
Rozpoznawanie patogenów przez wrodzony system odporności
Marta Sochocka 1Streszczenie
W pracy omówiono udział komórek NK (natural killer) i komórek dendrytycznych (DC – dendritic cell), receptorów PRRs (pathogen recognition receptors), struktur wykrywanych przez nie na powierzchni patogenów (PAMPs – pathogen associated molecular patterns) oraz udział procesów wewnątrzkomórkowego przekazywania sygnału we wrodzonej odporności. Przedstawiono również możliwe uczestnictwo we wrodzonej odporności nowo opisanej grupy komórek IKDCs (interferon-producing killer DCs), wykazujących aktywność zarówno NK, jak i DC. Opisano mechanizmy wrodzonej odporności z możliwym udziałem procesu autofagii, uruchamianym wskutek aktywacji endosomalnych receptorów TLR7 (Toll-like receptor 7). Autofagia pozwala na eliminację wewnątrzkomórkowych bakterii i wirusów przez stymulację wytwarzania dużych ilości interferonów typu I (type I IFNs). Równowaga między aktywacją i hamowaniem działania receptorów PRR, zwłaszcza TLR, jest warunkiem prawidłowego uruchamiania odpowiedzi immunologicznej i uniknięcia rozwoju chorób o podłożu autoimmunologicznym czy zakaźnym. Oprócz udziału cząsteczek, takich jak SOCS (suppressor of cytokine signaling) czy białka Tollip (Tollinteracting protein) przedstawiono możliwy udział mikroRNA w procesach regulacji wrodzonej odporności. Przełomowe odkrycie tych małych, niekodujących odcinków RNA znacznie zmieniło pogląd na regulację ekspresji wielu różnych genów.
Słowa kluczowe:odporność wrodzona • cytotoksyczność NK • TLR • autofagia
Summary
The contributions of NK (natural killer) cells, DCs (dendritic cells), PRRs (pathogen recognition receptors), and PAMPs (pathogen-associated molecular patterns) recognized by PRRs as well as the participation of intracellular signaling pathways in innate immunity are shown. The possible involvement of a newly described group of immune cells, IKDCs (interferon-producing DCs), in innate immunity which show both NK and DC activity is presented. This review describes the mechanisms of innate immunity with the likely participation of autophagy initiated after endosomal TLR7 (Toll-like receptor 7) activation. Autophagy is able to remove intracellular bacteria or viruses by stimulating type I IFNs (interferons). A balance between PRR, especially TLR, activation and inactivation is crucial to avoid an excessive inflammatory response, as occurs in autoimmune and infectious diseases. Apart from SOCS (suppressor of cytokine signaling proteins) or Tollip (Toll-interacting protein), the possible contribution of microRNA in the regulation of innate immunity is described. The discovery of miRNA is one of the major scientific milestones in recent years and has revolutionized the way we look at many different modes of gene regulation.
Key words:innate immunity • NK cytotoxicity • TLRs • autophagy
ODPORNOŚĆ WRODZONA
Układ odpornościowy każdego organizmu ma skutecznie chronić go przed różnego rodzaju zagrożeniami ze strony środowiska zewnętrznego, zwłaszcza natury infekcyjnej. Mechanizmy działania układu immunologicznego polegają na sprawnej identyfikacji, a następnie likwidacji różnych czynników chorobotwórczych, tolerując własne żywe i zdrowe komórki i tkanki. Proces wykrywania patogenów jest bardzo trudny, ponieważ im bardziej komplikują się sposoby obrony organizmu, tym bardziej patogeny udoskonalają swoje strategie. W tym celu organizmy żywe przez miliony lat ewolucji kształtowały zróżnicowane mechanizmy defensywne, określane dziś jako nieswoiste (wrodzone, naturalne) i swoiste (nabyte) reakcje układu immunologicznego. Pierwszą linię obrony, wspomagającą odporność nieswoistą, stanowią naturalne bariery organizmu w postaci skóry i błon śluzowych oraz wydzielanych przez nie substancji bakteriobójczych i bakteriostatycznych (lizozym, defensyny). Skuteczną ochronę warunkuje także niskie pH skóry i żołądka, a także naturalna flora bakteryjna i wytwarzane przez nią substancje. Przełamanie przez patogeny barier fizjologicznych skutkuje uruchomieniem bardziej skomplikowanych mechanizmów obronnych organizmu. Są to reakcje wrodzonego systemu odpornościowego, poprzedzające bezpośrednio rozwój reakcji swoistych. Odporność wrodzona jest filogenetycznie starszą gałęzią układu immunologicznego, obecną u wszystkich organizmów wielokomórkowych (Metazoa). Począwszy od gąbek aż do człowieka zaobserwować można wielką różnorodność receptorów oraz mechanizmów efektorowych, wspólnych niekiedy także z królestwem roślin [28].
Odporność wrodzona selektywnie i natychmiast rozróżnia między „obcy–swój”, nie wymagając uprzedniego kontaktu z antygenem. Nie pozostawia długotrwałej odporności przeciw patogenowi w postaci komórek pamięci immunologicznej, ale bardzo często wystarcza do całkowitej eliminacji zakażenia. Komórki wrodzonego systemu odporności ssaków, takie jak makrofagi, komórki dendrytyczne (DC) czy NK mają niezmienne, nabyte w ewolucji, receptory rozpoznające konserwatywne, ważne dla przetrwania struktury różnych grup mikroorganizmów i przekazujące sygnał o zakażeniu. Sygnały te mają za zadanie zainicjować kaskadę stanu zapalnego, związaną głównie z wytwarzaniem różnorodnych cytokin prozapalnych (TNF-α, IL-1,-6) oraz czynników chemotaktycznych, które pomagają zwalczać zakażenie oraz aktywować odpowiedź swoistą – drugą gałąź systemu immunologicznego [63,85]. Odporność nabyta polega na ekspansji różnych subpopulacji limfocytów, skierowanych przeciw patogenom. Jest to proces wysoce efektywny i o dużej swoistości, ale do pełnego rozwoju potrzebuje nawet kilku dni. Prezentacja antygenu limfocytom, a następnie ich selekcja klonalna prowadzi do wzmocnienia odpowiedzi i wytworzenia swoistych przeciwciał. W przypadku infekcji wszystkie charakterystyczne objawy stanu zapalnego (zaczerwienienie, opuchlizna, gorączka, ból, utrata prawidłowych czynności) są zapoczątkowywane przez odporność wrodzoną. Komórki odporności swoistej mogą wspierać i zaostrzać te efekty, jednak zasadniczy sygnał, który rozpoczyna i ostatecznie decyduje o odpowiedzi uzależniony jest od identyfikacji patogenu przez elementy odpowiedzi nieswoistej [9].
ZNACZENIE KOMÓREK NK I DC WE WRODZONEJ ODPORNOŚCI
Komórki NK (natural killer) charakteryzują się zdolnością do naturalnej, niewymagającej wstępnej aktywacji, cytotoksyczności. Większość z nich wywołuje lizę komórek docelowych przez wydzielanie perforyny i granzymów [51,85]. NK są odpowiedzialne również za tzw. nadzór immunologiczny. Brak bądź obniżona ekspresja na powierzchni kontrolowanych komórek antygenów zgodności tkankowej MHC klasy I, powoduje natychmiast aktywację NK. Komórki NK1, świeżo izolowane, wytwarzają IFN-γ, przy braku IL-4, -5 oraz -13. Natomiast NK2, nieuwalniające IFN-γ, wytwarzają duże ilości IL-4, -5 oraz -13. Wśród komórek NK wyróżnia się także dwie inne subpopulacje komórek: CD56dim/CD16+ (CD56–/+CD16++) oraz CD56bright/CD16– (CD56++CD16–/+). Wyznacznikiem podziału jest intensywność ekspresji adhezyjnej molekuły powierzchniowej N-CAM (neural cell adhesion molecule), czyli CD56. Grupy te różnią się między sobą rozmieszczeniem, zdolnościami do zabijania, receptorami oraz ilością, ale nie profilem wytwarzanych cytokin. Cytotoksyczność natomiast przypisuje się jedynie komórkom CD56dim [32,51]. Komórki NK ulegają aktywacji wskutek integracji sygnałów z aktywujących i hamujących receptorów, rozpoznających konkretne komponenty na powierzchni komórki. NK stanowią pierwszą linię obrony przeciw patogenom wirusowym, coraz bardziej wzrasta również ich znaczenie w odpowiedzi immunologicznej na zakażenia bakteryjne i pasożytnicze. [5,37,44,54,95]. Ponadto biorą udział w eliminacji komórek transformowanych, powstrzymując progresję nowotworu. Należy natomiast pamiętać, iż naturalna aktywność komórek NK jest różna w zależności od zaawansowania choroby i często zmniejsza się w stanach terminalnych z przerzutami [58]. Regulacja reaktywności i funkcji efektorowych NK zależy bezpośrednio od sygnałów wysyłanych przez receptory aktywujące i hamujące, występuje jednak również na etapie dojrzałych, krążących komórek w odpowiedzi na cytokiny. Interferony typu 1 oraz cytokiny, takie jak IL-2 czy IL-15 wzmacniają reaktywność NK [15]. Ludzkie komórki NK mogą także zabijać docelowe komórki opłaszczone przez przeciwciała w reakcji cytotoksyczności ADCC (antibody-dependent cellular cytotoxicity). W procesie tym główną rolę odgrywa receptor aktywujący FcγRIIIA, obecny na wszystkich typach NK. Ostatnie doniesienia wskazują jednak na udział w tej aktywności receptora hamującego FcγRIIB, obecnego jedynie na niewielkiej, nowo odkrytej subpopulacji NK- CD56+/NKp46+ [30].
Cytotoksyczność NK zależy głównie od obecnych we wnętrzu tych komórek charakterystycznych ziaren azurofilnych, zawierających lityczne białka, takie jak perforyna, białko TIA-1 (poly(A)-binding protein), NKCF (NK cytotoxic factor) i proteazy serynowe, określane jako granzymy i granulizyna. Uwolniona perforyna wbudowuje się i tworzy kanały w błonie komórek docelowych. Efektem końcowym jej działania jest uruchomienie apoptozy w komórce docelowej, dzięki wniknięciu jonów wapnia oraz proteaz serynowych i białka TIA-1 i degradacja DNA. W procesie tym biorą także udział granzymy, które wnikają do komórki docelowej przez pory utworzone przez perforynę i nabywają właściwości proteolitycznych. Granzym B trawi niektóre kaspazy, co przekształca je w cząsteczki uczestniczące w apoptozie [51]. Ponadto stwierdzono, że granzym K (GzmK) może nie tylko indukować szybką śmierć komórki, ale również wytwarzanie reaktywnych form tlenu (ROS – reactive oxygen species) [36]. Sygnał aktywujący komórki NK jest wynikiem oddziaływania receptorów, takich jak NKG2D czy NKp46 z odpowiednimi ligandami. Może być również wywołana działaniem chemokin MIP-1α czy IP-10 lub interleukin. Wśród interleukin największe znaczenie ma IL-2, która stymuluje proliferację NK i wzmaga działanie cytotoksyczne. Oprócz IL-2 aktywność NK stymulują także IL-12, -18, -21, a także IFN-α, -β i -γ [24,79,103,107]. Ponadto ważnym fizjologicznym czynnikiem wzrostu jest IL-15. Jest ona odpowiedzialna za różnicowanie i cytolityczną aktywność dojrzałych NK. Zhang i wsp. [114] wykazali, iż IL-15 zwiększa transkrypcję i ekspresję NKG2D na powierzchni NK, wytwarzanie cząsteczek cytotoksycznych (TRAIL i perforyny), a także fosforylację białek STAT1 (signal transducers and activator of transcription1) i ERK1/2 (extracellular signalregulated kinases). Wpływa to bezpośrednio na lityczne właściwości komórek NK i może mieć znaczenie w immunoterapii nowotworów. Interesujące wydają się również rezultaty Krzewskiego i wsp. [53], dotyczące białka WIP (Wiskott-Aldrich syndrome protein [WASp] – interacting protein). Wykazali oni, że białko WIP uczestniczy w tworzeniu dużych kompleksów białkowych zaangażowanych w rearanżację cytoszkieletu aktynowego NK, co jest niezbędne w procesie cytotoksyczności NK. WIP zostało zlokalizowane w otoczeniu litycznych substancji ziaren azurofilnych. Brak białka WIP powoduje blokadę polaryzacji komórki NK, co świadczy o tym, że jest ono zaangażowane w transport substancji granularnych i pełni ważną rolę w regulacji cytotoksycznych funkcji komórek NK. Aktywowane komórki NK są efektywniejsze, jeśli chodzi o działanie cytotoksyczne i jest to także związane z wytwarzaniem przez nie dużych ilości IFN-γ i TNF [103,116]. TNF sam może działać cytotoksycznie, zaś IFN-γ może pobudzać m.in. makrofagi. Obie cytokiny oddziałują ponadto na komórki prezentujące antygen (APC – antygen presenting cells) w miejscu zakażenia, co sprzyja rozwojowi swoistych mechanizmów immunologicznych. Badania ostatnich lat wykazują ogromny postęp w walce z różnymi nowotworami z użyciem komórek NK. Obecnie nowo opracowana metoda nukleofekcji z użyciem białka EGFP (enhanced green fluorescent protein) lub lucyferazy pozwala na skuteczny transfer genów do komórek NK bez użycia wektora wirusowego. Dzięki temu wzrasta cytotoksyczność komórek NK, co może się stać skutecznym narzędziem w immunoterapii nowotworów oraz infekcji wirusowych [78].
Oprócz komórek NK we wrodzonej odporności uczestniczą również komórki dendrytyczne (DCs – dendritic cells). Podobnie jak NK są grupą funkcjonalnie zróżnicowaną, różne subpopulacje DC odgrywają odmienną rolę we wrodzonej odporności. Mieloidalna frakcja DC (mDC – myeloid DC), nazywana inaczej DC1 lub komórkami prezentującymi antygeny typu I (APC1 – antygen presenting cell type 1), o fenotypie CD11c, CD1c, jest odpowiedzialna głównie za pochłanianie antygenów oraz aktywację limfocytów T. Druga grupa, plazmocytoidalna (pDC – plasmocytoid DC), nazywana również DC2 lub APC2, o fenotypie CD123+, BDCA2, BDCA4, wpływa na proces prezentacji antygenów przez wydzielanie dużych ilości IFN-a. Właśnie ze względu na wytwarzanie bardzo dużych ilości interferonu typu I (IFN type I), 1000 razy więcej niż inne komórki, ale także interferonu typu III (IFN type III), niedojrzałe pDC określane są również jako komórki NIPCs (natural interferon producing cells) – naturalnie wytwarzające interferon. pDC występujące we krwi obwodowej oraz tkance limfatycznej wydzielają IFN-α w odpowiedzi na DNA oraz RNA wirusów, poprzez sygnał z endosomalnych receptorów TLR7 i TLR9 [33,74].
Cechą charakterystyczną cyklu życiowego komórek DC jest ich transformacja ze stadium niedojrzałych do postaci dojrzałych DC. Niedojrzałe DC, rezydujące w tkankach są zdolne do wykrywania antygenów i endocytozy, a po wędrówce do węzłów chłonnych stają się już profesjonalnymi komórkami APC. Słowo „dojrzałe” w tym przypadku niekoniecznie musi oznaczać „całkowicie zdolne” do prezentacji antygenów. Czasem wymagają jeszcze dodatkowego sygnału do aktywacji. Zgodnie z tą teorią zwaną „paradygmatem komórek Langerhansa” (Langerhans cell paradigm), klasyczny cykl życiowy DC polega na dojrzewaniu DC po zetknięciu z komponentami patogenu, takimi jak ligandy TLR (np. LPS) lub mediatorami stanu zapalnego (np. TNF-α) [34,109]. Obecnie wiadomo już, że komórki DC są grupą heterogenną, ze względu na pochodzenie (mieloidalne i plazmocytoidalne), umiejscowienie w tkankach (obwodowe, tkankowe, węzłów chłonnych) oraz cykl życiowy i nie wszystkie zachowują się zgodnie z proponowanym paradygmatem komórek Langerhansa. Badania Wilsona i wsp. [109] na myszach pozbawionych genów cząsteczek adaptorowych MyD88 i TRIF wykazały, iż DC mogą migrować z tkanek obwodowych i dojrzewać mimo braku obecności jawnego zakażenia. Oznacza to, że komórki DC mogą dojrzewać w odpowiedzi na kontakt z endogennymi mediatorami stanu zapalnego w kontekście MHC II, podobnie jak w przypadku aktywacji przez receptory TLR. Proporcje i ekspresja markerów dojrzewania na migrujących DC w węzłach chłonnych była podobna jak u myszy normalnych. Wyniki te sugerują, iż migracja DC oraz dojrzewanie, podlega regulacji w stanie równowagi dynamicznej przez wrodzony mechanizm różnicowania DC, lub przez uwalnianie małych ilości sygnałów zapalnych z prawidłowych tkanek. Innym, nowo opisanym czynnikiem dojrzewania DC jest talaktoferryna (TLF), rekombinowana ludzka laktoferryna o właściwościach silnie przeciwnowotworowych. Po aktywacji TLF komórki DC charakteryzują się m.in. zwiększonym wydzielaniem IL-8 i ekspresją białka CXCL10, natomiast zdecydowanie spada w nich wytwarzanie IL-6 i -10 oraz ekspresja CCL20. Zmniejsza się również ich zdolność do pochłaniania antygenów, natomiast wzrasta zdolność do proliferacji i uwalniania IFN-γ w obecności alogenicznych ludzkich limfocytów T [88]. Ostatnie doniesienia mówią także o udziale DC i makrofagów w regulacji autoreaktywnych limfocytów B podczas wrodzonej odpowiedzi immunologicznej. DC i makrofagi powstrzymują zmienione komórki B od wytwarzania autoprzeciwciał przez wydzielanie IL-6 oraz rozpuszczalnej postaci CD40L (sCD40L). Mechanizm ten jest selektywny, ponieważ w warunkach prawidłowych, przy braku właściwości autoimmunologicznych, IL-6 oraz sCD40L nie hamują wytwarzania przeciwciał przez limfocyty B. Przyczyną licznych chorób autoimmunologicznych, np. tocznia układowego, jest defekt w tolerancji DC i makrofagów wobec limfocytów B. Rekonstrukcja prawidłowych właściwości tych komórek mogłaby się stać alternatywną metodą terapii immunosupresyjnych [104].
Niezwykle ważne dla rozwoju odpowiedzi immunologicznej jest kooperacja komórek NK i DC (ryc.1). Bezpośredni kontakt NK z mDC powoduje aktywację NK i dojrzewanie DC. DC mogą też wzmagać proliferację NK. Interakcja ta jest zależna od działania IL-2, -12, -15, -18 czy -4 uwalnianych w miejscu, gdzie rozwija się odpowiedź zapalna z udziałem NK.

Ryc. 1. Współpraca między komórkami NK i DC w kształtowaniu odpowiedzi immunologicznej (wg [25] zmodyfikowano)
Agaugue i wsp. [1] wykazali, iż krótkotrwała ekspozycja NK na działanie tych cytokin promuje następnie odmienne drogi pobudzania komórek T. W warunkach działania IL-2 czy IL-12 na NK dochodzi do dojrzewania DC, zdolnych do pobudzania Th1, które wydzielają IFN-γ. Jednak ekspozycja NK na IL-18 skutkuje indukcją polaryzacji Th1. W tej sytuacji IL-2 wydzielana przez limfocyty T oraz IL-12 pochodząca z DC wciąż promuje wydzielanie IFN-γ przez komórki T. Działanie IL-4 na NK nie polaryzuje komórek T, wydzielana jest też bardzo mała ilość IL-2. Aktywowane komórki NK są w stanie zabić DC, jeśli jest ich około 5 razy więcej, natomiast proporcja odwrotna, w obecności IFN-γ i TNF, powoduje gwałtowne dojrzewanie DC i ich oporność na zabijanie przez NK. Do kontaktu między oboma typami komórek dochodzi w tkankach obwodowych i w węzłach chłonnych, gdzie są rekrutowane przez chemokiny uwalniane we wczesnej fazie rozwoju odpowiedzi zapalnej. Różne subpopulacje NK: CD56dim/CD16+ oraz CD56bright/CD16– różnią się między sobą sposobem działania i lokalizacją. Komórki CD56bright/CD16– zdecydowanie dominują w węzłach chłonnych, podczas gdy CD56dim/CD16+ są w większości obecne we krwi obwodowej. Obie subpopulacje wykazują ponadto funkcjonalne różnice w aktywności cytolitycznej, wytwarzaniu cytokin i procesie proliferacji [31,67]. Komórkowe i molekularne mechanizmy kontrolujące interakcje między NK i DC są jednak wciąż słabo poznane. Najnowsze badania wskazują na udział różnych białek, takich jak kompleks Jagged2- Notch czy chemokiny CX3CL1 w procesie aktywacji i proliferacji NK przez DC, wzmacniania ich cytotoksyczności oraz wydzielania IFN-γ [49,71].
Komórki NK i DC są głównymi składnikami wrodzonej odporności. Niedawno opisano jednak trzecią linię komórek IKDCs (interferon-producing killer DCs), izolowanych z puli komórek śledziony myszy, odrębną od plazmocytoidalnych DC i konwencjonalnych DC (cDC – conventional DC) mających profil molekularny zarówno NK jak i DC. Pierwsze doniesienia wskazywały, iż komórki te charakteryzują się zdolnością do wytwarzania znacznych ilości IFN typu pierwszego oraz IL-12 lub IFN-γ w zależności od stymulatora aktywacji, przez co wykazywały cytotoksyczność identyczną z NK. Ponadto po ustaniu działania cytotoksycznego nabywały one zdolności do prezentacji antygenów w kontekście MHC II [20]. Późniejsze badania zweryfikowały nieco te doniesienia. Vremec i wsp. [106] wykazali, iż IKDCs są potencjalnymi producentami IFN-γ, jednak nie wytwarzają zupełnie IFN-a w odpowiedzi na stymulację niemetylowanymi sekwencjami CpG, co świadczyło o bliższym pokrewieństwie z NK niż DC. Wyniki te potwierdzili również Caminschi i wsp. [18] dowodząc, iż IKDCs są bardzo podobne w swoich litycznych funkcjach do NK, jednak nie są klasycznymi APC jak DC, nawet po aktywacji. Dodatkowo wykazali, iż profil wytwarzanych czynników transkrypcyjnych oraz odpowiedź na cytokiny IKDCs są podobne do NK, ale nie do DCs. Badania te, podobnie jak Vosshenricha i wsp. [105], sugerują zatem, iż IKDCs powinny być raczej uważane bardziej za subpopulację NK niż DCs. Najnowsze doniesienia dotyczą ustalenia dokładnego profilu markerów powierzchniowych IKDCs, wskazują także, że ilość oraz funkcja IKDCs mogą być związane z działaniem IL-15 [4,13]. Wykazano także ogromne znaczenie IKDCs w hamowaniu rozwoju nowotworu, w odpowiedzi na poli I:C. Kwas poli I:C promuje ekspansję IKDCs oraz wytwarzanie przez nie IFN-γ, co wzmacnia ich właściwości cytolityczne względem komórek nowotworowych. Badania te wyjaśniają również dlaczego poli I:C może funkcjonować jako adiuwant, poprawiający przeciwnowotworowe działanie komórek wrodzonego systemu odporności [43].
RECEPTORY I SZLAKI PRZESYŁANIA SYGNAŁU WRODZONEGO SYSTEMU ODPORNOŚCI
Głównym mechanizmem uruchamiającym stan zapalny w organizmie jest rozpoznanie patogenu przez receptory wrodzonego systemu odporności. Ogólna zasada rozpoznawania drobnoustrojów polega na identyfikacji takich struktur w ich budowie, które są strategicznie istotne do ich przeżycia. Cząsteczki te nazwano molekularnymi strukturami związanymi z patogenami (PAMPs – pathogen associated molecular patterns), a receptory je rozpoznające PRRs (pattern recognition receptors) [110]. Receptory PRR mogą funkcjonować jako:
• receptory błonowe (np. receptor mannozowy [MR – mannose receptor], dektynowy 1, receptory zmiatacze [SRs – scavenger receptors]);
• rozpuszczalne białka zaangażowane w opsonizację i aktywację dopełniacza (np. białko wiążące mannozę [MBL – mannose binding lectin]);
• receptory aktywujące komórki (receptory Toll-podobne [TLRs – Toll-like receptors], receptory na komórkach NK) [9,11,80,99].
Receptory TLR opisano pod koniec ubiegłego wieku. Wiadomo było, iż komórki odporności wrodzonej, takie jak makrofagi czy komórki dendrytyczne, są aktywowane przez różne struktury mikroorganizmów, np. LPS bakterii Gram-ujemnych, swoiste receptory odpowiedzialne za identyfikację LPS, a także pozostałych struktur, przez długi czas pozostawały niezidentyfikowane [3]. Gen Toll odkryto na początku lat osiemdziesiątych ub.w. przez niemieckich badaczy Nusslein-Volharda i Wieschausa w genomie muszki owocowej (Drosophila melanogaster). Okazało się, że jest on odpowiedzialny za grzbietowobrzuszną polaryzację owada w rozwoju embrionalnym. Dopiero w 1996 r. Toll opisano, jako główny receptor u Drosophila aktywujący efektywną odpowiedź przeciw Aspergillus fumigatus. Gen Toll koduje transmembranowe białko, homologiczne do receptora interleukiny 1(IL-1R), Toll/IL-1R (TIR) [3,11]. Kolejne badania wykazały, że odkryty ssaczy homolog receptora Toll, Toll-like receptor 4 (TLR-4), indukuje wytwarzanie cytokin prozapalnych. Doświadczenie na myszach pozbawionych genu TLR4, traktowanych LPS wykazało całkowity brak odporności u zwierząt na podany antygen. Rezultaty te stały się motorem napędowym dalszego rozwoju badań nad wrodzoną odpornością. Zidentyfikowano wiele podobnych genów i ich produktów u bezkręgowców, roślin i ssaków, u tych ostatnich ze względu na podobieństwo zostały one nazwane Toll-podobnymi (Toll-like). Receptory TLR są ewolucyjnie konserwatywnymi białkami. U ludzi opisano 11 różnych TLR (u innych ssaków 13), z których każdy ma swoją wewnętrzną drogę sygnałowania indukującą swoistą odpowiedź biologiczną przeciw różnym mikroorganizmom [3,102]. W obrębie receptorów TLR zidentyfikowano liczne polimorfizmy, które jak się sugeruje mogą być związane z podatnością do rozwoju wielu chorób, takich jak sepsa czy zakażenia Mycobacterium tuberculosis lub M. leprae [66]. Część cytoplazmatyczna receptorów TLR nazywana domeną TIR (Toll/IL-1R) odpowiada za aktywację i przekazywanie wewnątrzkomórkowego sygnału aktywacji. Zewnątrzkomórkowy region zawiera domeny bogate w leucynę (LRRs – leucyne-rich repeats), które są odpowiedzialne za rozpoznawanie różnych struktur na powierzchni patogenów. Interakcja ta powoduje dojrzewanie DC i wytwarzanie cytokin, co w rezultacie aktywuje rozwój odpowiedzi swoistej [96]. TLR występują na powierzchni m.in. fagocytów (makrofagi, neutrofile, DC), NK, NKT, limfocytach B, T, na komórkach nabłonkowych jelit i dróg oddechowych, ale także w błonach pęcherzyków cytoplazmatycznych ulegających fuzji z fagolizosomami [3,9,70]. TLR występują również na powierzchni komórek tucznych, które wskutek aktywacji są zdolne do fagocytozy adherentnych bakterii, uwalniania prozapalnych mediatorów bez rozległej degranulacji, a następnie prezentacji antygenów limfocytom T [41]. Poszczególne TLR rozpoznają różne PAMPs indukując odpowiedź immunologiczną (ryc.2).

Ryc. 2. Receptory TLRs oraz rozpoznawane przez nie PAMPs (wg [45] zmodyfikowano)
BAKTERIE
Głównym i silnym immunostymulatorem jest lipopolisacharyd (LPS), składnik ściany bakterii Gram-ujemnych [3,90]. W zależności od pochodzenia LPS stymuluje TLR4 lub TLR2, a to z kolei decyduje o rodzaju indukowanej odpowiedzi nabytej. Odpowiedź Th1- bądź Th2- zależna jest uruchamiana jako bezpośrednie następstwo działania LPS na komórki DC [57]. Lipopeptydy, peptydoglikan bakterii Gram-dodatnich i ujemnych, zymosan, lipoarabinomannan mikobakterii oraz kwas lipotejchojowy bakterii Gram-dodatnich są rozpoznawane przez TLR2, który jest zaangażowany również w identyfikację nietypowych postaci LPS nonenterobakterii [50,72,102]. Receptory TLR1 i TLR6 strukturalnie bardzo przypominają TLR2. Heterodimery TLR2 i TLR1 lub TLR6 rozróżniają molekularne struktury diacylo- i triacylolipopeptydów [10]. Bakteryjne białko flagellina, składnik wici, jest rozpoznawane przez TLR5. Receptor ten występuje na komórkach epitelialnych przewodu pokarmowego i jest głównym detektorem bakterii chorobotwórczych wnikających przez epitelium [7,102]. Kolejnym potencjalnym stymulatorem odpowiedzi immunologicznej jest bakteryjne DNA, zawierające niemetylowane motywy CpG. Badania na myszach bez TLR9 wykazały, że jest on właśnie receptorem CpG DNA [102]. Dotąd nie zidentyfikowano jednak ligandów TLR10, być może są to białka bakterii uropatogennych [98].
GRZYBY
Receptory TLR są zaangażowane w rozpoznawanie patogenów grzybiczych, takich jak: Aspergillus fumigatus, Candida albicans, Cryptococcus neoformans czy Pneumocystis carrini. Zymosan jest rozpoznawany przez heterodimer TLR2/TLR6, podczas gdy mannan pochodzący od Saccharomyces cerevisiae, wykrywany jest przez TLR4. Ponadto pochodne mannanu, takie jak fosfolipomannan czy glukuronoksylomannan również aktywują TLR2 i TLR4 [52]. W rozpoznawaniu grzybiczych PAMPs z receptorem TLR2 funkcjonalnie współpracuje receptor Dectin 1, dla którego ligandem jest β-glukan [64,80].
PROTOZOA
Różne komponenty pasożytów, takich jak Trypanosoma cruzi, Toxoplazma gondii, Plasmodium falciparum i inne, również są wykrywane przez receptory wrodzonego systemu odporności. Glikofosfatydyloinozytol (GPI), glikoinositolofosfolipidy czy genomowe DNA – pochodzące z T. cruzi są odpowiednio rozpoznawane przez TLR2, TLR4 i TLR9 [94]. Podczas gdy TLR9 rozpoznaje także hemozoinę wytwarzaną przez P. falciparum, profilinopodobne białko Toxoplazma profilin czy T. gondii jest ligandem dla TLR11 [73,97].
WIRUSY
Badania na myszach z mutacją w TLR4 wykazały ich dużą wrażliwość na zakażenie RSV (respiratory syncytial virus). Pozwoliło to wnioskować, iż receptor ten rozpoznaje nie tylko komponenty bakteryjne, ale także białka osłonek wirusów [84,101]. Struktury wirusów, human herpesvirus1 (HHV-1), cytomegalovirus (CMV) czy hepatitis C virus (HCV), wykrywane są także przez TLR2 [20,68,81]. Jednak głównymi stymulatorami odpowiedzi immunologicznej przeciw wirusom są dwuniciowy RNA (dsRNA), jego syntetyczny analog, kwas poliinozyno-policytydylowy (poli I:C), rozpoznawane przez TLR3 [62,113], niemetylowane motywy CpG DNA, imidazokwinolinopodobne cząsteczki oraz jednoniciowy RNA (ssRNA) rozpoznawane przez TLR7, TLR8 i TLR9, obecne głównie w przedziałach wewnątrzkomórkowych (fagocytosomach) [47,92,102]. Za mechanizm indukujący wytwarzanie IFN w odpowiedzi na zakażenie VSV (vesicular stomatitis virus) i HHV-1 uznano stosunkowo niedawno autofagię, samożerność (autophagy). Jest to filogenetycznie stary proces występujący we wszystkich komórkach eukariotycznych, nazywany także II typem programowanej śmierci komórki, mniej selektywny polegający na degradacji białek i innych organelli w lizosomach. Autofagia jest uruchamiana w odpowiedzi na braki składników odżywczych podczas głodu komórkowego, niedobór tlenu, uszkodzenia spowodowane przez toksyny komórkowe, oraz wskutek działania czynników indukujących rozwój i różnicowanie [27,89]. Autofagia, uznana jako nowy mechanizm odporności wrodzonej, jest również komórkowo autonomicznym systemem eliminacji patogenów wewnątrzkomórkowych. Formowanie się autofagosomów pełni istotną rolę w rozpoznawaniu niektórych wirusów i wzmaganiu wytwarzania IFN przez pDC. Badania wskazują, że autofagia może być indukowana przez niektóre ligandy TLR. Najbardziej widoczny efekt zauważono w przypadku stymulacji TLR 7 przez ssRNA. Inne ligandy tego receptora również aktywowały autofagię, co wykazało zupełnie nowe możliwości oddziaływania tych cząsteczek. Jednak dokładne sygnały i drogi sygnałowania uruchamiające mechanizm samożerności w odpowiedzi na patogeny pozostają nieznane [26].
Ostatnie doniesienia sugerują, że istnieje także mechanizm wykrywający replikujące się wirusy w cytoplazmie w sposób niezależny od TLR, prowadzący do aktywacji czynnika interferonu – IRF (interferon release factor) oraz czynnika jądrowego κB (NF-κB – nuclear factor κB). W procesie tym biorą udział cytoplazmatyczne receptory RIG-I (retinolic acid inducible gene-I) rozpoznający dsRNA wirusów oraz MDA5 (melanoma differentiation – associated gene 5) wiążący białko V paramyksowirusów [22,93]. W rozpoznawaniu wirusowego DNA bierze natomiast udział receptor DAI (DNA-dependent activator of IFN-regulatory factor) nazywany też ZBP1 (Z-DNA binding protein 1), który wzmacnia indukcję wytwarzania interferonów typu pierwszego oraz ekspresję innych genów wrodzonej odporności [91].
Niemal wszystkie TLR mają podobną drogę sygnałowania, w której cząsteczki adaptorowe (MyD88, TIRAP/MAL, TRIF, TRAM) tworzą molekularny kompleks z domeną TIR uruchamiając w ten sposób kaskadę sygnału (ryc.3). Każdy TLR aktywuje charakterystyczną odpowiedź poprzez inną kombinację TIR-TLR i cząsteczki adaptorowej. Sygnał może być przekazywany zależnie lub niezależnie od MyD88 [3,46,102]. Aktywacja receptora rozpoczynająca kaskadę sygnałową prowadzi z kolei do powstania aktywnej postaci jądrowego czynnika transkrypcyjnego NF-κB, który przemieszcza się z regionu cytoplazmatycznego do jądra. Tu wiąże się do promotorów genów dla różnych cytokin, chemokin i cząsteczek adhezyjnych (TNF-α, IL-1, -6) inicjując w ten sposób sygnał do rozwoju odpowiedzi swoistej.
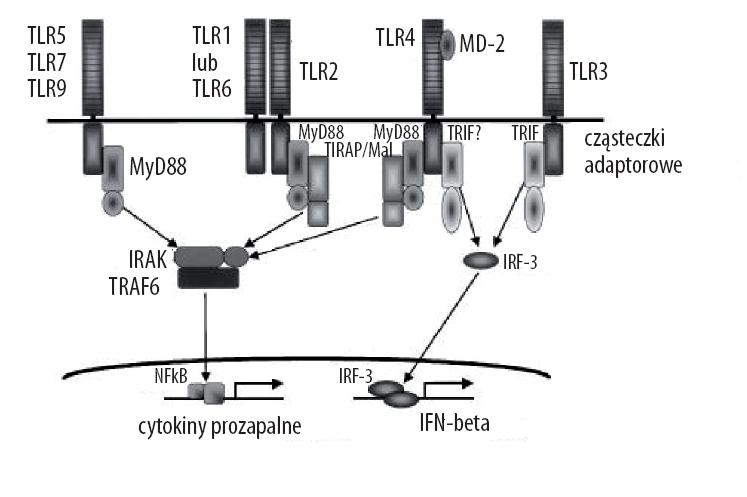
Ryc. 3. Drogi sygnałowe TLRs (wg [2] zmodyfikowano)
Poszczególne TLR aktywują ponadto alternatywne drogi z udziałem kinaz IKK-zależnych TBK1 (TRAF – family member-associated NF-kappaB activator (TANK) binding kinase) oraz IKKi, które uruchamiają przeciwwirusową wrodzoną odpowiedź systemu immunologicznego [48]. Najnowsze doniesienia Rubstova i wsp. [75] mówią także o udziale TLR w aktywacji komórek B i wspomagania odpowiedzi IgM zależnej od limfocytów T. Agoniści TLR powodują aktywację i mobilizację limfocytów B strefy brzeżnej (MZ – marginal zone) śledziony oraz jako adiuwanty komórek T, wzmacniają antygenowozależną odpowiedź IgM. Wzmacniają także wczesne wytwarzanie swoistych IgM oraz aktywację komórek B i pobudzają ich przemieszczanie się poza MZ.
Rozpoznawanie patogenów poprzez TLR zawsze indukuje wytwarzanie cytokin prozapalnych. Równowaga między aktywacją i inaktywacją TLR jest więc istotna w celu uniknięcia nadmiernej odpowiedzi zapalnej, która może prowadzić do rozwoju chorób o podłożu autoimmunologicznym, przewlekłych chorób zapalnych czy zakaźnych. Mutacje w genach białek sygnałowych mogą być odpowiedzialne za pierwotne niedobory odporności [12]. Badania potwierdzają m.in. rozwijanie się zapalenia mózgu w przewlekłej infekcji HHV-1 u chorych z deficytem TLR-3 [115]. Drogi aktywacji TLR mogą być kontrolowane przez wiele zewnątrz- i wewnątrzkomórkowych cząsteczek, utrzymujących równowagę między zdrowiem i chorobą. Blokowanie sygnałów z TLR może następować poprzez aktywność receptorów wabiących (decoy receptors), takich jak SIGIRR (single immunoglobulin IL-1R-related molecule) czy RP105 (CD180), rozpuszczalnych postaci TLR (sTLR – soluble TLR) oraz inhibitorów cytoplazmatycznych i jądrowych (SOCS, IRAK-M, NOD2, Tollip i in.), hamujących sygnał na różnych etapach aktywacji [82]. Czynnikami zewnątrzkomórkowymi są przede wszystkim rozpuszczalne postaci TLR, obecne w ludzkiej surowicy czy mleku, które regulują odpowiedź prozapalną i uruchamiają odporność wrodzoną [3]. Ich działanie jest najczęściej związane z blokowaniem sygnału postaci transbłonowej receptora przez oddziaływanie z jego ligandem. Receptory SIGIRR czy RP105 blokują sygnał TLR poprzez interferencję z „kompleksami sygnałowymi zapoczątkowującymi przekazywanie sygnału”. Endogenne regulatory odpowiedzi prozapalnej to liczne wewnątrzkomórkowe cząsteczki adaptorowe oraz kinazy. Jednym z najważniejszych negatywnych regulatorów są białka SOCS (suppressor of cytokine signaling proteins), indukowane przez cytokiny. Do rodziny białek CIS-SOCS należy 8 cząsteczek (CIS oraz SOCS1-SOCS7), które blokują sygnał inicjowany przez TLR, poprzez degradację i ubikwitynację kompleksów sygnałowych. Białka SOCS1 i SOCS3 hamują wytwarzanie cytokin prozapalnych, szlakiem zależnym od aktywacji NF-kB, poprzez degradację bezpośrednio podjednostki p65 NF-kB, ale także ufosforylowanej cząsteczki adaptorowej MAL [112]. Ponadto aktywowane ligandami TLR blokują również kompleksy kinaz JAK i czynników STAT (JAK-STAT) hamując m.in. indukcję genów IFN [6,23]. SOCS3 hamuje także wytwarzanie IFN przez silne blokowanie STAT3 aktywowanego przez receptor IL-6R, ale nie przez receptor IL-10R, ponieważ nie wiąże się do żadnej podjednostki tego receptora. Mimo niewielkiego wpływu SOCS3 na sygnał z IL-10R z niedawno przeprowadzonych badań wynika, iż długotrwała aktywacja STAT3 może mimo wszystko hamować wytwarzanie cytokin prozapalnych. Mechanizm ten nie jest jeszcze do końca wyjaśniony, jednak wiadomo, że uczestniczy w nim białko (X) indukowane przez STAT3, które jest odpowiedzialne za działanie antyzapalne. Mimo iż SOCS1 jest silnie regulowany przez sygnały z TLR, jednocześnie hamuje GM-CSF (granulocyte- macrophage colony stimulating factor), który stymuluje dojrzewanie komórek dendrytycznych [112]. Innymi adaptorowymi cząsteczkami blokującymi są białko interakcji- Toll [Tollip (Toll-interacting protein)], które blokuje aktywność IL-1 oraz NF-κB. Hamowanie to następuje poprzez supresję autofosforylacji i aktywacji kinaz IRAK. Ponadto obecność Tollip w komórkach epitelialnych układu pokarmowego obniża odpowiedź tych komórek na stymulację LPS i kwasem lipotejchojowym. Redukcji ulega także wytwarzanie IL-6 oraz TNF-α [3,82]. Oprócz udziału we wrodzonej odporności TLR uczestniczy w regulacji odpowiedzi swoistej poprzez stymulację aktywności limfocytów T regulatorowych. Majewska i wsp. [57] podkreślają także rolę TLR w utrzymaniu stanu homeostazy odpowiedzi immunologicznej przez udział w indukcji komórek Treg, a także w przełamaniu stanu tolerancji.
Jednym z przełomów w nauce ostatnich lat, który zrewolucjonizował pogląd na regulację ekspresji genów, było odkrycie mikro RNA (miRNAs), ewolucyjnie konserwatywnych, małych, niekodujących odcinków RNA. Mikro RNA w cytoplazmie łączy się komplementarnie z mRNA, co powoduje degradację mRNA i blokowanie translacji. Każdy miRNA wpływa na ekspresję różnych genów i każdy mRNA jest regulowany przez liczne miRNA. Ponadto miRNA wykazuje tkankowo i ewolucyjnie swoistą ekspresję, co świadczy o tym, iż podlega ona ścisłej regulacji komórkowej [56]. Uważa się, iż miRNA zaangażowany jest m.in. we wrodzoną odporność, ponieważ wpływa na regulację sygnału indukowanego przez patogeny za pośrednictwem receptorów TLR, co w rezultacie prowadzi do regulacji wytwarzania cytokin (miR-146). Antygen miR-181a/miR223 wpływa również na proces zasiedlania komórek immunologicznych oraz mechanizmy odporności nabytej, m.in. na prezentację antygenów (miR-155) czy przekazywanie sygnału przez komórki T (miR-181a) [21,86]. Mikro RNA odgrywa też ważną rolę w obronie przeciwwirusowej oraz chorobach zapalnych, nadmierne wytwarzania miR-203 jest związane np. z rozwojem łuszczycy [35,76,87].
Receptor mannozowy (MR) jest obecny na powierzchni makrofagów, komórek endotelialnych wątroby myszy i niedojrzałych komórkach DC. MR prezentowany jest także na komórkach naczyń limfatycznych, gdzie jego funkcja jest nieznana. Marttila-Ichihara i wsp. [61] wykazali jednak, że u myszy pozbawionych tego receptora następuje redukcja migracji limfocytów ze skóry do węzłów limfatycznych, podczas gdy układ naczyniowy zachowuje prawidłową strukturę. Jest to związane z redukcją właściwości adhezyjnych prawidłowych limfocytów oraz komórek nowotworowych. MR należy do rodziny lektyn typu C, bardzo licznej grupy białek odgrywających ważną rolę we wrodzonej odporności. MR rozpoznaje glikokoniugaty zawierające reszty mannozy, fukozy i N-acetyloglukozaminy. Wśród naturalnych ligandów MR obecnych na powierzchni licznych mikroorganizmów są: polisacharydy, glikoproteiny i glikolipidy. W budowie MR wyróżniamy domenę bogatą w cysteinę (CR – cysteine rich), powtórzenia fibronektynowe typu drugiego (FN II) oraz 8 domen rozpoznających reszty węglowodanowe (CRD-carbohydrate recognition domain) [55]. Funkcją receptora mannozowego jest przede wszystkim udział w rozpoznawaniu różnych PAMP na powierzchni patogenów, aktywacji endocytozy i fagocytozy. Wewnątrzkomórkowe mechanizmy zabijania patogenów przez fagocyty opierają się na procesach tlenowych i beztlenowych. Pobudzone komórki żerne w ciągu kilku sekund przechodzą zmiany prowadzące do powstania reaktywnych form tlenu o właściwościach silnie toksycznych, działających głównie w fagolizosomie. Makrofagi aktywowane IFN-γ, TNF-α czy IL-1 zaczynają także wytwarzać tlenek azotu (NO). NO jest bardzo ważnym mediatorem procesów biologicznych uczestniczących w zabijaniu mikroorganizmów, zwłaszcza po połączeniu z tlenem, kiedy tworzy bardziej toksyczną postać dwutlenku azotu, czy peroksyazotyn (ONOO–) w połączeniu z rodnikiem nadtlenkowym (O2). MR bierze także udział w ograniczaniu wzrostu mikroorganizmów, zwłaszcza grzybów [99,108]. Najczęstsze infekcje grzybicze wywoływane są przez drożdżaki, np. Candida albicans. MR pośredniczy w swoistym wiązaniu N-końcowych reszt mannanu grzyba przez DC, co wpływa jednocześnie na uruchomienie odpowiedzi zapalnej związanej z wytwarzaniem IL-6 [17]. Obecność MR i receptora Dectin 1 rozpoznającego β-glukan w ścianach komórkowych grzybów, wykazano także w mózgu, gdzie oba uczestniczą w fagocytozie patogenów przez komórki mikrogleju i wytwarzaniu reaktywnych form tlenu [80]. Receptor mannozowy, bierze również udział w rozpoznawaniu i eliminacji wirusów. Pośredniczy m.in. w wiązaniu glikoproteiny gp120 i wnikaniu wirusa HIV-1 (human immunodeficiency virus 1) do makrofagów i komórek mikrogleju i w ten sposób przyczynia się do dalszego transferu wirusa. Trujillo i wsp. sugerują również, że może to posłużyć jako punkt zwrotny w poszukiwaniu skutecznej szczepionki anty-HIV [100]. Niedawno Miller i wsp. [65] donieśli, iż MR obecny na powierzchni makrofagów uczestniczy w rozpoznawaniu wszystkich czterech serotypów wirusa dengi (DV – denga virus), wiążąc się do glikoproteiny w osłonce wirusa. Receptor mannozowy jest obecny także w naczyniach limfatycznych, gdzie jego funkcja nie jest do końca wyjaśniona. Wiadomo jednak, iż myszy pozbawione MR wykazywały zmniejszony przepływ limfocytów ze skóry do węzłów chłonnych przez aferentne naczynia limfatyczne. Zauważono także, że u myszy z rozwiniętymi zmianami nowotworowymi, pozbawionych MR, adhezja prawidłowych limfocytów i komórek rakowych do komórek naczyń limfatycznych jest również zmniejszona. Być może blokowanie MR mogłoby się stać nowym celem terapeutycznym w kontroli rozwoju stanu zapalnego i przerzutowania nowotworu [61].
Kolejna grupa receptorów wrodzonej odporności, znana jako receptory scavenger – zmiatacze (SR), występuje na komórkach DC, makrofagach oraz komórkach endotelialnych. Pod względem budowy receptory SR są zgrupowane w trzy odrębne klasy (A, B, C), jednak większość z nich ma domeny kolagenowe, odpowiedzialne za wiązanie struktur bakteryjnych, domeny CR oraz a-helikalną strukturę. SR pełnią istotną rolę w wychwytywaniu i usuwaniu zużytych komponentów komórkowych. Ligandami są dla nich modyfikowane lipoproteiny (LDL – low density lipoprotein i HDL – high density lipoprotein), fosfolipidy, ale również patogeny [40,69]. Receptory SR uczestniczą m.in. w rozpoznawaniu LPS bakterii Gram-ujemnych, kwasu lipotejchojowego bakterii Gram-dodatnich, a także fragmentów CpG DNA. SR uczestniczą również w wiązaniu i usuwaniu komórek apoptotycznych. Ligandami receptora są obecne na powierzchni tych komórek utlenione cząsteczki Ox-LDL [42]. Okazało się również, iż receptor SR-BI bierze udział w rozpoznawaniu, wychwytywaniu i prezentacji antygenów wirusa HCV, co może mieć istotne znaczenie w poszukiwaniu szczepionki i przebiegu celowej, skutecznej immunoterapii przeciwwirusowej [8,77]. Jako receptory LDL stanowią również prozapalny czynnik w przebiegu chorób naczyń krwionośnych, głownie związanych z odkładaniem się blaszek miażdżycowych w tętnicach [29]. Ostatnie doniesienia mówią także o bardzo ważnej roli SR klasy A w transferze antygenów między komórkami B a makrofagami. Receptory BCR zawierające antygen przenoszą go do profesjonalnych APC, jakimi są makrofagi. Blokada SR-A warunkuje prawidłowy przebieg kontaktu między obiema komórkami [39].
Receptory obecne na komórkach NK są odpowiedzialne głównie za indukcję wrodzonej odporności przeciwwirusowej i przeciwnowotworowej. Zarówno blokujące jak i aktywujące receptory NK są obecne na powierzchni obu subpopulacji NK, należą do rodziny białek Ig-podobnych (killer Ig-like receptors – KIR) i lektynopodobnych (lectinlike), takich jak lektynowe receptory cytotoksyczności naturalnej (NCRs – natural cytotoxicity receptors) i receptory lektynowe typu C (KLR – killer lectine-like receptors). Receptory blokujące mają w swojej budowie motywy ITIM (tyrosine-based inhibitory motif), za ich pośrednictwem dochodzi do blokowania reakcji cytotoksycznej komórek NK. Receptory stymulujące natomiast zawierają motywy ITAM (tyrosine-based activatory motif), które wzmagają mechanizmy prowadzące do lizy komórki. Działanie wszystkich receptorów NK jest bezpośrednio związane z funkcjami efektorowymi NK, takimi jak cytotoksyczność czy wytwarzanie cytokin (ryc.4) [14,111].
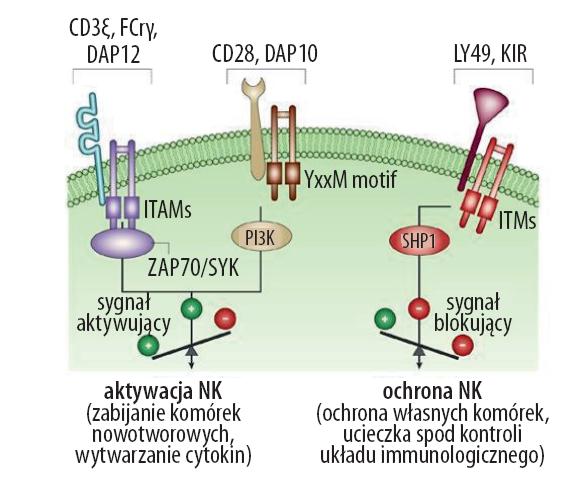
Ryc. 4. Receptory i szlaki sygnałowe NK (wg [83] zmodyfikowano)
Receptory hamujące rozpoznają przede wszystkim różne białka głównego układu zgodności tkankowej (MHC – major histocompatibility complex) klasy I, jednakże niektóre z nich potrafią także blokować zabijanie przez NK w sposób niezależny od MHC I. Blokowanie za pośrednictwem ITIM odbywa się już na bardzo wczesnym etapie aktywacji komórki NK. Fosforylacja tyrozyn w obrębie ITIM aktywuje następnie fosfatazy SHP-1 i SHP-2. To prowadzi do blokady wtórnych mediatorów i ostatecznie zablokowania mechanizmów cytotoksycznych. Wśród receptorów hamujących najlepiej poznaną grupą są KIR (killer cell inhibitory/ immunoglobulin-like receptors) [15,60]. Naturalne receptory cytotoksyczności, czyli aktywujące, obejmujące głównie receptory NKp30, NKp44, NKp46, są odpowiedzialne za rozpoznawanie komponentów wirusowych i działanie przeciwnowotworowe. Jednak do uruchomienia naturalnej cytotoksyczności, czyli degranulacji i wytwarzania cytokin przez NK, zawsze wymagana jest kooperacja w parach receptorów aktywujących. Motywy ITAM w parze z cząsteczką adaptorową DAP12, CD3 lub FcRg aktywują wewnątrzkomórkowe kinazy Syk (ZAP70/SYK), co prowadzi do uruchomienia cytotoksyczności NK. Alternatywna droga aktywacji poprzez receptory CD28 lub NKG2D odbywa się niezależnie od Syk. W tym przypadku poprzez motywy YxxM receptorów następuje połączenie z cząsteczką adaptorową DAP10, a następnie z kinazą PI3K (phosphatidylinositol- 3 kinase), co również uruchamia mechanizmy cytotoksyczności [15]. Zaburzenia w budowie czy funkcjonowaniu kompleksów inicjujących sygnał aktywujący NK (np. CARD9-Bcl10) prowadzą do różnych nieprawidłowości w wytwarzaniu cytokin na szlaku NF-κB [38]. Receptorem aktywującym jest również NKG2D, dla którego ligandami są głównie białka stresu komórkowego MICA, MICB i ULBP [16,59].
PIŚMIENNICTWO
[1] Agaugue S., Marcenaro E., Ferranti B., Moretta L., Moretta A.: Human natural killer cells exposed to IL-2, IL-12, IL-18 or IL-4 differently modulate priming of native T cells by monocyte-derived dendritic cells. Blood, 2008; 112: 1776-1783
[PubMed]
[2] Akira S., Yamamoto M., Takeda K.: Role of adapters in Toll-like receptor signaling. Biochem. Soc. Trans., 2003; 31: 637-642
[PubMed] [Full Text HTML] [Full Text PDF]
[3] Arancibia S.A., Beltrán C.J., Aguirre I.M., Silva P., Peralta A.L., Malinarich F., Hermoso M.A.: Toll-like receptors are key participants in innate immune responses. Biol. Res., 2007; 40: 97-112
[PubMed] [Full Text HTML] [Full Text PDF]
[4] Arina A., Murillo O., Dubrot J., Azpilikueta A., Gabari I., Perez- Gracia J.L., Alfaro C., Berasain C., Prieto J., Ferrini S., Hervas- Stubbs S., Melero I.: Interleukin-15 liver gene transfer increases the number and function of IKDCs and NK cells. Gene Ther., 2008; 15: 473-483
[PubMed]
[5] Athie MoralesV., O’Connor G.M., Gardiner C.M.: Activation of human NK cells by the bacterial pathogen-associated molecular pattern muramyl dipeptide. J. Immunol., 2008; 180: 4082-4089
[PubMed]
[6] Baetz A., Frey M., Heeg K., Dalpke A.H.: Suppressor of cytokine signaling (SOCS) proteins indirectly regulate toll-like receptor signaling in innate immune cells. J. Biol. Chem., 2004; 279: 54708-54715
[PubMed] [Full Text HTML] [Full Text PDF]
[7] Bandyopadhaya A., Sarkar M., Chaudhuri K.: IL-1β expression in Int407 is induced by flagellin of Vibrio cholerae through TLR5 mediated pathway. Microb. Pathog., 2008; 44: 524-536
[PubMed]
[8] Barth H., Schnober E.K., Neumann-Haefelin C., Thumann C., Zeisel M.B., Diepolder H.M., Hu Z., Liang T.J., Blum H.E., Thimme R., Lambotin M., Baumert T.F.: Scavenger receptor class B is required for hepatitis C virus uptake and cross-presentation by human dendritic cells. J. Virol., 2008; 82: 3466-3479
[PubMed] [Full Text HTML] [Full Text PDF]
[9] Barton G.M.: A calculated response: control of inflammation by the innate immune system. J. Clin. Invest., 2008; 118: 413-420
[PubMed] [Full Text HTML] [Full Text PDF]
[10] Bas S., Neff L., Vuillet M., Spenato U., Seya T., Matsumoto M., Gabay C.: The proinflammatory cytokine response to Chlamydia trachomatis elementary bodies in human macrophages is partly mediated by a lipoprotein, the macrophage infectivity potentiator, through TLR2/TLR1/TLR6 and CD14. J. Immunol., 2008; 180: 1158-1168
[PubMed]
[11] Błach- Olszewska Z.: Innate immunity: cells, receptors, and signaling pathways. Arch. Immunol. Ther. Exp., 2005; 53: 245-253
[PubMed]
[12] Błach- Olszewska Z., Leszek J.: Mechanisms of over- activated innate immune system regulation in autoimmune and neurodegenerative disorders. Neuropsychiatr. Dis. Treat., 2007; 3: 365-372
[13] Bonmort M., Dalod M., Mignot G., Ullrich E., Chaput N., Zitvogel L.: Killer dendritic cells: IKDC and the others. Curr. Opin. Immunol., 2008; 20: 558-565
[PubMed]
[14] Brown M.G., Scalzo A.A.: NK gene complex dynamics and selection for NK cell receptors. Semin. Immunol., 2008; 20: 361-368
[PubMed]
[15] Bryceson Y.T., Long E.O.: Line of attack: NK cell specificity and integration of signals. Curr. Opin. Immunol., 2008; 20: 344-352
[PubMed]
[16] Burgess S.J., Maasho M., Naravanan S., Borrego F., Coligan J. E.: The NKG2D receptor: immunobiology and clinical implications. Immunol. Res., 2008; 40: 18-34
[PubMed]
[17] Cambi A., Netea M.G., Mora- Montes H.M., Gow N.A., Hato S.V., Lowman D.W., Kullberg B.J., Torensma R., Williams D.L., Figdor C.G.: Dendritic cell interaction with Candida albicans critically depends on N-linked mannan. J. Biol. Chem., 2008; 283: 20590-20599
[PubMed]
[18] Caminschi I., Ahmet F., Heger K., Brady J., Nutt S.L., Vremec D., Pietersz S., Lahoud M.H., Schofield L., Hansen D.S., O’Keeffe M., Smyth M.J., Bedoui S., Davey G.M., Villadangos J.A., Heath W.R., Shortman K.: Putative IKDCs are functionally and developmentally similar to natural killer cells, but not to dendritic cells. J. Exp. Med., 2007; 204: 2579-2590
[PubMed] [Full Text HTML] [Full Text PDF]
[19] Chan C.W., Crafton E., Fan H.N., Flook J., Yoschimura K., Skarica M., Brockstedt D., Dubensky T.W., Stins M.F., Lanier L.L., Pardoll D.M., Housseau F.: Interferon-producing dendritic cells provide a link between innate and adaptive immunity. Nat. Med., 2006; 12: 207-213
[PubMed]
[20] Chan G., Guilbert L.J.: Ultraviolet- inactivated human cytomegalovirus induces placental syncytiotrophoblast apoptosis in a Toll-like receptor-2 and tumor necrosis factor-alpha dependent manner. J. Pathol., 2006; 210: 111-120
[PubMed]
[21] Chen X.M., Splinter P.L., O’Hara S.P., LaRusso N.F.: A cellular micro-RNA, let-7i, regulates Toll-like receptor 4 expression and contributes to cholangiocyte immune responses against Cryptosporidium parvum infection. J. Biol. Chem., 2007; 282: 28929-28938
[PubMed] [Full Text HTML] [Full Text PDF]
[22] Chi H., Flavell R.A.: Innate recognition of non-self nucleic acids. Genome Biol., 2008; 9: 211
[PubMed]
[23] Dalpke A., Heeg K., Bartz H., Baetz A.: Regulation of innate immunity by suppressor of cytokine signaling (SOCS) proteins. Immunobiology, 2008; 213: 225-235
[PubMed]
[24] de Rham C., Ferrari- Lacraz S., Jendly S., Schneiter G., Dayer J.M., Villard J.: The proinflammatory cytokines IL-2, IL-15 and IL-21 modulate the repertoire of mature human natural killer cell receptors. Arthritis Res. Ther., 2007; 9: R125
[PubMed] [Full Text HTML] [Full Text PDF]
[25] Degli-Esposti M.A., Smyth M.J.: Close encounters of different kinds: dendritic cells and NK cells take centre stage. Nat. Rev. Immunol., 2005; 5: 112-124
[PubMed]
[26] Delgado M.A., Elmaoued R.A., Davis A.S., Kyei G., Deretic V.: Toll-like receptors control autophagy. EMBO J., 2008; 27: 1110-1121
[PubMed]
[27] Deretic V.: Autophagy in innate and adaptive immunity. Trends Immunol., 2005; 26: 523-528
[PubMed]
[28] Du Pasquier L.: Meeting the demand for innate and adaptive immunities during evolution. Scand. J. Immunol., 2005; 62, Supp1: 39-48
[PubMed]
[29] Dunn S., Vohra R.S., Murphy J.E., Homer- Vanniasinkam S., Walker J.H., Ponnambalam S.: The lectin-like oxidized low-density-lipoprotein receptor: a pro-inflammatory factor in vascular disease., 2008; 409: 349-355
[PubMed]
[30] Dutertre C.A., Bonnin- Gélizé E., Pulford K., Bourel D., Fridman W.H., Teillaud J.L.: A novel subset of NK cells expressing high levels of inhibitory FcγRIIB modulating antibody-dependent function. J. Leukoc. Biol., 2008; 84: 1511-1520
[PubMed]
[31] Ferlazzo G.: Natural killer and dendritic cell liaison: recent insights and open questions. Immunol. Lett., 2005; 101: 12-17
[PubMed]
[32] Ferlazzo G.: Isolation and analysis of human natural killer cell subsets. Methods Mol. Biol., 2008; 415: 197-213
[PubMed]
[33] Fitzgerald-Bocarsly P., Dai J., Singh S.: Plasmocytoid dendritic cells and type I IFN: 50 years of convergent history. Cytokine Growth Factor Rev., 2008; 19: 3-19
[PubMed]
[34] Granucci F., Zanoni I., Ricciardi- Castagnoli P.: Central role of dendritic cells in the regulation and deregulation of immune responses. Cell. Moll. Life Sci., 2008; 65: 1683-1697
[PubMed]
[35] Grassmann R., Jeang K.T.: The roles of microRNAs in mammalian virus infection. Biochim. Biophys. Acta, 2008; 1779: 706-711
[PubMed]
[36] Guo Y., Chen J., Zhao T., Fan Z.: Granzyme K degrades the redox/DNA repair enzyme Ape1 to trigger oxidative stress cells leading to cytotoxicity. Mol. Immunol., 2008; 45: 2225-2235
[PubMed]
[37] Han X., Fan Y., Wang S., Jiao L., Qiu H., Yang X.: NK cells contribute to intracellular bacterial infection-mediated inhibition of allergic responses. J. Immunol., 2008; 180: 4621-4628
[PubMed]
[38] Hara H., Ishihara C., Takeuchi A., Xue L., Morris S.W., Penninger J.M., Yoshida H., Saito T.: Cell type-specific regulation of ITAM- mediated NF-kappaB activation by the adaptors, CARMA1 and CARD9. J. Immunol., 2008; 181: 918-930
[PubMed]
[39] Harvey B.P., Quan T.E., Rudenga B.J., Roman R.M., Craft J., Mamula M.J.: Editing antigen presentation: antigen transfer between human B lymphocytes and macrophages mediated by class A scavenger receptors. J. Immunol., 2008; 181: 4043-4051
[PubMed]
[40] Hazen S.L.: Oxidized phospholipids as endogenous pattern recognition ligands in innate immunity. J. Biol. Chem., 2008; 283: 15527-15531
[PubMed]
[41] Heib V., Becker M., Taube C., Stassen M.: Advances in the understanding of mast cell function. Br. J. Hematol., 2008; 142: 683-694
[PubMed]
[42] Jeannin P., Jaillon S., Delneste Y.: Pattern recognition receptors in the immune response against dying cells. Curr. Opin. Immunol., 2008; 20: 530-537
[PubMed]
[43] Jiang Q., Wei H., Tian Z.: IFN-producing killer dendritic cells contribute to the inhibitory effect of poly I:C on the progression of murine melanoma. J. Immunother., 2008; 31: 555-562
[PubMed]
[44] Jonjić S., Babić M., Polić B., Krmpotić A.: immune evasion of natural killer cells by viruses. Curr. Opin. Immunol., 2008; 20: 30-38
[PubMed]
[45] Kawai T., Akira S.: TLR signaling. Cell Death Differ., 2006; 13: 816-825
[PubMed] [Full Text HTML] [Full Text PDF]
[46] Kawai T., Akira S.: TLR signaling. Semin. Immunol., 2007; 19: 24-32
[PubMed]
[47] Kawai T., Akira S.: Antiviral signaling through pattern recognition receptors. J. Biochem., 2007; 141: 137-145
[PubMed]
[48] Kawai T., Akira S.: Signaling to NF-kappaB by Toll-like receptors. Trends Mol. Med., 2007; 13: 460-469
[PubMed]
[49] Kijima M., Yamaguchi T., Ishifune C., Maekawa Y., Koyanagi A., Yagita H., Chiba S., Kishihara K., Shimada M., Yasumoto K.: Dendritic cell-mediated NK cell activation is controlled by Jagged2-Notch interaction. Proc. Natl. Acad. Sci. USA, 2008; 105: 7010-7015
[PubMed] [Full Text HTML] [Full Text PDF]
[50] Knapp S., von Aulock S., Leendertse M., Haslinger I., Draing C., Golenbock D.T., van der Poll T.: Lipoteichoic acid-induced lung inflammation depends on TLR2 and the concerted action of TLR4 and the platelet- activating factor receptor. J. Immunol., 2008; 180: 3478-3484
[PubMed]
[51] Kopeć-Szlęzak J., Podstawka U.: Biologia komórek NK (Natural Killer). Onkol. Pol., 2007; 10/3: 115-119
[Abstract] [Full Text PDF]
[52] Koval’chuk L.V., Khoreva M.V., Varivoda A.S., Pashchenko O.E., Gracheva L.A., Bykova L.P., Kondratenko I.V., Bologov A.A.: Analysis of Toll-like receptor-dependent production of proinflammatory cytokines in vitro by human peripheral blond mononuclears of donors and patients with primary immunodeficiency. Bull. Exp. Biol. Med., 2007; 144: 63-65
[PubMed]
[53] Krzewski K., Chen X., Strominger J.L.: WIP is essential for lytic granule polarization and NK cell cytotoxicity. Proc. Natl. Acad. Sci. USA, 2008; 19,105: 2568-2573
[PubMed] [Full Text HTML] [Full Text PDF]
[54] Lanier L.L.: Evolutionary struggles between NK cells and viruses. Nat. Rev. Immunol., 2008; 8: 259-268
[PubMed]
[55] Linehan S.A.: The mannose receptor is expressed by subsets of APC in non-lymphoid organs. BMC Immunology, 2005; 6: 4
[PubMed] [Full Text HTML] [Full Text PDF]
[56] Lodish H.F., Zhou B., Liu G., Chen C.Z.: Micromanagement of the immune system by microRNAs. Nat. Rev. Immunol., 2008; 8: 120-130
[PubMed]
[57] Majewska M., Szczepanik M.: Rola receptorów toll-podobnych (TLR) w odporności wrodzonej i nabytej oraz ich funkcja w regulacji odpowiedzi immunologicznej. Post. Hig. Med. Dośw., 2006; 60: 52-63
[PubMed] [Full Text HTML] [Full Text PDF]
[58] Malmberg K.J., Bryceson Y.T., Carlsten M., Andersson S., Björklund A., Björkström N.K., Bauman B.C., Fauriat C., Alici E., Dilber M.S., Ljunggren H.G.: NK cell-mediated targeting of human cancer and possibilities for new means of immunotherapy. Cancer. Immunol. Immunother., 2008; 57: 1541-1552
[PubMed]
[59] Mandelboim O.M.: Virus and tumour recognition by the natural cytotoxicity receptors. Immunology, 2004; 113: Supp.1: 14-16
[60] Martinez-Borra J., Khakoo S.I.: Speed and selection in the evolution of killer cell immunoglobulin-like receptors. Int. J. Immunogenet., 2008; 35: 89-96
[PubMed]
[61] Marttila- Ichihara F., Turja R., Miiluniemi M., Karikoski M., Maksimov M., Niemelä J., Martinez- Pomares L., Salmi M., Jalkanen S.: Macrophage mannose receptor on lymphatic controls cell trafficking. Blood, 2008; 112: 64-72
[PubMed]
[62] Matsumoto M., Seya T.: TLR3: interferon induction by double stranded RNA including poly (I:C). Adv. Drug. Deliv. Rev., 2008; 60: 805-812
[PubMed]
[63] Medzhitov R.: Recognition of microorganisms and activation of the immune response. Nature., 2007; 449: 819-826
[PubMed]
[64] Mezger M., Kneitz S., Wozniok I., Kurzai O., Einsele H., Loeffer J.: Proinflammatory response of immature human dendritic cells in mediated by dectin-1 after exposure to Aspergillus fumigatus germ tubes., J. Infect. Dis., 2008; 197: 924-931
[PubMed]
[65] Miller J.L., Dewet B.J., Martinez- Pomares L., Radcliffe C.M., Dwek R.A., Rudd P.M., Gordon S.: The mannose receptor mediates dengue virus infection of macrophages. PLoS. Pathol., 2008; 4: e17
[PubMed] [Full Text HTML] [Full Text PDF]
[66] Misch E.A., Hawn T.R.: Toll-like receptor polymorphisms and susceptibility to human disease. Clinical Science, 2008; 114: 347-360
[PubMed]
[67] Moretta L., Ferlazzo G., Bottino C., Vitale M., Pende D., Mingari M.C., Moretta A.: Effector and regulatory events during natural killer-dendritic cell interactions. Immunol. Rev., 2006; 214: 219-228
[PubMed]
[68] Morrison L.A.: The Toll of herpes simplex virus infection. Trends Microbiol., 2004; 12: 353-356
[PubMed]
[69] Nieland T.J., Shaw J.T., Jaipuri F.A., Duffner J.L., Koehler A.N., Banakos S., Zannis V.I., Kirchhausen T., Krieger M.: Identification of the molecular target of small molecule inhibitors of HDL receptor SR-BI activity. Biochemistry, 2008; 47: 460-472
[PubMed] [Full Text HTML] [Full Text PDF]
[70] Nyman T., Stenmark P., Flodin S., Johansson I., Hammarstrőm M., Nordlund P.: The crystal structure of the human toll-like receptor 10 cytoplasmic domain reveals a putative signaling dimer. J. Biol. Chem., 2008; 283: 11861-11865
[PubMed]
[71] Pallandre J.R., Krzewski K., Bedel R., Ryffel B., Caignard A., Rohrlich P.S., Pivot X., Tiberghien P., Zitvogel L., Strominger J.L., Borg C.: Dendritic cell and natural killer cell cross-talk: a pivotal role of CX3CL1 in NK cytoskeleton organization and activation. Blood, 2008; 112: 4420-4424
[PubMed]
[72] Peiser M., Koeck J., Kirsching C.J., Wittig B., Wanner R.: Human Langerhans cells selectively activated via Toll-like receptor 2 agonists acquire migratory and CD4+ T cell stimulatory capacity. J. Leukoc. Biol., 2008; 83:1118-1127
[PubMed]
[73] Plattner F., Yarovinsky F., Romero S., Didry D., Carlier M.F., Sher A., Soldati-Favre D.: Toxoplasma profiling is essential for host cell invasion and TLR11-dependent induction of an interleukin-12 response. Cell. Host. Microbe., 2008; 3: 77-87
[PubMed]
[74] Randolph G.J., Ochando J., Partida-Sanchez S.: Migration of dendritic cell subsets and their precursors. Annu. Rev. Immunol., 2007; 26: 293-316
[PubMed]
[75] Rubstov A.V., Swanson C.L., Troy S., Strauch P., Pelanda R., Torres R.M.: TLR agonists promote marginal zone B cell activation and facilitate T-dependent IgM responses. J. Immunol., 2008; 180: 3882-3888
[PubMed]
[76] Sall A., Liu Z., Zhang H.M., Yuan J., Lim T., Su Y., Yang D.: MicroRNAs-based therapeutic strategy for virally induced diseases. Curr. Drug. Discov. Technol., 2008; 5: 49-58
[PubMed]
[77] Sawada S., Harada K., Isse K., Sato Y., Sasaki M., Kaizaki Y., Nakanuma Y.: Involvement of Escherichia coli in pathogenesis of xanthogranulomatous cholecystitis with scavenger receptor class A and CXCL16-CXCR6 interaction. Pathol. Int., 2007; 57: 652-653
[PubMed]
[78] Schoenberg K., Trompeter H.I., Uhrberg M.: Delivery of DNA into natural killer cells for immunotherapy. Methods Mol. Biol., 2008; 423: 165-172
[PubMed]
[79] Scott G.B., Meade J.L., Cook G.P.: Profiling killers; unraveling the pathways of human natural killer cell function. Brief. Funct. Genomic. Proteomic., 2008; 7: 8-16
[PubMed]
[80] Shah V.B., Huang Y., Keshwara R., Ozment-Skelton T., Williams D.L., Keshvara L.: β-glucan activates microglia without including cytokine production in Dectin-1-dependent manner. J. Immunol., 2008; 180: 2777-2785
[PubMed]
[81] Shehata M.A., Abou El-Enein A., El-Sharnouby G.A.: Significance of toll-like receptors 2 and 4 mRNA expression in chronic hepatitis C virus infection. Egypt. J. Immunol., 2006; 13: 141-152
[PubMed]
[82] Shibolet O., Podolsky D.K.: TLRs in the Gut. Negative regulation of Toll-like receptors and intestinal homeostasis: addition by subscription. Am. J. Physiol. Gastrointest. Liver Physiol., 2007; 292: G1469-G1473
[PubMed] [Full Text HTML] [Full Text PDF]
[83] Smyth M.J., Hayakawa Y., Takeda K., Yagita H.: New aspects of natural killer cell survivalence and therapy of cancer. Nat. Rev. Cancer, 2002; 2: 850-861
[PubMed]
[84] Snelgrove R., Gwyer E., Hussell T.: Modulation of immunity to respiratory viral infection. Future Virol., 2006; 1: 471-481
[85] Sochocka M., Błach- Olszewska Z.: Mechanizmy wrodzonej odporności. Post. Hig Med Dośw., 2005; 59: 250-258
[PubMed] [Full Text HTML] [Full Text PDF]
[86] Sonkoly E., Stahle M., Pivarcsi A.: MicroRNAs and immunity: novel players in the regulation of normal immune function and inflammation. Semin. Cancer Biol., 2008; 18: 131-140
[PubMed]
[87] Sonkoly E., Stahle M., Pivarcsi A.: MicroRNAs: novel regulators in skin inflammation. Clin. Exp. Dermatol., 2008; 33: 312-315
[PubMed]
[88] Spadaro M., Caorsi C., Ceruti P., Varadhachary A., Forni G., Pericle F., Giovarelli M.: Lactoferrin, a major defense protein of innate immunity, is a novel maturation factor for human dendritic cells. FASEB J., 2008; 22: 2747-2757
[PubMed]
[89] Stępień A., Izdebska M., Grzanka A.: Rodzaje śmierci komórki. Post. Hig. Med. Dośw., 2007; 61: 420-428
[PubMed] [Full Text HTML] [Full Text PDF]
[90] Sun Y., Shu R., Zhang M.Z., Wu A.P.: Toll-like receptor 4 signaling plays a role in triggering periodontal infection. FEMS Immunol. Med. Microbiol., 2008; 52: 362-369
[PubMed]
[91] Takaoka A., Wang Z., Choi M.K., Yanai H., Negishi H., Ban T., Lu Y., Miyagishi M., Kodama T., Honda K., Ohba Y., Taniguchi T.: DAI (DLM-1/ZBP1) is a cytosolic DNA sensor and an activator of innate immune response. Nature, 2007; 448: 501-505
[PubMed]
[92] Takeuchi O., Akira S.: Recognition of viruses by innate immunity. Immunol. Rev., 2007; 220: 214-224
[PubMed]
[93] Takeuchi O., Akira S.: MDA5/RIG-I and virus recognition. Curr. Opin. Immunol., 2008; 20: 17-22
[PubMed]
[94] Tarleton R.L.: Immune system recognition of Trypanosoma cruzi. Curr. Opin. Immunol., 2007; 19: 430-434
[PubMed]
[95] Thomson R.C., Petrik J., Nash A.A., Dutia B.M.: Expansion and activation of NK cell populations in a gammaherpesvirus infection. Scand. J. Immunol., 2008; 67: 489-495
[PubMed]
[96] Tipping P.G.: Toll-like receptors: the interface between innate and adaptive immunity. J. Am. Soc. Nephrol., 2006; 17: 1769-1771
[PubMed] [Full Text HTML] [Full Text PDF]
[97] Togbe D., Schofield L., Grau G.E., Schnyder B., Boissay V., Charron S., Rose S., Beutler B., Quesniaux V.F., Ryffel B.: Murine cerebral malaria development is independent of toll-like receptor signaling. Am. J. Pathol., 2007; 170: 1640-1648
[PubMed] [Full Text HTML] [Full Text PDF]
[98] Tokarz- Deptuła B., Niedźwiedzka P., Deptuła W.: Receptory Toll-podobne – nowe znaczniki w immunologii. Alergia Astma Immunologia, 2006; 11: 23-28
[Abstract] [Full Text PDF]
[99] Torrelles J.B., Azad A.K., Henning L.N., Carlson T.K., Schlesinger L.S.: Role of the C-type lectins in mycobacterial infection. Curr. Drug Targets, 2008; 9: 102-112
[PubMed]
[100] Trujillo J.R., Rogers R., Molina R.M., Dangond F., McLane M.F., Essex M.F., Brain J.D.: Noninfectious entry of HIV-1 into peripheral and brain macrophages mediated by the mannose receptor. Proc. Natl. Acad. Sci. USA, 2007; 104: 5097-5102
[PubMed] [Full Text HTML] [Full Text PDF]
[101] Tulic M.K., Hurrelbrink R.J., Prele C.M., Laing I.A., Upham J.W., Le Souef P., Sly P.D., Holt P.G.: TLR4 polymorphisms mediate impaired responses to respiratory syncytial virus and lipopolisaccharide. J. Immunol., 2007; 179: 132-140
[PubMed] [Full Text HTML] [Full Text PDF]
[102] Uematsu S., Akira S.: Toll- like receptors and innate immunity. J. Mol. Med., 2006; 84: 712-725
[PubMed]
[103] Vahlne G., Becker S., Brodin P., Johansson M.H.: IFN-γ production and degranulation are differentially regulated in response to stimulation in murine natural killer cells. Scand. J. Immunol., 2008; 67: 1-11
[PubMed] [Full Text HTML] [Full Text PDF]
[104] Vilen B.J., Rutan J.A.: The regulation of autoreactive B cells during innate immune response. Immunol. Res., 2008; 41: 295-309
[PubMed]
[105] Vosshenrich C.A., Lesjean- Pottier S., Hasan M., Richard-Le Goff O., Corcuff E., Mandelboim O., Di Santo J.P.: CD11cloB220+ interferon- producing killer dendritic cells are activated natural killer cells. J. Exp. Med., 2007; 204: 2569-2578
[PubMed] [Full Text HTML] [Full Text PDF]
[106] Vremec D., O’Keeffe M., Hochrein H., Fuschberger M., Caminschi I., Lahoud M., Shortman K.: Production of interferons by dendritic cells, plasmocytoid cells, natural killer cells and interferon- producing killer dendritic cells. Blood, 2007; 109: 1165-1173
[PubMed] [Full Text HTML] [Full Text PDF]
[107] Weiss J.M., Subleski J.J., Wigginton J.M., Wiltrout R.H.: Immunotherapy of cancer by IL-12 based cytokine combinations. Expert. Opin. Biol. Ther., 2007; 7: 1705-1721
[PubMed] [Full Text HTML] [Full Text PDF]
[108] Willment J.A., Brown G.D.: C-type lectin receptors in antifungal immunity. Trends. Microbiol., 2008; 16: 27-32
[PubMed]
[109] Wilson N.S., Young L.J., Kupresanin F., Naik S.H., Vremec D., Heath W.R., Akira S., Shortman K., Boyle J., Maraskovsky E., Belz G.T., Villadangos J.A.: Normal proportion and expression of maturation markers in migratory dendritic cells in the absence of germs or Toll-like receptor signaling. Immunol. Cell Biol., 2008; 86: 200-2005
[PubMed]
[110] Yokoyama W., Colonna M.: Innate immunity to pathogens. Curr. Opin. Immunol., 2008; 20: 1-2
[PubMed] [Full Text HTML] [Full Text PDF]
[111] Yokoyama W.M., Riley J.K.: NK cells and their receptors. Reprod. Biomed., 2008; 16: 173-191
[PubMed]
[112] Yoshimura A., Naka T., Kubo M.: SOCS proteins, cytokine signaling and immune regulation. Nat. Rev. Immunol, 2007; 7: 454-465
[PubMed]
[113] Zeytun A., van Velkinburgh J.C., Pardington P.E., Cary R.R., Gupta G.: Pathogen- specific innate immune response. Adv. Exp. Med. Biol., 2007; 598: 342-357
[PubMed]
[114] Zhang C., Zhang J., Niu J., Zhang J., Tian Z.: Interleukin-15 improves cytotoxicity of natural killer cells via up-regulating NKG2D and cytotoxic effector molecule expression as well as STAT1 and ERK1/2 phosphorylation. Cytokine, 2008; 42: 128-136
[PubMed]
[115] Zhang S.Y., Jouanguy E., Sancho-Schimizu V., von Bernuth H., Yang K., Abel L., Picard C., Puel A., Casanova J.L.: Human Toll-like receptor-dependent induction of interferons in protective immunity to viruses. Immunol. Rev., 2007; 220: 225-236
[PubMed]
[116] Zhong M., Kadota Y., Shimizu Y., Gohda E.: Induction of cytolytic activity and interferon-gamma production in murine natural killer cells by polymyxins B and E. Int. Immunopharmacol., 2008; 8: 508-513
[PubMed]