (PG)), a także obecności ergosterolu [86]. Obecność w błonie biologicznej związku amfifilowego o dodatnio naładowanej części hydrofilowej cząsteczki, wydaje się wpływać inhibicyjnie na aktywność białek błony komórkowej: H+-ATP-azę błony komórkowej i ogólną permeazę aminokwasową GAP [41,42,55,88]. Tak więc małe stężenia czwartorzędowych soli amoniowych mogą powodować zmianę właściwości białek, natomiast duże stężenia CSA prowadzą do rozpuszczenia białek i lipidów, co powoduje destrukcję błon.
Wyniki badań nad działaniem czwartorzędowych soli amoniowych na błony biologiczne, oprócz wartości poznawczych, mogą mieć również znaczenie aplikacyjne. Wskazują na możliwość wykorzystania biologicznie czynnych czwartorzędowych soli amoniowych do zwalczania szkodliwych drobnoustrojów. Poznanie natomiast mechanizmu ich działania może prowadzić do racjonalnej syntezy nowych związków tego typu z możliwością wykorzystania ich w medycynie, rolnictwie, leśnictwie i wielu innych dziedzinach gospodarki.
1.3. Zastosowanie czwartorzędowych soli amoniowych
Kationowe detergenty, jakimi są czwartorzędowe sole amoniowe (CSA), odznaczają się dużą aktywnością powierzchniową. Związki te mają zdolność łatwej dysocjacji w wodzie, w wyniku której powstają dodatnio naładowane jony kompleksowe i ujemnie naładowane jony chlorowca (do dezynfekcji bowiem dobiera się środki zawierające chlorowiec – chlor lub brom, ponieważ wzmaga on czynność bakteriobójczą). Grupa kationowa nadaje tym związkom dużą aktywność powierzchniową (zdolność obniżania napięcia powierzchniowego). Mechanizm takiego oddziaływania ułatwia adsorpcję kationu na powierzchni komórki i wnikanie do błony drobnoustrojów reszt hydrofobowych o działaniu bakteriobójczym. Stąd związki te mają szerokie zastosowanie, jako środki bakteriobójcze i grzybobójcze.
Celem działania czwartorzędowych soli amoniowych są błony komórkowe. W komórkach organizmów, które miały kontakt z kationowymi surfaktantami zachodzi następująca sekwencja zdarzeń:
• adsorpcja i penetracja czynnika przez ścianę komórkową,
• reakcja ze składnikami błony cytoplazmatycznej (białka lub lipidy), której następstwem jest dezintegracja błony,
• wyciek małocząsteczkowych wewnątrzkomórkowych składników,
• degradacja białek i kwasów nukleinowych,
• liza ściany komórkowej spowodowana działaniem autolitycznych enzymów [47,75].
W ten sposób następuje utrata organizacji strukturalnej i integralności błony cytoplazmatycznej u bakterii w połączeniu z innymi szkodliwymi efektami dla komórki bakteryjnej [12]. Prowadzone badania potwierdziły, że CSA wykazują dobrą skuteczność w stosunku do bakterii Gram-dodatnich, jak i Gram-ujemnych, chociaż te drugie mają tendencję do wykazywania oporności na działanie tych związków [12,47].
Takim znanym środkiem do zwalczania drobnoustrojów jest chlorek benzalkoniowy. Chlorek ten działa bakteriobójczo na bakterie Gram-dodatnie, a w dużych stężeniach hamuje rozwój bakterii Gram-ujemnych, grzybów, wirusów, bakterii kwasoodpornych. Działanie tego związku polega na łączeniu się z lipidami błon komórkowych drobnoustrojów powodując wytwarzanie na ich powierzchni nieprzepuszczalnej warstwy, która utrudnia wymianę substancji między wnętrzem komórki a otoczeniem, wynikiem czego jest zaburzenie procesów metabolicznych, prowadzące do śmierci komórki bakteryjnej [7,37].
U drożdży Saccharomyces cerevisiae czwartorzędowe sole amoniowe powodują wyciek jonów K+ i pentoz z komórki, indukują lizę protoplastu [29]. Ich toksyczne działanie polega na dezorganizacji błony plazmatycznej, spowodowanej rozerwaniem dwuwarstwy lipidowej [47]. Czwartorzędowe sole amoniowe są sporostatyczne, hamują rozwój komórki wegetatywnej ze spory [74]. CSA wykazują również działanie przeciwko wirusom otoczkowym HIV (human immunodeficiency virus) i HBV (hepatitis B virus) [47,83]. Produkty oparte na czwartorzędowych solach amoniowych powodują zmiany morfologiczne w strukturze ludzkiego wirusa HBV, czego rezultatem jest utrata przez wirusa infekcyjności [65]. Środki dezynfekcyjne na bazie CSA łączą wiele zalet, m.in. przy małej toksyczności, wykazują dużą aktywność antybakteryjną, są dobrze rozpuszczalne w wodzie, działają w szerokim zakresie pH (4–10), nie powodują korozji czyszczonych powierzchni [47]. Najczęściej stosowane są chlorki alkilodimetylobenzyloamoniowe, ponieważ są bezpieczne w użyciu i stosunkowo łatwo ulegają biodegradacji. Są skutecznymi dezynfektantami, gdyż powodują denaturację białka drobnoustrojów. Niszczą bakterie, ich przetrwalniki, grzyby i (nieco słabiej) wirusy.
Czwartorzędowe sole amoniowe ze względu na swoje charakterystyczne właściwości środków powierzchniowo czynnych stosowane są również, jako substancje antykorozyjne oraz jako antyelektrostatyczne wobec takich materiałów jak wełna, bawełna, włókna syntetyczne i celulozowe, wykorzystuje się je także jako zmiękczacze w produkcji tkanin i papieru [67]. W produktach użytku domowego substancje te stosowane są głównie do produkcji odżywek regeneracyjnych, balsamów do włosów i środków piorących [67]. Dzięki działaniu antystatycznemu CSA mogą mieć zastosowanie jako środki do płukania tkanin. Środki te dodane do ostatniej kąpieli płuczącej, powodują nie tylko usunięcie szorstkości i sztywności tkanin, ale nadają im miękkość i powodują odpływ ładunków elektrostatycznych. Ponadto czwartorzędowe sole amoniowe są wykorzystywane w technologii wytwarzania i przetwórstwa nienasyconych żywic poliestrowych, które służą przede wszystkim do wyrobu laminatów poliestrowo-szklanych, typowymi przykładami są kadłuby łodzi, jachtów, przenośne kabiny, umywalki, wanny itp. [33].
Oprócz tego, czwartorzędowe sole amoniowe, jako kationowe surfaktanty znalazły zastosowanie jako biocydy w ochronie drewna [12,16,73]. Związki te stosowane są również w przemyśle do zwalczania mikroorganizmów tworzących biofilmy w układach chłodniczych [73], a także w rolnictwie do ochrony roślin przed chorobami wywoływanymi przez grzyby.
Czwartorzędowe sole amoniowe znalazły również zastosowanie w anestezjologii jako leki zwiotczające mięśnie szkieletowe. Działają jako inhibitory transmisji synaptycznej neuronów poprzez blokowanie nikotynowego receptora cholinergicznego, przy czym związki zawierające dwie grupy amoniowe mają zdolność przyłączania się do receptorów nikotynowych błony postsynaptycznej neuronów [44]. Do najbardziej znanych leków należą D-tubokuraryna, toksyferyna [66].
Obecnie trwają badania nad wykorzystaniem związku, DMAE-CB (methacryloxylethyl cetyl ammonium chloride), jako dodatku do plomb i cementów dentystycznych. Stwierdzono jego skuteczne działanie bakteriobójcze względem wysoko próchnicotwórczych i wirulentnych szczepów bakteryjnych jamy ustnej: Actinomyces viscosus ATCC 15987 oraz Staphylococcus aureus ATCC 29213 [89].
Ponadto, związki te ze względu na właściwości bakteriobójcze, znalazły zastosowanie w medycynie jako środki dezynfekcyjne (dezynfekcje przedoperacyjne nieuszkodzonej skóry czy błony śluzowej, narzędzi chirurgicznych, sal operacyjnych) [47].
Czwartorzędowe sole amoniowe mają również zastosowanie w leczeniu takich chorób jak cukrzyca, arytmia serca, nerwice, alergie [24], a także wspomagają działanie leków przeciwnowotworowych. Syntetyczna czwartorzędowa sól amoniowa, bromek ipratropium (pochodna atropiny), stosowany przez zakażonych Retrowirusem typu 39 (który wywołuje infekcje górnych dróg oddechowych) powoduje zahamowanie symptomów choroby u połowy badanych [20]. Związek ten ma zastosowanie w leczeniu astmy oskrzelowej, poprawiając sprawność wentylacyjną płuc [68]. Czwartorzędowe sole amoniowe znalazły zastosowanie również w innych dziedzinach medycyny. Jodek trimetyloamoniowy stosowany jest w oftalmologii u pacjentów z jaskrą, do zmniejszenia ciśnienia wewnątrzgałkowego [25]. Chlorek benzalkoniowy znalazł zastosowanie jako substancja konserwująca w praktyce farmaceutycznej – przede wszystkim do jałowych postaci leków okulistycznych, głównie kropli do oczu. Ma także zastosowanie w urologii – przepłukiwanie pęcherza moczowego, a także w ginekologii – płukanie pochwy oraz jako dodatek do kąpieli odkażających [11,37].
Udowodniono, że chitozan powstający w wyniku N-deacylowania chityny wykazuje również aktywność antyoksydacyjną poprzez zdolność usuwania z komórki anionu nadtlenkowego, wolnego rodnika powstającego w wyniku metabolizmu tlenowego aerobów. Uważa się, że to właśnie zwiększona liczba wolnych rodników jest jedną z głównych przyczyn chorób neurodegradacyjnych, nowotworów, zaćmy, różnorodnych stanów zapalnych. Antyoksydacyjne zdolności chitozanu związane są z tym, że anion ponadtlenkowy reaguje z aktywnymi atomami wodoru czwartorzędowych soli amoniowych tworząc formy bardziej stabilne (przy czym sam chitozan nie wykazuje aktywności antyoksydacyjnych) [92]. Dodatkowo chitozan ulega szybkiej biodegradacji, wykazuje aktywność bakteriobójczą, ma także właściwości przeciwnowotworowe [63].
Czwartorzędowe sole amoniowe wykorzystane są w nanotechnologii, jako dodatek do nanowłókien stosowanych w inżynierii tkankowej, poprawiając ich biodegradalność i porowatość – dzięki nim można uzyskać włókniny będące odpowiednikiem kolagenu, które umożliwiają wzrost i adhezje komórek, co może mieć zastosowanie, jako antybakteryjne opatrunki stosowane bezpośrednio na żywe tkanki [63].
2. Badania nad działaniem czwartorzędowych soli amoniowych na mikroorganizmy
Powinowactwo czwartorzędowych soli amoniowych (CSA) do błon biologicznych sprawia, że działanie tych substancji nie ogranicza się do bakterii, glonów, ale obejmuje inne mikroorganizmy, w tym grzyby.
Badania nad grzybobójczym działaniem CSA były prowadzone w licznych ośrodkach. Wykazano, że nadtlenek N,N-dimetylo-1-metylo-dodecyloamoniowy hamuje wzrost nie tylko bakterii, ale drożdży i innych grzybów, a także przy pewnych stężeniach powoduje lizę ich protoplastów [85]. Z kolei chlorki N-alkilo-metyloamoniowe działały hemolitycznie na erytrocyty [31].
Dużo informacji na ten temat dostarczyły badania prowadzone w Instytucie Mikrobiologii Uniwersytetu Wrocławskiego pod kierunkiem prof. T.M. Lachowicza. Testowano serię fungicydów – benzylopochodnych czwartorzędowych soli amoniowych syntetyzowanych w Instytucie Technologii Organicznej i Tworzyw Sztucznych Politechniki Wrocławskiej przez prof. S. Witka. Określano aktywność biologiczną testowanych związków, ich wpływ na transport metioniny, na integralność błony komórkowej, procesy oddechowe, przeżywalność komórek Saccharomyces cerevisiae i kilka innych gatunków grzybów [36]. Efektem tych prac było stwierdzenie destrukcyjnego działania badanych związków (w wyższych stężeniach niż progowe) na komórkę grzyba w warunkach in vivo. Okazało się, że celem działania tych związków jest błona komórkowa, a wczesnym obserwowanym efektem jest natychmiastowe zahamowanie aktywnego transportu metioniny do komórki, po którym następuje wyciekanie z komórki niskocząsteczkowych składników cytoplazmy do środowiska. Świadczyło to o wzrastającej dezintegracji błony komórkowej powodującej śmierć komórki. Związki te były uprzednio testowane na liposomowym modelu błon [18,19,31], a ich wyniki były zgodne z rezultatami uzyskanymi na całych komórkach drożdży.
Badania nad chlorkami alkoksymetyloamoniowymi, zawierającymi łańcuchy alkilowe o różnej liczbie atomów węgla, wykazały ich inhibicyjny wpływ na procesy oddechowe, na regenerację protoplastów, na tempo wzrostu komórek i transport aminokwasów (metioniny) u drożdży Saccharomyces cerevisiae. Sugerowano przy tym, że największą aktywność wykazują związki zawierające łańcuch alkilowy zbudowany z 10–14 atomów węgla [81]. Wyniki te potwierdziły wcześniejsze rezultaty badań na modelu liposomowym błon i błonach erytrocytów [18,19,31].
Ponadto stwierdzono, że czwartorzędowe sole amoniowe i ich pochodne (chlorek benzalkoniowy, cetrimide, 3DOBP-4,10) działają bakteriobójczo (zarówno na bakterie Gram-dodatnie jaki i Gram-ujemne) i grzybobójczo na komórki Candida species i Saccharomyces cerevisiae, a fungistatycznie na Aspergillus ochraceus [26,27,79].
Badania wykazały, że chlorek chlorek N-dodecyloksy- karboksymetylo-N,N,N-trimetyloamoniowy inhibuje wzrost komórek drożdży. Ta inhibicja zależy od pH środowiska. Optymalne pH dla wzrostu drożdży jest w granicach 5,0–6,0. Tolerują one także wzrost w pH 8,0. Jednak w tych warunkach CSA działają inhibicyjnie na wzrost drożdży przy zdecydowanie niższym stężeniu, niż gdy pH jest obojętne lub lekko kwaśne [57,58].
Ponadto okazało się, że wrażliwość drożdży na te substancje jest uwarunkowana tłem genetycznym. Na to tło genetyczne zarówno u Saccharomyces cerevisiae, jak i u drożdżopodobnego organizmu Trichosporum cutaneum składa się zdolność do biosyntezy związków niskocząsteczkowych przede wszystkim aminokwasów [43,61]; mutanty auksotroficzne pozbawione tych zdolności biosyntetycznych wykazywały znacznie większą wrażliwość mierzoną minimalnym inhibicyjnym stężeniem (MIC) niż izogeniczne formy wyjściowe (prototrofy). Ponadto podwójne auksotrofy były na ogół jeszcze bardziej wrażliwe niż pojedyncze [43].
Wysunięto hipotezę, że czwartorzędowe sole amoniowe inhibują aktywny transport aminokwasów do komórki drożdży. Stąd, auksotrofy na podłożu pełnym (YPG) w obecności CSA nie mogąc pobierać wymaganych do wzrostu aminokwasów (których syntezę blokuje defekt genetyczny), giną na skutek braku niezbędnych im aminokwasów [43,61]. Stwierdzono również, że czwartorzędowe sole amoniowe inhibują ogólną permeazę aminokwasową GAP w warunkach derepresji [41,55,59]. Również arginina powoduje hamowanie transportu aminokwasów do komórki drożdży [41,42]. Wiadomo, że enzym H+-ATP-aza błony komórkowej odgrywa główną rolę w transporcie aminokwasów do komórki. Jej aktywność jest odpowiedzialna za utrzymanie gradientu elektrochemicznego błony komórkowej, od którego zależy transport substancji odżywczych. Ponadto wykazano, że czwartorzędowe sole amoniowe są inhibitorami zarówno H+-ATP-azy błony komórkowej, jak i ATP-azy mitochondrialnej [55,57,58].
Innym czynnikiem, od którego uzależniona jest wrażliwość na CSA to efektywność systemu oddechowego, czyli syntezy ATP. Drożdże są względnymi tlenowcami. W warunkach tlenowych uzyskują energię tak jak inne komórki eukariotyczne w wyniku fosforylacji oksydacyjnej. W warunkach beztlenowych, a także przy względnym nadmiarze glukozy drożdże uzyskują ATP w procesie fosforylacji substratowej. U drożdży piekarniczych znane są i bardzo dobrze scharakteryzowane różnego typu mutanty z defektami w oddychaniu. Jednym z typów takich mutantów są rho– i rho0 [61,62]. Nie są one zdolne do oddychania, co wynika z braku cytochromów a+a3 i b. Stąd nie mogą one korzystać z niefermentowalnych źródeł energii, takich jak etanol, glicerol, pirogronian czy mleczan. Otóż te mutanty defektywne oddechowo z mutacjami na poziomie mitochondrialnego DNA wykazują znacznie większą wrażliwość na czwartorzędową sól amoniową (chlorek N-dodecyloksykarboksymetylo-N,N,N-trimetyloamoniowy) niż izogeniczny szczep kompetentny oddechowo (rho+) (ryc. 3) [61,62]. Natomiast defekt oddechowy będący wynikiem mutacji w genach jądrowych (mutanty pet) oddziaływał różnorodnie; jedne mutanty monogeniczne wykazywały zwiększoną wrażliwość, taką jak rho– czy rhoo lub nie różniły się zasadniczo pod tym względem od szczepu wyjściowego, kompetentnego oddechowo (rho+). Były też formy superwrażliwe [60,61,62].

Ryc. 3. Wrażliwość mutantów oddechowych Saccharomyces cerevisiae (rho– i rhoo) na czwartorzędową sól amoniową IM (chlorek N-(dodecyloksykarboksymetylo)-N,N,N-trimetyloamoniowy) (w stężeniach końcowych 800, 1600, 3200, 6400 µM): A) szczep kompetentny oddechowo rho+; B) mutant rho–; C) mutant rhoo.
Aby zlokalizować bardziej precyzyjnie cel działania czwartorzędowej soli amoniowej na komórki drożdży zmiany morfologiczne w komórkach były obserwowane w transmisyjnym mikroskopie elektronowym (ryc. 4). Zaobserwowano, że znaczna frakcja komórek poddanych działaniu CSA wykazuje deformację ściany i błony cytoplazmatycznej. Wewnątrz komórkę wypełniały elektronowo gęste ziarnistości. W cytoplazmie można było zobaczyć pojedyncze krople lipidowe. W takich komórkach trudno było zidentyfikować typowe wakuole czy inne organelle komórkowe. Takich zmian nie obserwowano w komórkach inkubowanych bez związku [54]. Wykazano również, że w komórkach drożdży w obecności czwartorzędowej soli amoniowej (chlorku N-dodecyloksykarboksymetylo- N,N,N-trimetyloamoniowego) było zwiększone wytwarzanie lipidów. Zmiany te były obserwowane na ultracienkich skrawkach komórek drożdży w transmisyjnym mikroskopie elektronowym (ryc. 5) [56].
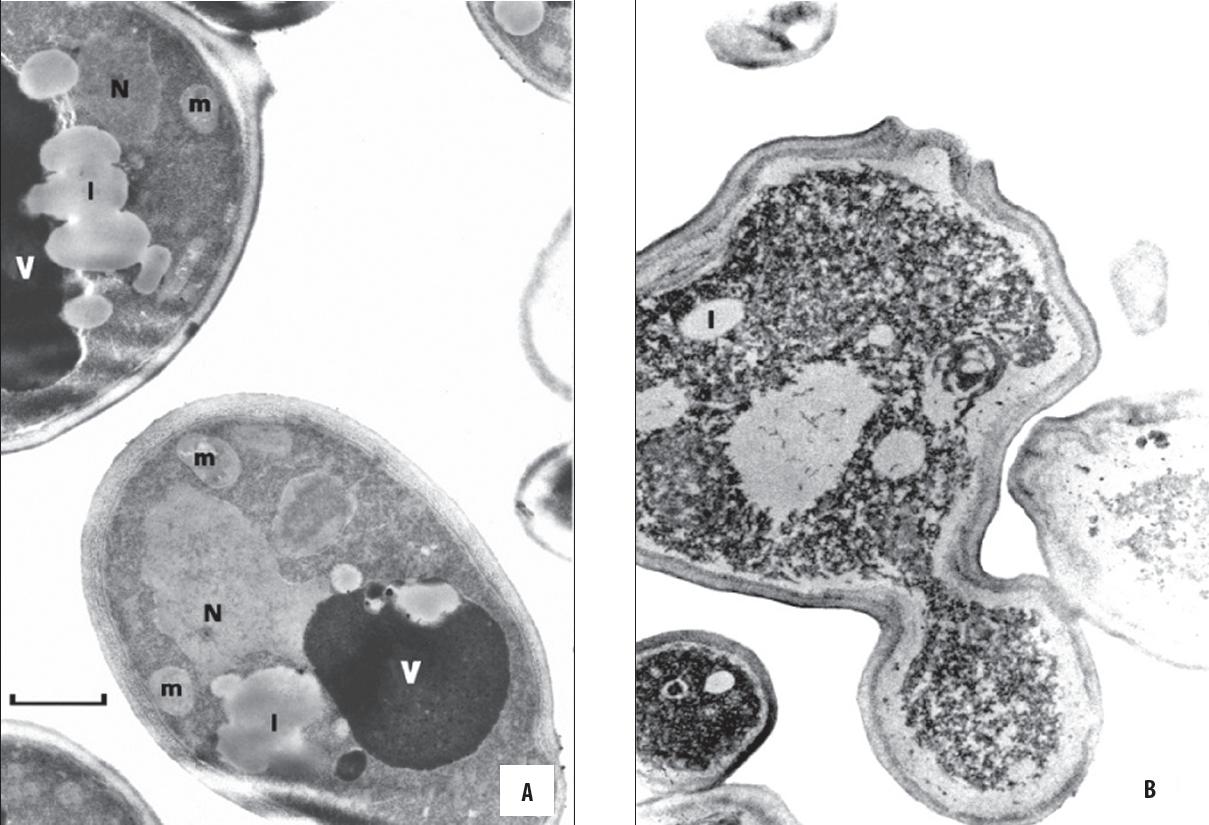
Ryc. 4. Ultracienkie skrawki preparatów komórek drożdży Saccharomyces cerevisiae (szczepu wrażliwego na CSA – metylobromek N,N-dimetyloalaninianu n-dodecylowego (DMALM-12)) z transmisyjnego mikroskopu elektronowego [54]: A) inkubowane w podłożu YPD bez CSA (kontrola); B) inkubowane w podłożu YPD w obecności CSA; v – wakuola; l – krople lipidowe; N – jądro komórkowe; m – mitochondria

Ryc. 5. Ultracienkie skrawki preparatów komórek drożdży Saccharomyces cerevisiae (mutanta opornego na CSA-IM (chlorek N-(dodecyloksykarboksymetylo)-N,N,N-trimetyloamoniowy) z transmisyjnego mikroskopu elektronowego [56]: A) inkubowane w podłożu YPD bez CSA (kontrola); B,C,D) inkubowane w podłożu YPD w obecności CSA; v – wakuola; l – krople lipidowe; N – jądro komórkowe; m – mitochondria
Badania Shirai i wsp. [79] w transmisyjnym mikroskopie elektronowym ultracienkich skrawków komórek drożdży inkubowanych w obecności CSA (gemini) wykazały, że struktura ściany komórkowej była zachowana, a zniszczeniu uległy wewnętrzne organelle komórkowe.
Dubnikowa i wsp. [15] stwierdzili, że kompozycja kwasów tłuszczowych zewnętrznej błony E. coli była modyfikowana w obecności czwartorzędowych soli amoniowych. Różnice dotyczyły ilości i jakości kwasów tłuszczowych. Wiadomo, że czwartorzędowe sole amoniowe w środowisku płynnym dysocjują i oddziaływają z polarnymi komponentami błony, zmieniając jej płynność i przepuszczalność.
Znajomość mechanizmu działania CSA jest więc istotna ze względów teoretycznych, tj. poznania struktury i funkcji błony komórkowej, jak również z praktycznego punktu widzenia, zastosowania ich w medycynie i rolnictwie.
3. Mechanizmy oporności na czwartorzędowe sole amoniowe i inne inhibitory
W populacji grzybów, która jest początkowo wrażliwa na fungicyd, mogą się pojawiać formy oporne. Może być to spowodowane zmianami genetycznymi lub fenotypowymi w komórce grzyba. Jeżeli zmiany te nie są związane z mutacjami w chromosomalnym DNA, to jest to rezultatem adaptacji fenotypowej. Adaptacja może wystąpić w hodowli po długiej ekspozycji na fungicyd w warunkach korzystnych dla organizmu. Oporność fenotypowa jest zwykle tracona po przeniesieniu do środowiska wolnego od fungicydu.
Bezpośrednią przyczyną oporności bakterii na różnego rodzaju związki jest obecność i ekspresja odpowiednich genów w komórce o lokalizacji chromosomalnej bądź plazmidowej [23,50]. Ważnym i rozpowszechnionym mechanizmem oporności drobnoustrojów na różne związki jest ich czynne usuwanie z wnętrza komórki z użyciem transporterów błonowych ABC (ATP – binding cassette) i MFS (major facilitator superfamily) [34,35,69,91]. I tak za zjawisko oporności na czwartorzędowe związki amoniowe, bromek etydyny, akryflawinę u Staphylococcus aureus odpowiedzialne są geny qacA i qacB bakteryjnych pomp efflux QacA i QacB [22,49]. Produkty tych genów, białka QacA i QacB należą do rodziny MFS transporterów. Gen qacA występuje na plazmidzie, ale może także być umiejscowiony na chromosomie, zwłaszcza u izolatów klinicznych [23]. Gen qacB występuje na plazmidach oporności na sole metali ciężkich. Oporność na antyseptyki i środki dezynfekcyjne, takie jak cetrymid, chlorheksydyna, chlorek benzalkoniowy wynika z budowy 14-segmentowej proteiny QacA, która w pozycji 323 transmembranowego segmentu 10 ma naładowaną ujemną resztę aminokwasową zdolną do rozpoznania i przyłączenia kationu CSA. Efektem takiego wiązania jest wzrost zależnego od pompy QacA eksportu środków bakteriobójczych z komórki bakterii i brak wrażliwości na tego typu związki [49].
Ponadto wykazano również, że fenotyp plejotropowej oporności u drożdży Saccharomyces cerevisiae może wynikać ze zwiększonej ekspresji genów np. PDR5, SNQ2 i YOR1, które są pod kontrolą czynników transkrypcyjnych Pdr1p i Pdr3p. Tak np. zwiększona ekspresja genu PDR5 nadaje oporność na cykloheksymid, chloramfenikol, antymycynę, fungicydy, herbicydy, leki antynowotworowe [34,35]. Również produkt genu SGE1 powoduje oporność drożdży na bromek etydyny i fiolet krystaliczny. U drożdży głównymi składnikami błony komórkowej jest ergosterol i fosfolipidy. Ich wzajemne oddziaływania wpływają na płynność i asymetrię błony, która kontroluje transport do komórki [91]. Wykazano, że oporność może być związana ze zmianą lipidów błony komórkowej [15,30].
Do poznania mechanizmów oporności, oprócz badań genetycznych, niezbędna jest również znajomość warunkujących ją procesów biochemicznych. Wiadomo, że gen musi być najpierw transkrybowany na mRNA, który z kolei przeniesiony na rybosomy ulega translacji, czego produktem jest białko. Następstwem mutacji w genie może być modyfikacja kodowanego przez ten gen białka. Tak zmodyfikowane białko może tracić powinowactwo do fungicydu, w wyniku czego nie jest on przez komórkę pobierany lub nie łączy się z jej strukturami. Istnieje też możliwość powstawania za pośrednictwem mutacji enzymu rozkładającego fungicyd. Wreszcie, komórka oporna może całkowicie tracić zdolność syntezy białka wykazującego powinowactwo z fungicydem. Główne mechanizmy rozwoju oporności mogą polegać na:
• modyfikacji miejsca wrażliwego enzymu tak, że fungicyd traci powinowactwo do białka;
• uruchomieniu alternatywnej drogi biosyntezy, pozwalającej na ominięcie szlaku zablokowanego przez fungicyd;
• zmniejszeniu pobierania fungicydu przez komórkę (zmiana składu kompozycji błony) lub zwiększeniu jego intensywności wydalania (ABC i MFS transportery);
• włączenie cząsteczki fungicydu w szlaki metaboliczne komórki, prowadzące do jego inaktywacji;
• zmniejszenie intensywności konwersji związku nietoksycznego w toksyczny;
• amplifikacji genu, co powoduje wzrost ilości danego enzymu, na który działa inhibitor;
• zmianie wewnątrzkomórkowego pH, które może być przyczyną zmiany struktury chemicznej fungicydu [21,23,35,50,90]. W zależności od rodzaju organizmu, oporność na konkretny związek może opierać się o różne, opisane wyżej mechanizmy.
PIŚMIENNICTWO
[1] Abe S., Kaneda T.: Effect of betaines and taurine and its derivates on plasma cholesterol levels in rats. Eiyo To Shokuryo, 1975; 28: 125-128
[2] Agarwal J.S., Rastogi R.P.: Chemical examination of the water-soluble fraction of Mappia foetida. Indian J. Chem., 1975; 13: 758-759
[3] Anthoni U., Christophersen C., Hougard L., Nielsen P.H.: Quaternary ammonium compounds in the biosphere-an example of a versatile adaptive strategy. Comp. Biochem. Physiol., 1991; 99B: 1-18
[4] Anthoni U., Nielsen P.H., Smith-Hansen L., Wium-Anderson S., Christophersen C.: Charamin a quaternary ammonium ion antibiotic from the green-alga Chara globularis. J. Org. Chem., 1987; 52: 694-695
[5] Breton M., Wolf C., Colard O.: Linoleate incorporation into rat liver membranes phospholipids: effect on plasma membrane ATPase activities and physical properties. Biochem. Biophys. Res. Commun., 1983; 117: 809-816
[PubMed]
[6] Broda T., Przestalski S.: The effect of biological active substances in the red blood cell membranes. Post. Biol. Kom., 1984, 11: 525-526
[7] Carson R.T., Larson E., Levy S.B., Marshall B.M., Aiello A.E.: Use of antibacterial consumer products containing quaternary ammonium compounds and drug resistance in the community. J. Antimicrob. Chemother., 2008; 62: 1160-1162
[PubMed] [Full Text HTML] [Full Text PDF]
[8] Colley C.M., Zwaal R.F., Roelofsen B., van Deenen L.L.: Lytic and non-lytic degradation of phospholipids in mammalian erythrocytes by pure phospholipases. Biochim. Biophys. Acta, 1973, 307: 74-82
[PubMed]
[9] Csonka L.N.: Physiological and genetic responses of bacteria to osmotic stress. Microbiol. Rev., 1989; 53: 121-147
[PubMed] [Full Text PDF]
[10] de la Maza A., Parra J.L.: Solubilizing effects caused by the nonionic surfactant dodecylmaltoside in phosphatidylcholine liposomes. Biophys. J., 1997; 72: 1668-1675
[PubMed] [Full Text PDF]
[11] Debbasch C., de Saint Jean M., Pisella P.J., Rat P., Warnet J.M., Baudouin C.: Quaternary ammonium cytotoxity in a human conjunctival cell line. J. Fr. Ophtalmol., 1999; 22: 950-958
[PubMed] [Full Text HTML] [Full Text PDF]
[12] Denyer S.P.: Mechanism of action of antibacterial biocides. Int. Biodeterior. Biodegrad., 1995; 36: 227-245
[13] Dickson D.M., Kirst G.O. Osmotic adjustment in marine eukaryotic algae: The role of inorganic ions, quaternary ammonium, tertiary sulfonium and carbohydrate solutes. I. Diatoms and a rhodophyte. New Phytol., 1987, 106: 645-655
[14] Domagk G.: Eine neue klasse von desinfektionsmitteln. Dtsch. Med. Wochenschr., 1935; 61: 829-832
[Abstract]
[15] Dubnicková M., Rezanka T., Koscová H.: Adaptive changes in fatty acids of E. coli strains exposed to a quaternary ammonium salt and an amine oxide. Folia Microbiol., 2006; 51: 371-374
[PubMed]
[16] Dubois J.W., Ruddick J.N.: The fungal degradation of quaternary ammonium compounds in wood. International Research Group (IRG) on Wood Protection, 1998
[17] Dufour J.P., Goffeau A.: Molecular and kinetic properties of the purified plasma membrane ATPase of the yeast Schizosaccharomyces pombe. Eur. J. Biochem., 1980; 105: 145-154
[PubMed] [Full Text PDF]
[18] Gabryjelska J., Grobelny D., Kuczera J., Przestalski S., Witek S., Zyłka R.: Effect of some benzylammonium chlorides of sulpathe and calcium ion transport across liposome membrane. Stud. Biophys., 1979; 77: 193-200
[19] Gabryjelska J., Kuczera J., Oświęcimska M., Przestalski S., Witek S., Żyłka R.: Effect of alkyl chain length in alkoxymethylene trimethylammonium chlorides on ion transport across liposome membranes. Stud. Biophys., 1981; 82: 149-155
[20] Gaffey M.J., Hayden F.G., Boyd J.C., Gwaltney J.M. Jr.: Ipratropium bromide treatment of experimental rhinovirus infection. Antimicrob. Agents Chemother., 1988; 32: 1644-1647
[PubMed] [Full Text PDF]
[21] Ghannoum M.A., Rice L.B.: Antifungal agents: mode of action, mechanisms of resistance, and correlation of these mechanisms with bacterial resistance. Clin. Microbiol. Rev., 1999; 12: 501-517
[PubMed] [Full Text HTML] [Full Text PDF]
[22] Gilbert P.: Microbial resistance to preservative systems. W: Microbial quality assurance in pharmaceuticals, cosmetics and toiletries. Red. S.F. Bloomfield, R. Baird, R.E. Leak, R. Leech. Ellis Horwood, Chichester, England, 1988, 171-194
[23] Gillespie M.T., May W., Skurray R.A.: Plasmid-encoded resistance to acriflavine and qarternary ammonium compounds in methicillin-resistant Staphylococcus aureus. FEMS Microbiol. Lett., 1986; 34: 47-51
[24] Giolando S., Rapaport R.A., Larson R.J.: Environmental fate and effects of DEEDMAC: A new rapidly biodegradable cationic surfactant for use in fabric softeners. Chemosphere, 1995; 30: 1067-1083
[25] Grant W.M.: A study of the actions of nonaromatic quaternary ammonium compounds on the eye. Trans. Am. Ophthalmol. Soc., 1956; 54: 417-451
[PubMed] [Full Text PDF]
[26] Guérin-Méchin L., Leveau J.Y., Dubois-Brissonnet F.: Resistance of spheroplasts and whole cells of Pseudomonas aeruginosa to bactericidal activity of various biocides: evidence of the membrane implication. Microbiol. Res., 2004; 159: 51-57
[PubMed]
[27] Gupta A.K., Ahmad I., Summerbell R.C.: Fungicidal activities of commonly used disinfectants and antifungal pharmaceutical spray preparations against clinical strains of Aspergillus and Candida species. Med. Mycol., 2002; 40: 201-208
[PubMed]
[28] Helenius A., Simons K.: Solubilization of membranes by detergents. Bioch. Biophys. Acta, 1975; 415: 29-79
[PubMed]
[29] Hiom S.J., Furr J.R., Russel A.D., Dickinson J.R.: Effects of chlorhexidine diacetate and cetylpyridinium chloride on whole cells and protoplasts of Saccharomyces cerevisiae. Microbios, 1993; 74: 111-120
[PubMed]
[30] Kawai M., Yamada S., Ishidoshiro A., Oyamada Y., Ito H., Yamagishi J.: Cell-wall thickness: possible mechanism of acriflavine resistance in meticillin-resistant Staphylococcus aureus. J. Med. Microbiol., 2009; 58: 331-336
[PubMed] [Full Text HTML] [Full Text PDF]
[31] Kleszczyńska H., Matyjasik S., Sarapuk J., Grobelny D., Witek S.: Interaction of some quaternary ammonium salts with red cells and planar lipid membranes. Studia Biophys., 1981; 84: 173-178
[32] Kleszczyńska H., Sarapuk J., Dziamska A.: The physicochemical properties of some new aminophosphonates. Cell. Mol. Biol. Lett., 2000; 5: 415-422
[Abstract] [Full Text PDF]
[33] Kłosowska-Wołkowicz Z., Królikowski W., Penczek P.: Żywice i laminaty poliestrowe. WNT, Warszawa, 1986
[34] Kołaczkowska A., Kołaczkowski M., Goffeau A., Moye-Rowley W.S.: Compensatory activation of the multidrug transporters Pdr5p, Snq2p, and Yor1p by Pdr1p in Saccharomyces cerevisiae. FEBS Lett., 2008; 582: 977-983
[PubMed] [Full Text PDF]
[35] Kołaczkowski M., Kołaczkowska A., Łuczyński J., Witek S., Goffeau A.: In vivo characterization of the drug resistance profile of the major ABC transporters and other components of the yeast pleiotropic drug resistance network. Microb. Drug Resist., 1998; 4: 143-158
[PubMed]
[36] Kołodyński J., Ułaszewski S., Grobelny D., Witkowska R., Witek S., Lachowicz T.: Effect of some quaternary benzylammonium salts on physiology of yeast. Acta Microbiol. Pol., 1984; 33: 119-130
[PubMed]
[37] Koyama K., Shimazu Y.: Benzalkonium chlorides. Drugs and Poisons in Humans, 2005; 23: 407-413
[38] Kuczera J., Fogt A., Żyłka R., Oświęcimska R., Witek S.: Wpływ wybranych biologicznie aktywnych pochodnych estrów glicyny na błony fosfolipidowe. Zeszyty Nauk. Akad. Rol. Wrocław, 1985; 152
[39] Kuczera J., Sarapuk J., Janas T., Witek S., Grobelny D.: Effects of some cyclic elements containing amphiphilic compounds on stability and transport properties of model lecithin membranes. Gen. Physiol. Biophys., 1987; 6: 645-654
[PubMed]
[40] Kyte J.: Properties of the two polypeptides of sodium- and potassium-dependent adenosine triphosphatase. J. Biol. Chem., 1972; 247: 7642-7649
[PubMed] [Full Text PDF]
[41] Lachowicz T.M., Obłąk E., Piątkowski J.: Auxotropy -stimulated sensitivity to quaternary ammonium salt and its relation to active transport in yeast. Bul. Pol. Acad. Sci., 1992; 40: 173-182
[42] Lachowicz T.M., Piątkowski J., Witek S.: Quaternary ammonium salts and arginine are inhibitors of general amino acid permease in yeast. Pestic. Sci., 1995; 43: 169-171
[43] Lachowicz T.M., Witkowska R., Obłąk E.: Amino acid auxotrophy increases sensitivity of Saccharomyces cerevisiae to a quaternary ammonium salt IM. Acta Microbiol. Pol., 1990; 39: 157-162
[PubMed]
[44] Lee C.: Structure, conformation, and action of neuromuscular blocking drugs. Br. J. Anaesth., 2001; 87: 755-769
[PubMed] [Full Text HTML] [Full Text PDF]
[45] Lichtenberg D., Robson R.J., Dennis E.A.: Solubilization of phospholipids by detergents. Structural and kinetic aspects. Biochim. Biophys. Acta, 1983; 737: 285-304
[PubMed]
[46] Makino S., Reynolds J.A., Tanford C.: The binding of deoxycholate and Triton X-100 to proteins. J. Biol. Chem., 1973; 248: 4926-4932
[PubMed] [Full Text PDF]
[47] McDonnell G., Russell A.D.: Antiseptics and disinfectants: activity, action, and resistance. Clin. Microbiol. Rev., 1999; 12: 147-179
[PubMed] [Full Text HTML] [Full Text PDF]
[48] McGregor D.O., Dellow W.J., Robson R.A., Lever M., George P.M., Chambers S.T.: Betaine supplementation decreases post-methionine hyperhomocysteinemia in chronic renal failure. Kidney Int., 2002; 61: 1040-1046
[PubMed] [Full Text HTML] [Full Text PDF]
[49] Mitchell B.A., Brown M.H., Skurray R.A.: QacA multidrug efflux pump from Staphylococcus aureus: comparative analysis of resistance to diamidines, biguanidines, and guanylhydrazones. Antimicrob. Agents Chemother., 1998; 42: 475-477
[PubMed] [Full Text HTML] [Full Text PDF]
[50] Mitchell B.A., Paulsen I.T., Brown M.H., Skurray R.A.: Bioenergetics of the staphylococcal multidrug export protein QacA. Identification of distinct binding sites for monovalent and divalent cations. J. Biol. Chem., 1999; 274: 3541-3548
[PubMed] [Full Text HTML] [Full Text PDF]
[51] Netherton J.C.3rd, Gurin S.: Biosynthesis and physiological role of homarine in marine shrimp. J. Biol. Chem., 1982; 257: 11971-11975
[PubMed] [Full Text PDF]
[52] Nobile S., Deshusses J.: Evidence for a role of a vicinal dithiol in the transport of γ-butyrobetaine in Agrobacterium sp. Biochimie, 1988; 70: 1411-1416
[PubMed]
[53] Nozaki Y., Reynolds J.A., Tanford C.: The interaction of a cationic detergent with bovine serum albumin and other proteins. J. Biol. Chem., 1974; 249: 4452-4459
[PubMed] [Full Text PDF]
[54] Obłąk E., Adamski R., Lachowicz T.M.: pH-dependent influence of a quaternary ammonium salt and an aminoester on the yeast Saccharomyces cerevisiae ultrastructure. Cell. Mol. Biol. Lett., 2003; 8: 105-110
[PubMed]
[55] Obłąk E., Bącal J., Lachowicz T.M.: A quaternary ammonium salt as an inhibitor of plasma membrane H+-ATPase in yeast Saccharomyces cerevisiae. Cell. Mol. Biol. Lett., 2000; 5: 315-324
[Full Text PDF]
[56] Obłąk E., Gamian A., Adamski R., Ułaszewski S.: The physiological and morphological phenotype of a yeast mutant resistant to the quaternary ammonium salt N-(dodecyloxycarboxymethyl)-N,N,N-trimethyl ammonium chloride. Cell. Mol. Biol. Lett., 2010; 15: 215-233
[PubMed]
[57] Obłąk E., Lachowicz T.M., Łuczyński J., Witek S.: Comparative studies of the biological activities of lysosomotropic aminoesters and quaternary ammonium salts on the yeast Saccharomyces cerevisiae. Cell. Mol. Biol. Lett., 2001; 6: 871-880
[PubMed]
[58] Obłąk E., Lachowicz T.M., Łuczyński J., Witek S.: Lysosomotropic N,N-dimethyl α-aminoacid n-alkyl esters and their quaternary ammonium salts as plasma membrane and mitochondrial ATPases inhibitors. Cell. Mol. Biol. Lett., 2002; 7: 1121-1129
[PubMed]
[59] Obłąk E., Lachowicz T.M., Witek S.: DL-leucine transport in a Saccharomyces cerevisiae mutant resistant to quaternary ammonium salts. Folia Microbiol., 1996; 41: 116-118
[PubMed]
[60] Obłąk E., Ułaszewski S., Lachowicz T.M.: Mutants of Saccharomyces cerevisiae resistant to a quaternary ammonium salt. Acta Microbiol. Pol., 1988; 37: 261-270
[PubMed]
[61] Obłąk E., Ułaszewski S., Morawiecki A., Witek S., Witkowska R., Majcher K., Lachowicz T. M.: Quaternary ammonium salt-resistant mutants in yeast Saccharomyces cerevisiae. Yeast, 1989; 5 (Suppl. 1): S273-S277
[PubMed]
[62] Obłąk E., Witkowska R., Witek S., Lachowicz T.M.: Respiratory deficiency and sensitivity to a quaternary ammonium salt in yeast Saccharomyces cerevisiae. W: Genetic of respiratory enzymes in yeast. Wrocław University Press 1990: 96-102
[63] Ohkawa K., Kim H., Lee K.: Biodegradation of Electrospun Poly(e-caprolactone) Non-woven Fabrics by Pure-Cultured Soil Filamentous Fungi. J. Polym. Env., 2004; 12: 211-218
[Abstract]
[64] Petrocci A.N.: Surface-active agents: quaternary ammonium compounds. W: S.S. Block (red.): Disinfection, sterilization and preservation, Lea and Febiger. Philadelpha, 1983; 309-329
[65] Prince D.L., Prince H.N., Thraenhart O., Muchmore E., Bonder E., Pugh J.: Methodological approaches to disinfection of human hepatitis B virus. J. Clin. Microbiol., 1993; 31: 3296-3304
[PubMed] [Full Text PDF]
[66] Raghavendra T.: Neuromuscular blocking drugs: discovery and development. J. R. Soc. Med., 2002; 95: 363-367
[PubMed] [Full Text HTML] [Full Text PDF]
[67] Rietschel R.L.: Fisher’s Contact Dermatitis. Williams& Wilkins, Maryland, USA 1995
[68] Rodrigo G., Rodrigo C., Burschtin O.: A meta-analysis of the effects of ipratropium bromide in adults with acute asthma. Am. J. Med., 1999; 107: 363-370
[PubMed]
[69] Rogers B., Decottignies A., Kolaczkowski M., Carvajal E., Balzi E., Goffeau A.: The pleitropic drug ABC transporters from Saccharomyces cerevisiae. J. Mol. Microbiol. Biotechnol., 2001; 3: 207-214
[PubMed] [Full Text PDF]
[70] Roll D.M., Ireland C.M., Lu H.S., Clardy J.: Fascaplysin, an unusual antimicrobial pigment from the marine sponge Fascaplysinopsis sp. J. Org. Chem., 1988; 53: 3276-3278
[Abstract] [Full Text PDF]
[71] Różycka-Roszak B., Przyczyna A.: Interaction between N-dodecyl-N,N-dimethyl-N-benzylammonium halides and phosphatidylcholine bilayers – the effect of counterions. Chem. Phys. Lipids, 2003; 123: 209-221
[PubMed]
[72] Różycka-Roszak B., Żyłka R.: The interaction of amphiphilic quatenary ammonium salts with model membranes: the counterion effect. Cell. Mol. Biol. Lett., 2002; 7 (Suppl. 1): 215
[73] Rucka M., Oświęcimska M., Witek S.: New biocides for cooling water treatment. III. Quaternary ammonium salts derivatives of glycine esters. Environ. Prot. Eng., 1983; 9: 25-31
[74] Russell A.D.: Bacterial spores and chemical sporicidal agents. Clin. Microbiol. Rev., 1990; 3: 99-119
[PubMed] [Full Text PDF]
[75] Salton M.R.: Lytic agents, cell permeability, and monolayer penetrability. J. Gen. Physiol., 1968; 52: 227-252
[PubMed]
[76] Sato A., Fenical W.: Gramine-derived bromo-alkaloids from the marine bryozoan Zoobotryon verticillatum. Tetrahedr. Lett., 1983; 24: 481-484
[Abstract]
[77] Scholte H.R., Luyt-Houwen I.E., Vaandrager-Verduin M.H.: The role of the carnitine system in myocardial fatty acid oxidation: carnitine deficiency, failing mitochondria and cardiomyopathy. Basic Res. Cardiol., 1987; 82 (Suppl. 1): 63-73
[PubMed]
[78] Serrano R., Montesinos C., Sanchez J.: Lipid requirement of the plasma membrane ATPases from oat roots and yeast. Plant Sci., 1988; 56: 117-122
[79] Shirai A., Sumitomo T., Kurimoto M., Maseda H., Kourai H.: The mode of the antifungal activity of gemini-pyridinium salt against yeast. Biocontrol Sci., 2009; 14: 13-20
[PubMed]
[80] Siatra-Papastaikoudi T., Papadaki-Valiraki A., Tsantili-Kakoulidou A., Tzouvelekis L., Mentis A.: Synthesis, lipophilicity studies and antibacterial properties of some novel quaternary ammonium salts. Chem. Pharm. Bull., 1994; 42: 392-394
[PubMed]
[81] Skała J., Orłowska-Matuszewska G., Misiewicz M., Oświęcimska M., Witek S., Kotylak Z.: Biological effect of alkoxymethylene trimethylammonium chlorides on yeast Saccharomyces cerevisiae. Acta Microbiol. Polon., 1988; 37: 271-280
[PubMed]
[82] Smith L.T., Pocard J.A., Bernard T., Le Rudulier D.: Osmotic control of glycine betaine biosynthesis and degradation in Rhizobium meliloti. J. Bacteriol., 1988; 170: 3142-3149
[PubMed] [Full Text PDF]
[83] Springthorpe V.S., Sattar S.A.: Chemical disinfection of virus-contaminated surfaces. Crit. Rev. Environ. Sci. Technol., 1990; 20: 169-229
[Abstract]
[84] Subczyński W.K., Przestalski S., Kuczera J., Podolak M., Hyde J.S.: Effect of some amphiphilic ammonium salts on phase transition and fluidity of phosphatidilcholine membranes: a spin label study. Studia Biophys., 1988; 125: 155-163
[85] Takácsová G., Subik J.: Antimicrobial and cytolytic activity of N,N-dimethyl-1-methyldodecylamine oxide. Folia Microbiol., 1979; 24: 153-156
[PubMed]
[86] Trivedi A., Khare S., Singhal G.S., Prasad R.: Effect of phosphatidylcholine and phosphatidylethanolamine enrichment on the structure and function of yeast membrane. Biochim. Biophys. Acta, 1982; 692: 202-209
[PubMed]
[87] Waditee R., Bhuiyan M.N., Rai V., Aoki K., Tanaka Y., Hibino T., Suzuki S., Takano J., Jagendorf A.T., Takabe T., Takabe T.: Genes for direct methylation of glycine provide high levels of glycinebetaine and abiotic-stress tolerance in Synechococcus and Arabidopsis. Proc. Natl. Acad. Sci. USA, 2005; 102: 1318-1323
[PubMed] [Full Text HTML] [Full Text PDF]
[88] Witek S., Goffeau A., Nader J., Łuczyński J., Lachowicz T.M., Kuta B., Obłąk E.: Lysosomotropic aminoesters act as H+-ATPase inhibitors in yeast. Folia Microbiol., 1997; 42: 252-254
[PubMed]
[89] Xiao Y.H., Chen J.H., Fang M., Xing X.D., Wang H., Wang Y.J., Li F.: Antibacterial effects of three experimental quaternary ammonium salt (QAS) monomers on bacteria associated with oral infections. J. Oral Sci., 2008; 50: 323-327
[PubMed] [Full Text PDF]
[90] Yamada N., Promden W., Yamane K., Tamagake H., Hibino T., Tanaka Y., Takabe T.: Preferential accumulation of betaine uncoupled to choline monooxygenase in young leaves of sugar beet – importance of long-distance translocation of betaine under normal and salt-stressed conditions. J. Plant Physiol., 2009; 166: 2058-2070
[PubMed]
[91] Yang Y.L., Lo H.J.: Mechanisms of antifungal agent resistance. J. Microbiol. Immunol. Infect., 2001; 34: 79-86
[PubMed]
[92] Zhu X.Y., Wu J.M., Jia Z.S.: Superoxide anion radical scavenging ability of quaternary ammonium salt of chitosan. Chin. Chem. Lett., 2004; 15: 808-810
[Full Text PDF]
Autorzy deklarują brak potencjalnych konfliktów interesów.