
Choroba Alzheimera – nowe możliwości terapeutyczne oraz stosowane modele eksperymentalne
Adriana Kubis 1 , Maria Janusz 2Streszczenie
Choroby neurodegeneracyjne, a wśród nich choroba Alzheimera (AD), należą do najpoważniejszych schorzeń współczesnego społeczeństwa. Cechą procesu chorobowego jest stopniowe i nieodwracalne zaburzenie homeostazy organizmu. Stopniowemu zwyrodnieniu i obumieraniu neuronów towarzyszy powstawanie złogów amyloidowych, splotów nadmiernie ufosforylowanego białka tau, zaburzenie homeostazy jonów Ca2+. Występują również fagocytoza i aktywacja wydzielania czynników zapalnych, takich jak: cytokiny, reaktywne rodniki tlenowe czy tlenek azotu. Hiperaktywacja mikrogleju i astrocytów indukuje apoptozę komórek nerwowych oraz prowadzi do uszkodzenia bariery krew-mózg. Procesy te pociągają za sobą napływ komórek immunologicznie kompetentnych z obwodu i ich aktywny udział w lokalnym odczynie zapalnym. Zaburzenie mechanizmów kontroli procesu zapalnego prowadzi do zaburzeń w funkcjonowaniu organizmu i rozległej degeneracji struktury mózgu. Charakterystycznymi objawami AD jest stopniowy zanik pamięci powiązany z zaburzeniami procesów poznawczych, takich jak poprawne liczenie, orientacja przestrzenna czy upośledzenie mowy. Śledząc sekwencję zjawisk prowadzących do demencji o typie alzheimerowskim możliwości terapeutyczne można wiązać z modulacją aktywności sekretaz odpowiedzialnych za powstawanie amyloidogennych peptydów Aβ40–43, hamowaniem agregacji lub deagregacją peptydów Aβ, regulacją odpowiedzi zapalnej. Pomimo intensywnych badań oraz wysokich nakładów finansowych, nie udało się dotychczas opracować skutecznego i pozbawionego szkodliwych działań środka farmaceutycznego dla chorych na AD. Nowe perspektywy efektywnej terapii wiązane są z transplantacją komórek nerwowych oraz terapią genową. Prowadzone są również badania nad zastosowaniem białek uszkadzających strukturę β amyloidu. Ze względu na wieloprzyczynowy i wielokierunkowy charakter zmian leżących u podstaw AD bardziej efektywne wydaje się stosowanie w terapii kilku leków lub leku o wielokierunkowym działaniu. Doświadczenia prowadzone post mortem, a także neurochemiczne i anatomiczne badania mózgu stwarzają nowe możliwości zrozumienia mechanizmów leżących u podstaw chorób mózgu. Jednak ograniczony dostęp do mózgów jak i wyprowadzonych, pierwotnych linii komórkowych będących najodpowiedniejszym modelem badawczym, skłoniło badaczy do poszukiwania modeli pozwalających na monitorowanie zjawisk występujących w AD. Wydaje się, że spośród wielu modeli wprowadzonych do badań biomedycznych, zwierzęta transgeniczne spełniają wieloletnie dążenia do odzwierciedlenia mechanizmu procesu chorobowego. Pomimo dużej różnorodności stosowanych modeli badawczych często trudno jest znaleźć jednoznaczną odpowiedź nie tylko na pytanie jak przebiega proces neurodegeneracji, ale również w jaki sposób i jakimi czynnikami można go spowolnić, zatrzymać lub mu zapobiegać oraz w sposób jednoznaczny wykazać skuteczność proponowanej terapii. Jest to wciąż wyzwanie do prowadzenia interdyscyplinarnych badań.
Słowa kluczowe:choroba Alzheimera • amyloid β • białko tau • cytokiny • jony Ca2+ • stres oksydacyjny • modele eksperymentalne • kierunki terapeutyczne
Summary
Neurodegenerative disorders such as Alzheimer’s disease (AD) are the most common disease of modern society. The gradual and irreversible disturbances in homeostasis are characteristic features of the disease process. The cardinal features of AD include the formation of extracellular protein deposits in the brain that consist predominantly of aggregates of β amyloid protein (senile plaques), neurofibrilary tangles (hyperfosforylated tau protein) in the intracellular compartments, disturbances in calcium homeostasis, and degeneration/loss of synapses and neurons. An inflammatory process in the central nervous system is believed to play an important role in the pathway leading to neuronal cell death. The inflammatory response is mediated by activated microglia, resident immune cells of the central nervous system. Chronic activation of the microglia and astrocytes may cause damage of the brain-blood barrier and neuronal damage through the release of potentially cytotoxic molecules such as proinflammatory cytokines, reactive oxygen species, NO, and complement proteins. These alterations cause influx of immunocompetent cells from the periphery and their active participation in the local inflammatory reaction. Disturbances in the control mechanism of the inflammatory processes leads to perturbations in function and extensive brain degeneration. A characteristic symptom of AD dementia, is associated with dysfunctions of cognitive memory such as calculation, space orientation, and speech impairment. By tracking the sequence of events leading to the Alzheimer’s type of dementia, the therapeutic possibilities can be combined with modulation of secretase activation responsible for the formation of amyloidogenic forms Aβ40–43, inhibition of aggregation or β amyloid deaggregation, and regulation of the inflammatory response. Several strategies for drug intervention in both the treatment and prevention of AD has been pursued, but so far there is no fully effective cure without side effects. Transplantation of nerve cells and genetic therapy are looked upon as new perspectives. Research is being conducted on the application of proteins deforming β-sheet structures. Due to the pluricausal and multidirectional type of biological changes characteristic of AD, it seems likely that multidrug therapy or multidirectional medicine would be more efficient. Post-mortem experiments as well as neurochemical and anatomical brain studies helps to reveal new facts about the mechanisms underlying brain diseases. However limited access to fresh brain tissues or primary cell lines, which would be the best experimental models, compel researchers to look for other experimental models allowing investigation of disease occurrence. It seems that transgenic animals fulfill the requirements of relecting the disease process. In spite of the wide range of applied experimental models it is difficult to find clear answers to such questions as what are the exact stages of neurodegenerative process? How and what kind of factor could stop, slow down, or prevent this alterations? These questions are still open.
Key words:Alzheimer disease • β amyloid • tau protein • cytokines • Ca2+ ions • oxidative stress • experimental models • therapeutic prospects
Wykaz skrótów:
aa – reszty aminokwasowe; AD – choroba Alzheimera (Alzheimer’s disease); ADAS – skala oceny (objawów) choroby Alzheimera (Alzheimer’s disease assessment scale); ADAS-CGIC – skala oceny globalnej (objawów) choroby Alzheimera (Alzheimer’s disease assessment scale – clinical global impression of change); ADAS-cog – skala oceny zaburzeń funkcji poznawczych (Alzheimer’s disease assessment scale – cognitive subscale); AICD – wewnątrzkomórkowa domena białka APP (APP intracellular domain); ApoE – apolipoproteina E; APP – białko prekursorowe amyloidu (amyloid precursor protein); Aβ – amyloid β; Aβ40 – 40-aminokwasowy peptyd amyloidu β; Aβ42 – 42-aminokwasowy peptyd amyloidu β; Aβ43 – 43-aminokwasowy peptyd amyloidu β; BACE – enzym trawiący białko APP w pozycji β (beta-site APP cleaving enzyme); FRET – fluorescencyjny rezonans energetyczny (fluorescence resonance energy transfer); GDS – skala oceny rozpadu poznawczego (global detrioration scale); HTLV-1 – ludzki retrowirus T – limfocytotropowy (human T-cell lymphotropic virus); IADL – skala oceny złożonych czynności życia codziennego (instrumental activities of daily living); IFN-γ – interferon γ; Ig – immunoglobulina; IL – interleukina; iNOS – indukowalna syntaza tlenku azotu (inducible nitric oxide synthase); LPS – lipopolisacharyd z Escherichia coli, serotyp 055: B5; MAPK – kinaza białkowa aktywowana mitogenem (mitogen-activated protein kinase); MHC – główny układ zgodności tkankowej (major histocompatibility complex); MMSE – test oceniający podstawowe wymiary aktywności poznawczej (mini-mental state examination); NF–κB – jądrowy czynnik transkrypcyjny κB (neuronal factor κB); NFT – sploty neurowłókienkowe (neurofibryllary tangles); NK – komórki cytotoksyczne (natural killers); NKT – naturalne cytotoksyczne limfocyty T (natural killers T); NMR – jądrowy rezonans magnetyczny (nuclear magnetic resonance); NOS – syntaza tlenku azotu (nitric oxide synthase); O2– – jon ponadtlenowy; OH– – jon hydroksylowy; PRP – kompleks polipeptydowy bogaty w prolinę (proline-rich polypeptide complex); PS – presenilina (presenilin); ROS – wolne rodniki tlenowe (reactive oxygen species); SOD – dysmutaza ponadtlenkowa (superoxide dysmutase); SPECT – fotonowa tomografia emisyjna (single photon emission computed tomography); TNF-α – czynnik martwicy nowotworu α (tumor necrosis factor α); wit. D3 – 1,25-dwuhydroksywitamina D3
1. CHOROBA ALZHEIMERA JAKO PRZYKŁAD SCHORZENIA NEURODEGENERACYJNEGO
Choroby neurodegeneracyjne, a wśród nich najczęściej występujące choroby Alzheimera (AD) i Parkinsona, należą do najpoważniejszych schorzeń współczesnego społeczeństwa. AD występuje głównie u osób starszych, po 65 roku życia. Szacuje się, że na tę chorobę cierpi około 20 milionów osób, co jest odzwierciedleniem wydłużenia okresu życia – starzenia się społeczeństwa. Około 5% ludzi w wieku 65–74 lat, 20% 74–80 roku życia, a 33–50% w wieku 90 lat jest chorych na AD. Z prognoz demograficznych wynika, że w Polsce w 2011 roku będzie 237–285 tys. osób powyżej 65 roku ze zdiagnozowaną AD [161]. Cechą procesu chorobowego jest stopniowe i nieodwracalne zaburzenie homeostazy organizmu. Neurodegeneracji – stopniowemu zwyrodnieniu i obumieraniu neuronów towarzyszy powstawanie niefizjologicznych form białek zdolnych do agregacji i opornych na działanie enzymów proteolitycznych, oraz uszkodzenie szlaków przekazywania sygnałów. W organizmach chorych można zaobserwować różnorakie formy patologiczne: zbudowane z β-amyloidu (Aβ) blaszki starcze, sploty neurofibrylarne złożone z nadmiernie ufosforylowanego białka tau, ciałka Picka lub Lewiego i inne wtręty zawierające np. α-synukleinę. Dochodzi do nadmiernej indukcji astrocytów oraz komórek mikrogleju. Zachodzi również fagocytoza oraz uaktywnia się wydzielanie wielu czynników zapalnych, takich jak: cytokiny, reaktywne rodniki tlenowe czy tlenek azotu. Hiperaktywacja mikrogleju i astrocytów indukuje apoptozę komórek nerwowych oraz prowadzi do uszkodzenia bariery krew–mózg tak istotnej dla integralności oraz prawidłowego funkcjonowania układu nerwowego. Procesy te pociągają za sobą napływ komórek immunologicznie kompetentnych z obwodu i ich aktywny udział w lokalnym odczynie zapalnym. Zaburzenie mechanizmów kontroli procesu zapalnego prowadzi do rozległej degeneracji struktury mózgu oraz poważnych zaburzeń w funkcjonowaniu całego organizmu.Do czynników ryzyka wystąpienia AD zalicza się: zaawansowany wiek, mutacje genów presenilin, polimorfizm apolipoproteiny E (ApoE), zmiany aktywności enzymów, takich jak: α-antychymotrypsyna, butyrylocholinoesteraza K czy kompleks dehydrogenazy ketoglutarowej [27,38].
Procesy neurodegeneracyjne są trudne do zdefiniowania, gdyż ich objawy oraz obserwowane zmiany histologiczne i fizjologiczne są do siebie zbliżone, co skutecznie utrudnia postawienie jednoznacznej diagnozy, a co za tym idzie zastosowanie odpowiedniej terapii. Niezbędne jest założenie, iż proces neurodegeneracyjny to wieloprzyczynowe, ogólne zwyrodnienie mózgu, u którego podstaw leży agregacja zdegenerowanych białek [157].
Charakterystycznymi objawami AD jest stopniowy zanik pamięci powiązany z zaburzeniami procesów poznawczych, takich jak poprawne liczenie, orientacja przestrzenna czy upośledzenie mowy. Średnio i mocno zaawansowane stadium choroby cechuje pogłębiająca się dysfunkcja procesów poznawczych oraz funkcjonalnych, które prowadzą do niesamodzielności i całkowitej zależności od opiekuna. W trakcie rozwoju choroby niszczony jest układ odpornościowy, spada masa ciała, zwiększa się ryzyko infekcji płuc i gardła. Większą zachorowalność obserwuje się u kobiet niż u mężczyzn, co jest związane z prawdopodobnym zaangażowaniem estrogenów w rozwój AD [39,99].
Oprócz wieku i obciążeń rodzinnych jako czynniki ryzyka wymienia się:
• długotrwałe, wysokie ciśnienie,
• przebyte urazy głowy,
• duże stężenie homocysteiny.
Wyróżnia się dwa typy AD. We wczesnym stadium choroby pierwsze symptomy są zauważalne przed 60 rokiem życia, co stanowi 5–10% wszystkich przypadków i jest postacią postępującą znacznie szybciej niż postać starcza. Występuje w rodzinach, gdzie obserwuje się autosomalną, dziedziczną mutację dominującą. Dotychczas zidentyfikowano cztery geny związane z AD. Są to geny białka prekursorowego β-amyloidu (APP) na chromosomie 21, preseniliny 1 (PS1) na chromosomie 14, preseniliny 2 (PS2) na chromosomie 1 oraz ApoE (allel e4). Podejrzewa się również, iż w etiopatogenezie AD mogą mieć znaczenie geny: α2-makroglobuliny, składnika dehydrogenazy a-ketoglutaranu, wariantu K butyrylocholinoesterazy oraz niektóre geny mitochondrialne. W częściej występującej, późnej postaci AD rola genów jest bardziej pośrednia i mniej poznana. Mogą one być związane z podwyższonym prawdopodobieństwem tworzenia płytek starczych oraz splotów neurofibrylarnych [27,154].
Odkrycie mechanizmów leżących u podstaw procesów neurodegeneracyjnych to wyzwanie dla współczesnej nauki. Mimo ogromnego postępu wiedzy większość z nich nadal ma charakter hipotetyczny. Należy poszukiwać ulepszonych oraz nowoczesnych metod diagnozowania i leczenia coraz bardziej rozpowszechniających się chorób neurodegeneracyjnych.
Na podstawie badań mających na celu wyjaśnienie mechanizmów powstawania AD, prowadzonych w ostatnim dziesięcioleciu, można wysunąć dwie główne hipotezy: hipotezę kaskady β-amyloidu oraz hipotezę degeneracji cytoszkieletu. Hipoteza kaskady β-amyloidu zakłada, że proces neurodegeneracji to ciąg zdarzeń na poziomie biochemicznym pociągających za sobą nieprawidłowe przetwarzanie białka prekursora β-amyloidu (APP) [12].
W hipotezie kaskady Aβ wyróżnia się specyficzną sekwencję zdarzeń prowadzącą do degeneracji systemu nerwowego i demencji (ryc. 1) [134].

Ryc. 1. Schemat sekwencji zdarzeń prowadzących do degeneracji systemu nerwowego i demencji
Według hipotezy kaskady Ab pierwotnym wydarzeniem w etiopatogenezie AD jest odkładanie się złogów Aβ40–43 w postaci blaszek amyloidowych (starczych) w korze mózgowej chorych. Pozostałe zmiany neuropatologiczne: zwyrodnienie włókienkowe neuronów typu Alzheimera (NFT), zwyrodnienie synaps oraz zanik neuronów mają charakter wtórny i są następstwem gromadzenia się Aβ. Argumentem na występowanie zaburzeń w wytwarzaniu Aβ przed powstawaniem NFT jest to, iż mutacje genów dla APP, PS1 oraz PS2 zwiększają wydzielanie Aβ.
Hipoteza degeneracji cytoszkieletu jako proces wyjściowy zakłada zmiany cytoszkieletu neuronów prowadzące do degeneracji aksonów, zaburzeń przesyłania sygnałów itp. [57].
1.1. Mechanizm powstawania peptydów Aβ
Peptydy Aβ są produktami katalizowanej przez proteazy (α-, β- i γ-sekretazy), degradacji APP, będącego integralnym białkiem membranowym. Białko APP ma krótki czas połowicznego rozpadu i jest szybko metabolizowane. Niektóre izoformy APP (APP751 oraz APP770) powstające w wyniku alternatywnego składania genów (splicing), mają blisko N-końca domenę inhibitora Kunitza. Gen APP, o bardzo zmiennym transkrypcie, składa się z 18 eksonów o łącznej długości ponad 170 kDa. Region odpowiedzialny za sekwencję Aβ obejmuje część eksonu 16 i 17. Koduje 40–43 reszt aminokwasowych usytuowanych w transmembranowej i zewnątrzkomórkowej części APP. Białko prekursorowe Aβ składa się z dużej, zewnątrzkomórkowej domeny, hydrofobowej domeny transbłonowej oraz krótkiego, cytoplazmatycznego C-końcowego fragmentu sprzężonego z białkami cytoszkieletu. Pierwsze 17 reszt aminokwasowych sekwencji Aβ (reszty 597–613 APP) znajduje się na zewnątrz błony, natomiast pozostałe 26 reszt (614–639 APP) należy do części transbłonowej APP. N-końcowy odcinek sekwencji Aβ to bogata w cysteinę domena wiążąca heparynę (VHHQK). APP jest białkiem konserwatywnym. Sekwencja aminokwasowa jest niemal identyczna u różnych gatunków poczynając od Drosophila melanogaster a na człowieku kończąc. Sekwencja Aβ człowieka różni się jedynie trzema resztami aminokwasowymi od występującego u gryzoni: piąta reszta – R u ludzi, G u gryzoni; dziesiąta – Y u ludzi, F u gryzoni oraz trzynasta reszta – H u ludzi, R u gryzoni [52,66,154].
Budowa i funkcjonowanie sekretaz nie są jeszcze w pełni zbadane. Sekretaza-α tnąc APP pomiędzy 687 a 688 aminokwasem w zewnątrzkomórkowej części domeny, prowadzi do powstania pozostającego w błonie, 83 aa, C-końcowego fragmentu CT83, oraz rozpuszczalnego, neuroprotekcyjnego sAPPα. α-sekretaza ma cechy zakotwiczonej w błonie metaloproteazy a jej funkcja może być kontrolowana poprzez kinazę będącą pod wpływem receptora muskarynowego. Funkcję α-sekretazy mogą pełnić białka z rodziny dezintegryn, metaloproteaz lub enzym aktywujący czynnik martwicy nowotworów α. Powstające pod wpływem α-sekretazy białko CT83 jest substratem γ-sekretazy, która poprzez proteolizę w środku domeny transbłonowej powoduje powstanie 3 kDa białka p3 oraz wewnątrzkomórkowej domeny amyloidu CT57–59 (AICD) i/lub CT31 (ryc. 2). Reakcje te nie prowadzą do powstania patogennych postaci Aβ [57,77,79].

Ryc. 2. Mechanizm fizjologicznego przetwarzania APP [174]
W procesie prowadzącym do powstania amyloidogennych, patologicznych postaci peptydów, β-sekretaza przecina APP pomiędzy N-końcowymi resztami aminokwasowymi 671–672 prowadząc do powstania rozpuszczalnego fragmentu sAPPβ oraz CT99. Następnie, pod wpływem γ-sekretazy z CT99 powstają 4 kDa Aβ40/Aβ42/Aβ43 oraz wewnątrzkomórkowa domena APP – AICD (ryc. 3, tab. 1).
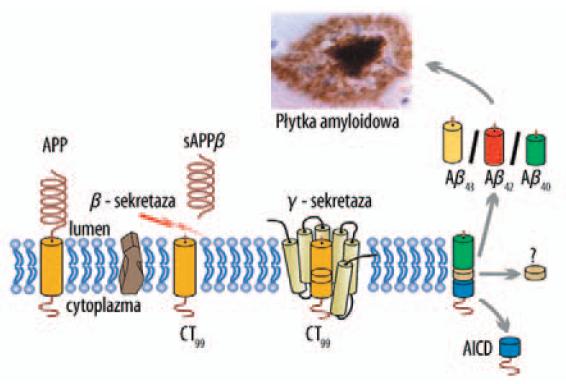
Ryc. 3. Mechanizm przetwarzania APP prowadzący do powstania patogennych postaci Aβ40–43 [174]
Tabela 1. Białka powstające w procesie proteolizy APP oraz ich znane i przypuszczalne działanie [115,134]

β-sekretaza jest białkiem należącym do rodziny proteaz aspartylowych. Składa się z pojedynczej domeny transmembranowej, sekwencji sygnałowej oraz peptydowego regionu na N-końcu. W domenie zewnętrznej występują dwie reszty kwasu asparaginowego Asp93 oraz Asp289, które są niezbędne do prawidłowej aktywności enzymu. Wewnątrzkomórkowa β-sekretaza występuje przede wszystkim w aparacie Golgiego i endosomach. Gen kodujący jest umiejscowiony na chromosomie 11. Mutacje tego genu nie wpływają na częstotliwość występowania AD. γ-sekretaza ma cechy proteazy aspartylowej i odznacza się bardzo niewielką swoistością w stosunku do sekwencji substratu. Mutacje APP obejmujące reszty aminokwasowe w pobliżu miejsca działania enzymu nie wpływały hamująco na wytwarzanie Aβ. Uważa się, iż bardzo ważną rolę w kompleksie białkowym γ-sekretazy odgrywają dwa wysoce homologiczne białka zwane presenilinami (PS1 i PS2) [80,142]. Są one białkami transbłonowymi mającymi 7–9 odcinków przenikających przez błonę komórki. Występują głównie w komórkach nerwowych, gdzie mogą pełnić funkcje receptorów błonowych lub kanałów wapniowych. Ponadto powodują one wzrost podatności na apoptozę oraz regulują homeostazę wapniową w retikulum endoplazmatycznym. Najsilniejszą ekspresję presenilin obserwuje się w komórkach hipokampa oraz komórkach Purkiniego. Sugeruje się, że PS1 może spełniać funkcję kofaktora γ-sekretazy lub też działać jako γ-sekretaza uczestnicząc w ten sposób w kontroli przetwarzania APP. Mutacje w genach presenilin są związane z powstawaniem 40, 42 i 43 aminokwasowych postaci peptydów amyloidu β [48,57,133,174].
Wydaje się, że działanie γ-sekretazy nie jest swoiste i może mieć ona wpływ również na inne białka czy enzymy biorące udział w różnego rodzaju szlakach metabolicznych. Większość Aβ powstającego pod wpływem działania gsekretazy ma 40 reszt aminokwasowych, ale powstaje też niewielka pula dłuższych 42 lub 43 aminokwasowych, i bardziej hydrofobowych wariantów – Aβ42 oraz Aβ43. Odznaczają się one większą skłonnością do oligomeryzacji i agregacji w porównaniu z Aβ40. Mimo że Aβ42 oraz Aβ43 powstają w mniejszej ilości w porównaniu z jego krótszą formą, to właśnie te białka są podstawowymi formami budującymi płytkę amyloidową [86,108,116].
Neurotoksyczność patologicznych postaci Aβ42–43 jest związana m.in. z powodowaniem zaburzeń w homeostazie Ca2+, interakcją z lipidami błony komórkowej oraz aktywacją swoistych receptorów. Akumulacja Aβ42–43, zarówno na zewnątrz, jak i wewnątrz komórki, inicjuje kaskadę wydarzeń prowadzących do neurodegeneracji, takich jak: uszkodzenie neuronów, aktywacja astrocytów i komórek mikrogleju (odczyn zapalny), zakłócenie równowagi jonowej w neuronach, uszkodzenia oksydacyjne, zmiany w aktywności kinaz/fosfataz, tworzenie się NFT, dysfunkcja neuronów i w końcu śmierć komórki. Aβ42 występujący w niewielkich (1–10 nM) ilościach w płynie mózgowordzeniowym ludzi zdrowych oraz w płynie znad hodowli izolowanych komórek nerwowych pełni prawdopodobnie rolę w fizjologicznych funkcjach ośrodkowego układu nerwowego. Działanie Aβ42 nie jest jeszcze w pełni poznane [135,138].
W mózgu obecne są również proteazy inne niż sekretazy, które prawdopodobnie uczestniczą w powstawaniu i regulacji peptydów Aβ. Mogą to być katepsyna D i E, żelatynaza A lub B, trypsyno- lub chymotrypsynopodobne endopeptydazy, aminopeptydazy, neprylizyna, kompleks α2-makroglobuliny i proteaz serynowych lub enzym degradujący insulinę. Mimo prowadzonych w szerokim zakresie badań nad mechanizmem powstawania fizjologicznych i patologicznych postaci Aβ nie poznano dotychczas wszystkich enzymów biorących w nim udział [132,163].
1.2. Rola wewnątrzneuronalnych splotów w procesach neurodegeneracyjnych
Białko tau jest fosfoproteiną wiążącą mikrotubule. Prawdopodobnie odpowiada za ich łączenie się i stabilizację. W komórkach nerwowych białko to występuje w aksonach. W stanach patofizjologicznych transportowane jest do ciała komórki oraz dendrytów [92].
W dojrzałym mózgu człowieka występuje sześć izoform białka tau, które są produktem alternatywnego cięcia mRNA pojedynczego genu leżącego na chromosomie 17q21. Izoformy różnią się występowaniem lub brakiem 29 lub 58 aminokwasowych fragmentów w pobliżu N-końca oraz przy C-końcu 31 aminokwasowej, konserwatywnej sekwencji kodowanej przez ekson 10. Ta ostatnia zawiera w sobie fragment odpowiedzialny za wiązanie mikrotubuli. Jej wpływ na gromadzenie i wiązanie się mikrotubuli jest wprost proporcjonalny do ilości powtórzeń tej sekwencji w białku tau [88]. Procentowa zawartość poszczególnych izoform białka tau różni się pomiędzy gatunkami. Ze wzrostem fosforylacji obniża się zdolność białka tau do wiązania się ze szkieletem komórkowym. Za obniżenie poziomu fosforylacji odpowiedzialne są fosfatazy. Zahamowanie hiperfosforylacji białka tau polega na hamowaniu aktywności enzymów, takich jak: kinazy syntezy glikogenu 3 (GSK3), cyklinozależnej kinazy 5 (CDK5), kinazy białkowej aktywowanej mitogenem (MAPK), kinazy białkowej aktywowanej stresem (SAP) lub aktywacji głównej fosfatazy występującej w mózgu – białkowej fosfatazy 2A (PP2A) [70,140,144,167].
W chorobie Alzheimera pierwszym mechanizmem związanym z białkiem tau jest jego hiperfosforylacja a następnie ubikwitynacja. Hiperfosforylowane izoformy białka tau tworzą sploty wewnątrzneuronalne (NFT) zbudowane ze sparowanych włókien helikalnych (SPF). Po śmierci komórek NFT będące wcześniej w cytosolu, są obecne w przestrzeni międzykomórkowej w postaci tzw. „duchów splotów”, które są zbudowane przede wszystkim z ubikwitynylowanych, powtarzających się fragmentów sekwencji białka tau. Mimo iż mechanizm powstawania NFT nie jest do końca poznany, sugeruje się, iż zwiększona ilość reszt fosforanowych wzmaga ich odłączanie się od mikrotubuli, przez co zwiększa się ilość niezwiązanej fosfoproteiny. Ta postać jest o wiele odporniejsza na degradację i bardziej skłonna do agregacji niż postać związana z mikrotubulami. Obecność NFT, złogów amyloidowych oraz ubytek neuronów poważnie upośledzają funkcje pamięciowe i powodują rozwój otępienia [65,76,126].
Mechanizm powstawania NFT jest w pewnego rodzaju opozycji do procesu formowania się płytek amyloidowych. Ilość oraz miejsca tworzenia się złogów Aβ są bardzo różnorodne i zależne od cech osobniczych. W hipokampie oraz w korze mózgowej złogi te powstają później niż NFT, co nie jest zgodne z hipotezą kaskady amyloidu. Hipotetyczny model łączący Aβ z ufosforylowanym białkiem tau zakłada, iż Aβ42 powoduje zaburzenia transportu międzysynaptycznego, osłabienie sygnału, który wstrzymywał aktywację kinaz i/lub inhibicję fosfataz, co prowadzi do nadmiernej fosforylacji określonych reszt białka tau. Zmiany konformacyjne tej fosfoproteiny prowadzą do powstawania zaburzeń w transporcie aksonalnym, a w końcu do ograniczenia funkcjonowania i żywotności neuronu i jego apoptozy [51,89,113].
1.3. Rola stresu oksydacyjnego oraz metabolizmu wolnych rodników tlenowych (ROS)
Stres oksydacyjny opisywany jest jako stan, w którym komórkowa obrona antyoksydacyjna jest nieskuteczna wobec nadmiernego wydzielania oksydantów. Może on występować lokalnie. Przezwyciężenie obrony antyoksydacyjnej w określonych organach lub tkankach nie wpływa na aktywność antyoksydacyjną całego organizmu. Mechanizmy obrony antyoksydacyjnej są swoiste dla poszczególnych ROS. Podstawowe konsekwencje stresu oksydacyjnego to: fragmentacja lipidów lub ich zmiany strukturalne, denaturacja białek, zaburzenia w mechanizmach replikacji DNA oraz deformacje organelli komórkowych, a co za tym idzie całych komórek. Stres wywołany przez wolne rodniki tlenowe prowadzi nie tylko do wystąpienia reakcji zapalnej, ale również uruchamia zależną od NF-κB transkrypcję genów dla różnych czynników zapalnych [9,22].
ROS są niezbędnymi mediatorami wielu ważnych reakcji biologicznych. Mogą również niszczyć makrocząsteczki wywołując stres oksydacyjny. Odgrywają one istotną rolę m.in. jako wtórne przekaźniki uczestniczące w kontroli ekspresji genów. Ponadto stanowią podstawowe narzędzie walki komórek fagocytarnych, które wykorzystują oksydanty do zabijania organizmów patogennych. Wolne rodniki mają na zewnętrznej orbicie elektronowej pojedynczy niesparowany elektron. Należą do nich atom wodoru, cząsteczka tlenu zawierająca dwa niesparowane elektrony w tym samym spinie, ale na dwóch osobnych orbitalach, NO•, jon ponadtlenowy O2–, rodnik hydroksylowy OH–, rodnik peroksynitrowy ONOO–. Reakcje, w których biorą udział ROS są katalizowane głównie przez jony metali, takie jak Fe2+, Cu2+. Wyjątkowa wrażliwość mózgu na uszkodzenia spowodowane ROS wynika z obecności wielonienasyconych kwasów tłuszczowych oraz dużego stężenia jonów Fe3+, Cu2+, Zn2+ występujących w substancji czarnej i prążkowiu. Oba te czynniki zwiększają także wrażliwość błon komórkowych mózgu na proces peroksydacji lipidów. Ze względu na dużą zależność funkcjonowania mózgu od metabolizmu tlenowego poziom aktywności oddechowej lokalnych mitochondriów jest znacznie wyższy niż w innych tkankach. Zwiększa to ryzyko niekontrolowanego wypływu ROS z tych organelli. Aktywacja mikrogleju prowadzi do zwiększonego wytwarzania i uwalniania cytokin oraz NO, łatwo wchodzącego w reakcje z rodnikami tlenowymi tworząc ONOO–, którego sprotonowana postać rozpada się do NO2 i OH–. Obrona przed rodnikami polega na przekształceniu ich za pomocą dysmutazy do H2O2 rozkładanego przez katalazę. Większość ROS jest niestabilna i ma krótki okres półtrwania oraz dąży do przekazania niesparowanego elektronu do najbliższej cząsteczki lub połączenia z innym wolnym elektronem. Reakcje katalizowane m.in. przez wolne rodniki są wyjątkowo ważne w procesach zachodzących w komórkach eukariotycznych. NO pełni rolę neuroprzekaźnika w ośrodkowym układzie nerwowym, gdzie bierze udział w procesach rozwoju mózgu, uczenia się i pamięci oraz w regulacji czynności ruchowych i pobierania pokarmu. W obwodowym układzie nerwowym współuczestniczy w rozkurczach mięśni gładkich przewodu pokarmowego, narządów miednicy oraz dróg oddechowych. Tlenek azotu działa jako międzykomórkowy przekaźnik regulujący napięcie naczyń krwionośnych, aktywujący płytki krwi oraz uczestniczący w kontroli odpowiedzi immunologicznej. Ochronne działanie NO przejawia się hamowaniem degranulacji komórek tucznych, blokowaniem wytwarzania toksycznego jonu ponadtlenowego oraz hamowaniem adhezji płytek krwi i neutrofilów do śródbłonka naczyń krwionośnych. Tlenek azotu reguluje procesy wydzielania cytokin oraz prowadzi komórkę na drogę apoptozy lub jej zapobiega. Hamuje ekspresję selektyn typu P i E, przez co ogranicza wędrówkę limfocytów Th1 do miejsca zapalenia [26,35,46,73,150].
Badania prowadzone na myszach transgenicznych dowiodły, iż w mózgu zwierząt z amyloidozą i patologiczną postacią preseniliny 1 obserwuje się czterokrotnie większą aktywność indukcyjnej syntazy tlenku azotu (iNOS). Toksyczne działanie Aβ przejawia się zdolnością do aktywowania czynnika transkrypcyjnego NF-κB nadzorującego ekspresję iNOS i wytwarzanie NO. Stężenie iNOS zwiększa się w mózgu pod wpływem procesu zapalnego będącego następstwem powstawania złogów amyloidowych. Hiperaktywacja tego enzymu prowadzi do uwalniania dużych ilości NO zdolnych do nieodwracalnego zaburzenia funkcji komórek. Co więcej, pobudzone astrocyty i mikroglej otaczające złogi amyloidowe wytwarzają IL-1β, TNF-α oraz ROS, które aktywują ekspresję iNOS [35,40,90,96].
Ochronnemu działaniu NO często towarzyszy wydzielanie małej ilości ROS, które powodują:
• inaktywację białek zawierających grupy tiolowe,
• utlenianie produktów glikacji białek,
• zahamowanie glikolizy przez inaktywację dehydrogenazy aldehydu 3-fosfoglicerynowego,
• peroksydację lipidów prowadzącą do zaburzenia potencjału jonowego błony komórkowej,
• uszkodzenie DNA [68,82].
Między niskocząsteczkowymi związkami o właściwościach antyoksydacyjnych, enzymami rozkładającymi je oraz tempem wytwarzania ROS istnieje równowaga dynamiczna, która wpływa na metabolizm i aktywność wolnych rodników. Nawet drobne zaburzenia tej równowagi prowadzą do wyjątkowo niebezpiecznego i trudnego do opanowania stresu oksydacyjnego. Do enzymów czuwających nad prawidłowym poziomem ROS należą:
• peroksydaza glutationowa,
• reduktaza glutationu,
• dysmutazy ponadtlenkowe umiejscowione w cytoplazmie (FeSOD oraz Cu/Zn SOD),
• dysmutazy ponadtlenkowe macierzy mitochondrialnej (MnSOD),
• katalaza,
• mieloperoksydaza,
• kompleks oksydaz NADPH [131].
1.4. Kaskada cytokin
Cytokiny to grupa ponad 100 niskocząsteczkowych glikoprotein o właściwościach i funkcjach zbliżonych do hormonów, a działających już w stężeniach pikomolowych. Ich wielokierunkowe oddziaływanie objawia się poprzez regulację wzrostu, procesów proliferacji, pobudzania aktywności komórek układu odpornościowego oraz hemopoetycznego. Wpływają one nie tylko na leukocyty, ale również na inne komórki organizmu, stymulując powstawanie gorączki, regulując morfogenezę komórek i tkanek, czy też działając cytotoksycznie. W zależności od stanu organizmu cytokiny aktywują odpowiedź komórkową lub humoralną oraz budują wyjątkowo efektywny, ale także bardzo złożony i wrażliwy system powiązań pomiędzy komórkami układu immunologicznego oraz między samymi cytokinami, tzw. sieć cytokin. Poszczególne komórki mają na swojej powierzchni receptory określonych cytokin, dlatego też ich działanie jest wysoce selektywne. Nie każda komórka jest w stanie reagować na daną cytokinę, nie każda także może tę cytokinę wytwarzać. Funkcjonowanie sieci cytokin uwarunkowane jest wieloma czynnikami, m.in.: lokalnym stężeniem oraz rodzajem wydzielanych cytokin („koktajl cytokin”), rodzajem komórek, ich współdziałaniem i obecnymi na nich receptorami cytokin [1,125].
W AD hiperaktywacja komórek mikrogleju wywołuje stan zapalny, w którym dochodzi do uszkodzenia bariery krew–mózg. Aktywowane komórki mikrogleju wytwarzając duże ilości cytokin wpływają stymulująco na astrocyty. Pobudzają je do wytwarzania białek prozapalnych, ROS oraz NO. Sprzyja to powstawaniu nierozpuszczalnej postaci białka Aβ wykazującego neurotoksyczne właściwości.
IL-10 ma właściwości immunosupresyjne. Uczestniczy w wygaszaniu odpowiedzi immunologicznej typu komórkowego i wytwarzaniu immunotolerancji. Reguluje wytwarzanie ROS i NO przez makrofagi oraz ekspresję cząsteczek MHC klasy II na monocytach. Jest odpowiedzialna za utrzymanie dynamicznej homeostazy między aktywnością proi antyzapalną komórek układu immunologicznego przez co chroni przed ich hiper- lub hipoaktywacją [152]. W patogenezie AD podkreślona jest rola IL-10, jako cytokiny o właściwościach antyzapalnych mogącej niwelować działanie cytokin prozapalnych. Obniżona sekrecja IL-10 obserwowana była u pacjentów z AD, jednak nie było wyraźnej zależności od stopnia zaawansowania choroby [8,149].
IL-6 jest wytwarzana w mózgu przez astrocyty i mikroglej, a głównymi czynnikami indukującymi jej wytwarzanie są IL-1 oraz TNF. Przypuszczalnie stymuluje ona różnicowanie się neuronów. W warunkach fizjologicznych zabezpiecza mózg przed toksycznymi czynnikami zapalnymi oraz niedoborami tlenu. Ze względu na neutropowe właściwości IL-6, jej nadmierne wytwarzanie prawdopodobnie wpływa aktywująco na ekspresję genów APP. Przez hamowanie podziału komórkowego oraz immunostymulujące właściwości może wpływać na rozwój niektórych nowotworów. Przeciwciała przeciwko tej cytokinie stosuje się z dużym powodzeniem w leczeniu reumatoidalnego zapalenia stawów [49,153].
Wzrost sekrecji IL-1β przez aktywowane komórki gleju wiązany jest z postępem AD [85,102,123]. Poziom wydzielanej IL-1β obniżony był u pacjentów leczonych donepezilem [50]. Według ostatnio wysuniętej hipotezy, na podstawie wyników uzyskanych w modelowych badaniach na myszach transgenicznych z nadekspresją IL-1β, przypuszcza się, że ta cytokina może stymulować usuwanie złogów Aβ w procesie fagocytozy [81].
Istotną rolę w patogenezie AD odgrywają IFN-γ oraz TNF-α. Efekt działania IFN-γ jest dwukierunkowy. Z jednej strony poprzez aktywację wydalania kwasu glutaminowego, wraz z NO powstającym w stresie oksydacyjnym, powoduje apoptozę neuronów. Z drugiej strony, tak jak i TNF-α hamuje wytwarzanie APP, przez co chroni komórki nerwowe przed toksycznymi produktami jego przetwarzania [19,40,120,138]. TNF-α wytwarzany jest przez komórki mikrogleju w odpowiedzi na czynnik zapalny. Na istotną rolę TNF-α w patogenezie AD wskazuje 25-krotny wzrost poziomu tej cytokiny w płynie mózgowo-rdzeniowym chorych wzrastający w zależności od nasilenia procesu chorobowego. TNF-α synergistycznie z IFN-γ stymuluje rozpoczęcie i prawidłowy przebieg reakcji układu odpornościowego. Białka rodziny TNF regulują wydzielanie NO oraz innych cytokin, takich jak np.: IFN-g przez limfocyty, IL-1, IL-6, GM-CSF, G-CSF, M-CSF, EGF, NGF czy IFN-β przez makrofagi. Myszy pozbawione receptorów TNF były o wiele bardziej podatne na uszkodzenie neuronów powstających np. wskutek niedokrwienia mózgu, co sugeruje neuroprotekcyjne właściwości tych białek. Tak jak w przypadku wcześniej opisanych cytokin próbuje się zastosować w terapii TNF w połączeniu z innymi cytokinami lub z chemioterapią [34,96,110,130].
Ze względu na złożony i nie do końca poznany mechanizm oddziaływań w obrębie sieci cytokin, zastosowanie ich w terapii jest wyzwaniem współczesnej nauki. Frapujące właściwości immunomodulatorowe cytokin stwarzają wiele możliwości zastosowań w lecznictwie, jednak delikatna równowaga istniejąca pomiędzy indukowaniem i hamowaniem wydzielania poszczególnych cytokin stwarza utrudnienia w ich stosowaniu. Plejotropowość cytokin powoduje, że mogą one mieć działania niepożądane. Mimo tych ograniczeń IFN stosowany jest w leczeniu wirusowego zapalenia wątroby typu C. Próbuje się również wykorzystać fragmenty IL-1 oraz białka homologiczne do IL-1 w regulacji procesów zapalnych, IL-10 w hamowaniu reakcji odrzucenia przeszczepu czy G-CSF w terapii niedoborów neutrofilów [13,125,130].
1.5. Funkcja i metabolizm jonów Ca2+
Oprócz złogów amyloidowych i NFT tworzonych przez hiperfosforylowane białko tau istotną rolę w patologii AD odgrywają jony Ca2+. Stężenie tych jonów w komórce jest regulowane przez pompy wapniowe, potasowe lub przez przyłączanie do białek. Istotną rolę odgrywają również kanały jonowe regulowane przez receptory neuroprzekaźników, np. kwasu glutaminowego. Uszkodzenie receptorów prowadzi do zaburzenia funkcji mitochondriów, aktywacji proteaz oraz lipaz. Organellami komórkowymi przechowującymi jony są mitochondria, retikulum endoplazmatyczne, ciałka wydalnicze, lizosomy oraz jądro. Ca2+ jest wtórnym przekaźnikiem sygnału działającym zarówno na błony komórkowe jak i wewnątrz komórki. Tak zwane fale wapniowe, czyli lokalnie przemieszczające się skupiska podwyższonego stężenia Ca2+ w cytosolu są charakterystyczne dla stanu pobudzenia. Jest on niezbędny w szybkich odpowiedziach komórki, takich jak skurcz i rozkurcz [24].
Transport Ca2+ odgrywa ważną rolę m.in. w apoptozie oraz w uszkodzeniach mózgu charakteryzujących się niedokrwieniem prowadzącym do śmierci komórek nerwowych. W szczurzych tkankach hipokampu śmierć komórek może być zahamowana przez wzrost wewnątrzkomórkowego gradientu Ca2+. Aβ powoduje aktywację transportu jonów do komórki. Wykazano, iż syntetyczne fragmenty aminokwasowe Aβ1–38 oraz Aβ25–35 zwiększają spoczynkowe stężenie Ca2+ oraz podwyższają wapniozależną odpowiedź na depolaryzację błony komórkowej. Aβ może pełnić funkcję kanału jonowego kontrolującego wpływ jonów do komórki. W błonach komórek nerwowych chorych na AD zaobserwowano zwiększoną aktywność wymiennika sodowo-wapniowego, a w badaniach post mortem podwyższoną mobilizację białka aktywowanego Ca2+ – kalpainy. Z podwyższonym stężeniem Ca2+ wiąże się obniżenie stabilności mRNA dla iNOS [24,98].
U chorych na AD, na poziomie komórkowym zachodzą zmiany morfologiczne typowe dla procesów apoptotycznych: uwypuklenie dwuwarstwowej błony lipidowej z wytworzeniem charakterystycznych pęcherzyków, powstawanie ciałek apoptotycznych, kondensacja chromatyny czy reorganizacja cytoszkieletu komórkowego. Reorganizacja struktury wewnątrzkomórkowej prowadzi do degeneracji neuronów. Proces ten jest obserwowany podczas stresu termicznego lub oksydacyjnego. W AD obserwuje się również powstawanie ciałek Hirano, czyli parakrystalicznych eozynofilnych filamentów o grubości 7 nm zbudowanych z aktyny, tubuliny, winkuliny, tropomiozyny, białka MAP, α-aktyniny, kofiliny. Aβ indukuje polimeryzację aktyny poprzez białka szoku cieplnego (HSP – heat shock protein) i aktywację kaskady kinaz. W wyniku tego procesu na powierzchni komórek powstają włókna stresowe, których tworzenie hamowane jest przez inhibitory MAPK oraz γ-sekretazy [23,111].
1.6.Objawy i diagnostyka AD
We wczesnym stadium AD objawy choroby są trudno zauważalne, często mylone z typowym zachowaniem związanym z wiekiem. W ostatnim stadium choroby rozwijającej się przez kilka lat pacjent traci kontakt z otoczeniem, nie radzi sobie z wykonywaniem podstawowych czynności, wymaga stałej pomocy ze strony opiekunów [27,161].
Przyczyny demencji można ustalić za pomocą jądrowego rezonansu magnetycznego (NMR), fotonowej tomografii emisyjnej (SPECT) lub pozytronowej tomografii emisyjnej (PET). Pomimo intensywnych badań i rozwoju nauk medycznych AD jest przeważnie zbyt późno diagnozowana. Wczesne rozpoznanie utrzymałoby lub podwyższyło jakość życia chorych, obniżyłoby koszty społeczne oraz zmniejszyłoby ciężar opieki ze strony rodziny [91,93].
Nie ma swoistego biochemicznego czy genetycznego markera wczesnego rozpoznania AD. W diagnostyce schorzeń neurologicznych najczęściej stosowane są oznaczenia w płynie mózgowo-rdzeniowym, co oprócz utrudnionego pobierania materiału, u chorych na AD jest dodatkowo ograniczone obniżonym wydzielaniem tego płynu. Kontroluje się poziom oraz stosunek peptydów Aβ42: Aβ43 i Aβ40: Aβ42 – wskaźników liczby blaszek starczych; ilość białka tau oraz stopień jego fosforylacji jako markera splotów neurofibrylarnych. Istotne dla diagnostyki AD jest to, że postać hiperfosforylowana występująca w mózgu ma zmniejszoną rozpuszczalność w stosunku do postaci fizjologicznej. W przeciwieństwie do zdrowych, starzejących się ludzi w płynie rdzeniowo-mózgowym u chorych na AD wzrasta poziom ApoE wraz z postępem choroby, co może dawać podstawy do traktowania go jako markera monitorującego nasilanie się stanu chorobowego. Zastosowanie zestawu kilku markerów oraz metod neuroobrazowania może znacznie podwyższyć czułość diagnostyki AD [47,93,135].
W osoczu krwi chorych na AD zidentyfikowano 18 białek sygnałowych, których analiza biologiczna wskazuje, iż są one charakterystyczne dla procesów, takich jak: dysregulacja hematopoezy oraz odpowiedzi immunologicznej, apoptozy oraz innych zmian presymptomatycznych AD. Obecność tych białek we krwi pozwala na stwierdzenie z prawie 90% dokładnością, że pacjent cierpi na łagodne zaburzenia pamięci, które w ciągu najbliższych 2–6 lat doprowadzą do AD [122].
Chorobie Alzheimera towarzyszy rozwój procesu zapalnego w mózgu. Można go obserwować oznaczając aktywność α-1-antychymotrypsyny oraz stężenia IL-1β i IL-6. Następny marker – peroksydację lipidów indukowaną przez wolne rodniki aktywujące proces szoku tlenowego, można monitorować przez badanie w moczu poziomu 8-izoprostaglandyny F2a powstającej z kwasu arachidonowego. Jej podwyższony poziom koreluje z ilością pochodnej tromboksanu B2 i jest charakterystyczny dla otępienia typu AD. Ostatnie doniesienia sugerują, iż podwyższony poziom kalikreiny 6 (proteazy serynowej) w płynie mózgowo-rdzeniowym, osoczu, krwi oraz ekstraktach z tkanki mózgowej jest charakterystyczny dla pacjentów z AD. Oznaczany w mózgu stosunek wolnej 8-hydroksy-2-dezoksyguanozyny (produkt ataku rodników tlenowych na DNA) do związanej z DNA był ponad stukrotnie wyższy u chorych na AD niż w grupie kontrolnej. Szczególnie w przypadkach występowania demencji w rodzinie warto zastosować środki prewencji, takiej jak np.: dieta niskotłuszczowa, aktywność psychofizyczna, przyjmowanie niesteroidowych leków przeciwzapalnych czy statyn [117,145,146].
2. STOSOWANE I POTENCJALNE ŚRODKI TERAPEUTYCZNE
Obserwując sekwencję zjawisk prowadzących do demencji o typie alzheimerowskim (ryc. 4), możliwości terapeutyczne można wiązać z modulacją aktywności sekretaz odpowiedzialnych za powstawanie amyloidogennych peptydów Aβ40–43, hamowaniem agregacji/deagregacji peptydów Aβ, regulacją odpowiedzi zapalnej (wydzielanie cytokin, ROS, ochrona neuronów przed uszkodzeniem powodowanym Aβ). Pomimo intensywnych badań oraz wysokich nakładów finansowych, nie udało się dotychczas opracować skutecznego i pozbawionego działań niepożądanych środka farmaceutycznego dla chorych na AD. Podstawowymi elementami w terapii AD są:
• spowolnienie postępu choroby,
• kontrola nad zachowaniami, dezorientacją oraz stanami niepokoju,
• modyfikacja środowiska domowego,
• wsparcie członków rodziny oraz opiekunów.

Ryc. 4. Hipotetyczna sekwencja patologicznych zmian zachodzących w AD
Wspomagające w procesie leczenia są: zmiana stylu życia (słuchanie muzyki relaksującej oraz stosowanie technik relaksacyjnych, regularne spacery etc.), stosowanie leków wspomagających oraz suplementów (antyutleniacze), takich jak witamina E czy wyciąg z Ginkgo biloba [94,100].
Jedyną dotychczas zaaprobowaną i szeroko stosowaną metodą leczenia jest stosowanie inhibitorów cholinoesterazy. Esteraza acetylocholinowa to enzym, który rozkłada acetylocholinę. Zahamowanie jego działania, powoduje w mózgu wtórne podwyższenie stężenia acetylocholiny będącej najważniejszym przekaźnikiem biorącym udział w procesach pamięciowych. Leki o aktywności inhibitorów acetylocholinesterazy powodują podwyższenie stężenia acetylocholiny w synapsach, co polepsza transport cholinergiczny oraz łagodzi zaburzenia procesów poznawczych. Ich efektywność jest niestety uzależniona od dawki. Analizując dane uzyskane w badaniach klinicznych wykazano korzystny, aczkolwiek łagodny wpływ na parametry poznawcze oceniane w teście ADAS w trakcie 6–12 miesięcy stosowania terapii. Objawowe leczenie polegające na podwyższeniu poziomu acetylocholiny w mózgu znane jest już od końca lat osiemdziesiątych dwudziestego wieku. Początkowo stosowanymi lekami były: takryna i welakryna, które pojawiły się na rynku amerykańskim. Wieloośrodkowe badania kliniczne wykazały, iż leki te, zwłaszcza w dużych dawkach, skutecznie łagodzą zaburzenia poznawcze. Niestety, przestały być używane ze względu na ich bardzo dużą hepatotoksyczność [16,54]. W 1997 r. wprowadzono lek nowej generacji – donepezil (Aricept) będący wybiórczym inhibitorem esterazy acetylocholinowej pozbawionym już właściwości uszkadzania miąższu wątroby. Hamuje on jedynie esterazę acetylocholinową nie wpływając na butyrylocholinesterazę. Randomizowane, wieloośrodkowe badania porównywane z grupą, której podawano placebo dowiodły skuteczności leczenia donepezilem w okresie 52 tygodni, w toku są badania wieloletnie. Dzięki jego działaniu część chorych wydłuża okres względnej samodzielności o dwa lata, co, zważywszy na zaawansowany wiek osób z AD, jest okresem znaczącym. Objawy niepożądane to krótkotrwałe nudności, czasami zwolnienie czynności serca, zawroty głowy, koszmarne sny [54,136,163]. Kolejnym inhibitorem, który pojawił się na rynku i był zarejestrowany z tym samym wskazaniem jest rivastigmina (Exelon). Lek przyjmuje się dwa razy dziennie. Powolne podwyższanie dawek zmniejsza objawy niepożądane, jakimi są: zaburzenia przewodu pokarmowego, bóle i zawroty głowy oraz nadmierne pobudzenie. Wyniki badań wskazują, że poza poprawą możliwości poznawczych lek wpływa także korzystnie na zaburzenia zachowania i wykonywanie codziennych czynności. Od donepezilu różni się tym, że hamuje także działanie butyrylocholinoesterazy, co również podwyższa poziom acetylocholiny w mózgu. Podawanie rivastigminy i donepezilu jest ograniczone występującymi działaniami niepożądanymi oraz zmniejszającym się wraz z czasem stosowania korzystnym oddziaływaniem [44,143]. Ostatnio pojawił się na rynku trzeci, wybiórczy inhibitor esterazy acetycholinowej pochodzenia roślinnego – galantamina (Reminyl), która dodatkowo moduluje allosterycznie działanie receptora nikotynowego. Wpływa ona również pozytywnie na możliwości poznawcze chorego. Istnieje coraz więcej danych wskazujących, że hamowanie esterazy acetylocholinowej hamuje także amyloidogenezę [25]. Obecnie inhibitory esterazy acetylocholinowej przyjmuje bardzo wielu chorych. Tym samym wiedza na temat ich przydatności po długim okresie stosowania, czy ich wpływu na przebieg choroby będzie coraz pełniejsza. Objawy niepożądane bywają niekiedy tak uciążliwe, że zmuszają do rezygnacji z kuracji. Często też chory nie podejmuje leczenia z powodów ekonomicznych. Przydatność tych leków w przedłużaniu fazy łagodnej i średnio zaawansowanej, pozwalającej na samodzielną lub prawie samodzielną egzystencję jest niewątpliwym postępem w leczeniu AD. Korzystne działania w szczególności na pamięć poznawczą zaobserwowano u pacjentów chorych na AD podczas 2-letnich otwartych badań klinicznych nad galantaminą [32]. Prowadzone są badania nad poznaniem mechanizmów działania inhibitorów esterazy acetylocholiny i ich wpływu na:
• przemianę APP w Aβ,
• cytotoksyczność indukowaną Aβ,
• promowanie działania α-sekretazy,
• hamowanie aktywności β-sekretazy.
Obecnie dąży się do zahamowania agregacji peptydów amyloidowych lub fosforylacji białka tau oraz inaktywacji GSK-3β (kinazy 3β syntezy glikogenu) [59].
Rola cholesterolu w centralnym systemie nerwowym jest często niedoceniana lub pomijana. Poziom cholesterolu w mózgu jest sześć razy wyższy niż w wątrobie i we krwi łącznie. Ważnym białkiem biorącym udział w metabolizmie oraz dystrybucji cholesterolu w neuronach jest Apo E. Promuje ona powstawanie i agregację płytek amyloidowych oraz oddziałuje ze składnikami cytoszkieletu indukując powstawanie splotów neurowłókienkowych. ApoE kodowana jest przez gen na chromosomie 19 i występuje w postaci trzech izoform: ε2, ε3 i ε4 różniących się resztami cysteiny i argininy w pozycjach 112 i 158. Cholesterol powstający in vivo w neuronach i mikrogleju z udziałem ApoE warunkuje plastyczność synaps. Podwyższony poziom cholesterolu może być czynnikiem ryzyka AD i może być obniżany przez zastosowanie statyn hamujących działanie reduktazy 3-hydroksy-3-metyloglutarylo-koenzymu A. Są one stosowane w hamowaniu powstawania miażdżycy oraz w chorobach układu krwionośnego. Regulują proces fosforylacji białka tau oraz metabolizm ROS i dlatego mogą być alternatywną metodą terapii różnego rodzaju schorzeń neurodegeneracyjnych, w tym AD. Zastosowanie statyn może zmniejszać wydzielanie Aβ poprzez regulację równowagi pomiędzy aktywnością α- oraz β- i γ-sekretazy, a także może wpływać na hamowanie procesów związanych ze stanem zapalnym. Wydaje się również, iż mogą one usuwać Aβ z mózgu poprzez oddziaływanie z białkami pokrewnymi receptorowi LDL (LRP) w ścianach naczyń krwionośnych. Badania epidemiologiczne wykazały zmniejszenie częstotliwości występowania AD u chorych poddanych terapii statynowej. Nie miała ona jednak wpływu na tempo rozwoju demencji [29,114,121,124,164,171].
Badania nad możliwością zastosowania immunoterapii w AD są konsekwencją pozytywnych wyników prac nad stworzeniem transgenicznych modelów zwierzęcych. Wykazały one, iż immunizacja chroni przed powstawaniem neuropatologicznych objawów typowych dla AD, a w niektórych przypadkach udało się spowodować rozpuszczenie płytek amyloidowych. Zastosowanie aktywnej jak i biernej immunizacji obniża częstość występowania zmian patologicznych charakterystycznych dla AD oraz przywraca funkcje poznawcze u transgenicznych myszy, mimo że sam mechanizm działania przeciwciał nie jest jeszcze do końca poznany. Na przykład, u myszy szczepu CRND8 immunizowanych Aβ, w 50% zahamowane było powstawanie złogów amyloidowych w porównaniu z grupą kontrolną. Najbardziej efektywne w badaniach in vivo i ex vivo okazało się przeciwciało IgG2a, wykazujące duże powinowactwo do receptora Fc mikrogleju. Istnieje kilka hipotez na poparcie korzystnych efektów immunoterapii w chorobie AD. Jedna z nich zakłada wiązanie się przeciwciał do fibryli Ab i przez to prezentowanie ich fagocytującemu mikroglejowi. Inne wyjaśnienie poparte jest obserwacją, iż długoterminowe podawanie przeciwciał monoklonalnych m266 powoduje wzrost Aβ w osoczu z jednoczesnym zahamowaniem powstawania nowego Aβ. Przeciwciała m266 powodując degradację Aβ we krwi, zaburzają homeostazę między ośrodkowym układem nerwowym a osoczem, co prowadzi do wydalania Aβ z mózgu. Kolejna hipoteza zakłada, że przeciwciała hamują fibrylogenezę i chronią komórki przed cytotoksycznym działaniem Aβ poprzez oddziaływanie z resztami 4–10 Aβ42. Po pomyślnych wynikach immunoterapii uzyskanych na myszach, królikach, świnkach morskich oraz małpach rozpoczęto II fazę badań klinicznych na ochotnikach. Zastosowano szczepionkę AN-1792 przeciwko AD koniugowaną z QS-21– środkiem wspomagającym odpowiedź immunologiczną na antygen zastosowany w szczepionce. Jednak stosowanie immunizacji Aβ u ludzi zostało zawieszone przede wszystkim z powodu indukcji procesów zapalnych w ośrodkowym układzie nerwowym. U 6% ochotników biorących udział w badaniach rozwinęły się objawy zapalenia mózgu. U myszy domiejscowe podawanie szczepionki powodowało rozpuszczanie złogów amyloidowych. Zauważono jednak powstawanie krwotoków, co stanowiło uboczny proces związany z immunizacją. Pomimo to, stosowanie przeciwciał monoklonalnych skierowanych przeciwko różnym fragmentom Aβ może zaowocować uzyskaniem lepszych szczepionek. Przeciwciała anty-Aβ mogą hamować cięcie APP przez sekretazy via blokowanie konformacyjne [55, 56,69,71,97,118,137,158,166,168].
Wydaje się, iż zastosowanie związków pochodzenia organicznego, takich jak: kwas rozmarynowy – naturalny roślinny polifenol o aktywności antyoksydacyjnej, przeciwzapalnej oraz antymikrobiotycznej; ekstrakty z alg czy z ziół mogą chronić komórki przed odpowiedzią zapalną i procesami cytotoksycznymi wywołanymi Aβ [4,28,75]. Ekstrakt Egb 761 z Ginkgo biloba poprawiał pamięć oraz zachowanie u chorych z zaburzeniami poznawczymi związanymi z wiekiem oraz chorych na demencję. Ma on właściwości antyoksydacyjne oraz prawdopodobnie przeciwzapalne. Hamuje powstawanie płytek amyloidowych. Wieloośrodkowe badania na ludziach nie dostarczają jednak jednoznacznych informacji na temat pozytywnego wpływu wyciągu z Ginkgo biloba u chorych na AD oraz inne schorzenia neurologiczne. Prowadzone są badania nad zastosowaniem ekstraktu z Zingiber officinale i Alpina galanga. Zaobserwowano bowiem hamujący wpływ tych ekstraktów na indukowaną LPS, cytokinami lub amyloidem b ekspresję genów czynników prozapalnych, takich jak TNF-α, IL-1β, COX-2, MIP-α, MCP-1 oraz IP-10 na komórkach THP-1 [67,106].
Ze względu na zaangażowanie kaskady antyoksydacyjnej w ochronie organizmu przed AD wydaje się, iż zastosowanie środków przeciwzapalnych, takich jak: leki antyhistaminowe, niesteroidowe leki przeciwzapalne (NSAIDs), indometacyna, naproksen czy rofecoksib mogą korzystnie wpływać na hamowanie rozwoju choroby [165]. Badania prowadzone na pacjentach przyjmujących leki przeciwzapalne wykazały niższy stopień zachorowalności na AD w porównaniu do ludzi niestosujących tych środków. Jak wiadomo w przypadku AD, u podstaw procesów patologicznych leży stres oksydacyjny. Wykazano, iż przed śmiercią neuronów w mózgu, jak i niszczącym wpływem wolnych rodników chronią substancje o właściwościach antyoksydacyjnych, takie jak flawonoidy, witamina E czy C [6,18]. Stosowane i potencjalne środki terapeutyczne oraz ich mechanizm działania przedstawiono w tab. 2. Dowiedziono, iż terapia estrogenowa zmniejsza ryzyko wystąpienia AD u kobiet przez zahamowanie powstawania złogów Aβ [99]. Memantyna (Namenda) – antagonista receptora N-metylo- D-asparaginianu (NMDA) korzystnie wpływa na pamięć, funkcjonowanie, zachowanie i samopoczucie chorych na AD. Terapia ta jest stosowana w łagodnych i średniozaawansowanych stadiach choroby. Może być również stosowana wraz z inhibitorami cholinesteraz [37,112]. Nowe perspektywy efektywnej terapii wiążą się z transplantacją komórek nerwowych oraz metodami terapii genowej [21,43,109].
Tabela 2. Stosowane środki terapeutyczne i mechanizm ich działania [42]

Prowadzone są również badania nad zastosowaniem białek niszczących strukturę β amyloidu istotną dla tworzeniu się fibryli amyloidowych (ryc. 5).
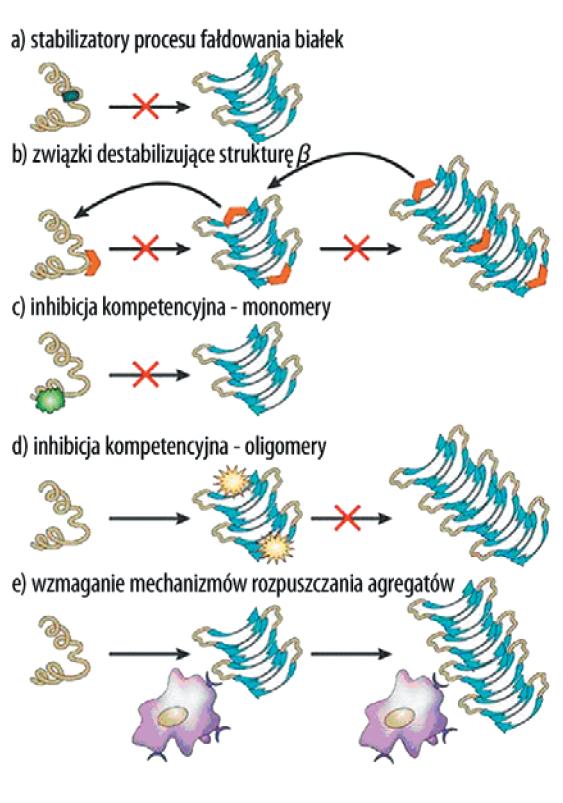
Ryc. 5. Strategie terapeutyczne mające na celu regulację zmian konformacyjnych białek lub zahamowanie ich agregacji [103,147]; a) stabilizacja prawidłowego procesu fałdowania się białek, b) hamowanie lub odwracanie patogennego procesu fałdowania się białek poprzez związki swoiście destabilizujące konformację β-kartki, c) inhibicja kompetycyjna podczas procesu oligomeryzacji białek przez związki wiążące się do form monomerycznych, d) inhibicja kompetycyjna procesu agregacji białek poprzez wiązanie się do zagregowanych struktur β-kartki – blokowanie dalszego przyłączania monomerow, e) zastosowanie związkow aktywujących mechanizmy prowadzące do rozpuszczenia powstałych agregatow lub zaburzających ich stabilność
Jako matrycę do projektowania tego typu związków zastosowano środkowy, hydrofobowy region pomiędzy 17-20 resztami aminokwasów (LPFD) w N-końcowej domenie Aβ. Ponadto dodano reszty proliny, które są powszechnie znanymi blokerami struktury β oraz aminokwasy zawierające ładunek w celu zwiększenia rozpuszczalności projektowanego białka. W ten sposób otrzymany iAβ5 hamował in vitro powstawanie agregatów Aβ40 i Aβ42, a także rozpad fibryli już zagregowanych oraz zmniejszał cytotoksyczność Aβ42 w stosunku do ludzkich komórek neuroblastoma i w mózgach szczurów szczepu Fischer-334. Jego główną wadą jest jednak bardzo krótki czas rozkładu in vivo [2,148].
Barierami w zastosowaniu peptydów w terapii chorób są: niski poziom przenikalności przez barierę krew–mózg, wrażliwość na działanie enzymów proteolitycznych, słaba rozpuszczalność oraz cytotoksyczność. W początkowej fazie procesu tworzenia się złogów amyloidowych szczególnie ważną rolę pełnią sekretazy. Kontrola lub – chociaż częściowe zahamowanie działania β- i/lub γ-sekretazy – pociągnęłoby za sobą monitorowanie powstawania różnych postaci Aβ [141,172]. W przypadku β-sekretazy poszukuje się silnie działających inhibitorów, które mogłyby dopasowywać się do miejsca aktywacji tej proteazy serynowej i jednocześnie bez przeszkód przechodziłyby przez barierę krew–mózg. Poszukuje się również związków blokujących aktywację cytotoksycznych procesów indukowanych oddziaływaniem Aβ z powierzchnią komórki z użyciem blokerów kanałów jonowych. Kolejną atrakcyjną metodą leczenia może być przeciwdziałanie lub hamowanie powstawania i odkładania się patologicznych postaci Aβ i kontrola polimeryzacji białka tau (ryc. 6) [7,31].
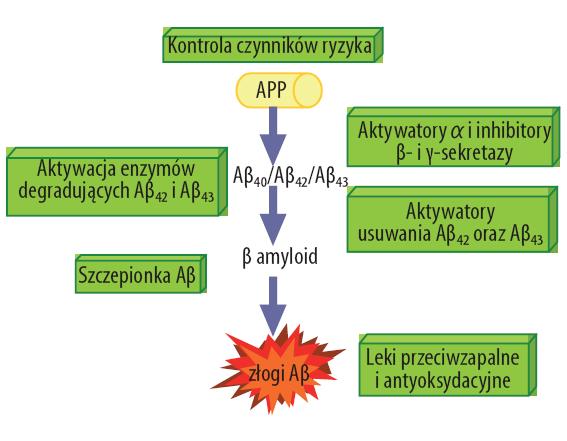
Ryc. 6. Potencjalne miejsca interwencji terapeutycznej i profilaktyki AD [58]
Celem współczesnych badań nad opracowaniem skutecznego środka terapeutycznego w AD jest również – zaangażowane w proces neurodegeneracji – białko tau. Poddaje się analizie miejsca wiążące oraz przeprowadza się badania nad obniżeniem ekspresji genów tego białka. Pozytywną rolę w hamowaniu reakcji prozapalnych mogą też odegrać jony metali. Na podstawie badań in vitro oraz na modelach zwierzęcych wykazano, że lit spełnia kryteria leku przeciw demencji [87,113].
Jako nowe podejście terapeutyczne proponuje się zastosowanie komórek macierzystych izolowanych z układu krwiotwórczego. Wiąże się z tym nadzieje, że przywrócą one prawidłowe funkcjonowanie ośrodkowego układu nerwowego. Podstawową, bardzo przydatną cechą tych komórek jest samoodnawialność oraz to, iż występują w płodowym i dojrzałym systemie nerwowym. Hodowle tych komórek mogą być prowadzone w warunkach laboratoryjnych i mogą się różnicować do komórek glejowych lub neuronów. Wykazano, iż po uszkodzeniu mózgu, neurony mogą powstawać z endogennych komórek macierzystych [91].
Poszukiwania skutecznego leku zmierzają także w kierunku zahamowania, czy raczej regulacji funkcjonowania zbyt silnie pobudzonych komórek mikroglejowych. Takie działanie może przejawiać naczyniowoczynne białko jelitowe (VIP), które jest neuropeptydem o potencjalnych właściwościach przeciwzapalnych. Jego korzystne działanie wykazano w chorobach szoku endotoksycznego czy reumatoidalnym zapaleniu stawów. W badaniach in vitro i in vivo wykazano, iż peptyd ten chronił komórki nerwowe przed apoptozą przez hamowanie wytwarzania czynników zapalnych, takich jak: TNF-α, IL-1β czy NO [3].
Prowadzenie badań nad środkami mogącymi mieć znaczenie terapeutyczne w AD musi zakładać wieloprzyczynowy oraz wielokierunkowy charakter zmian leżących u podstaw tego schorzenia. Dlatego też pojedynczy, swoisty związek może nie być efektywny w ochronie oraz w terapii AD. Wydaje się, iż zastosowanie terapii składającej się z kilku leków będzie bardziej efektywne. Jest ona jednak bardziej skomplikowana i przez to trudniejsza do opracowania. Najkorzystniejsze efekty dałoby zastosowanie leku przejawiającego wielokierunkowe działanie będącego jednocześnie nieszkodliwym dla organizmu.
Kryteria takiego leku spełnia kompleks polipeptydowy bogaty w prolinę izolowany z siary owczej – Colostrinina™. Skuteczność Colostrininy bez działań niepożądanych wykazana została w dwuletnich badaniach klinicznych i potwierdzona w wieloośrodkowych badaniach prowadzonych w 6 polskich ośrodkach klinicznych. Obejmowały one 15-tygodniową podwójną ślepą próbę kontrolowaną przez placebo, a następnie 15-tygodniowy okres podawania badanego preparatu. U ponad 100 pacjentów objętych 30-tygodniowym okresem badań, nie stwierdzono działań niepożądanych. Skuteczność terapii przejawiającą się jako poprawa lub stabilizacja wykazano w testach IADL i ADAScog. Określają one płaszczyznę funkcjonalną (aktywność w życiu codziennym) – poznawczą (pamięć, mowa) oraz zdolność rozumowania [15,83,84].
3. MODELE BADAWCZE
Prowadzenie badań nad etiopatogenezą AD i monitorowanie działania potencjalnych leków jest utrudnione względami etycznymi. Przeprowadzanie eksperymentów na chorych, otrzymywanie i przechowywanie skrawków tkanki mózgowej oraz izolowanie komórek z mózgu post mortem jest trudne do wykonania. Stwarza to wyzwanie do tworzenia modeli badawczych umożliwiających śledzenie możliwie jak największej liczby elementów procesu chorobowego.
3.1. Linie komórkowe
Z organów i tkanek, w tym również z mózgu ludzkiego, można wyprowadzić linie ciągłe lub linie pierwotne o określonym czasie życia (ryc. 7).
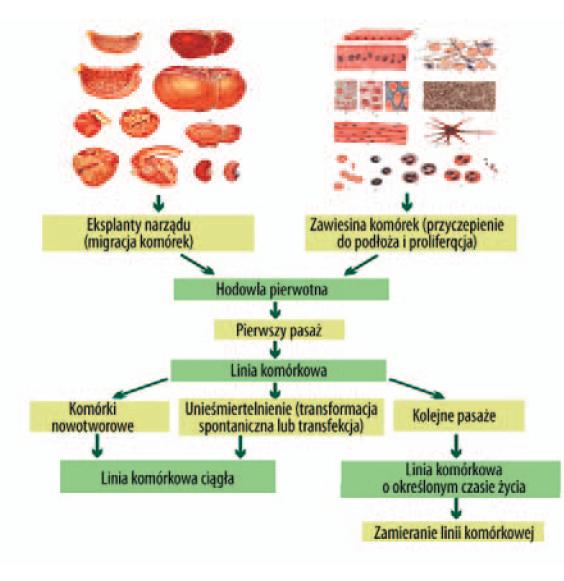
Ryc. 7. Wyprowadzanie linii pierwotnych z tkanek lub narządow [155]
Linie ciągłe to hodowle komórek izolowanych z guzów nowotworowych, zawierające nieograniczoną zdolność proliferacji. Uzyskane z tego typu komórek linie można wielokrotnie pasażować. Linie ciągłe można również wyodrębnić z mieszaniny komórek pochodzenia mózgowego np. dzięki ich zdolności do wyłapywania acetylowanej lipoproteiny o niewielkiej gęstości. Czas życia takich komórek w hodowli zależy od wieku organizmu, z którego pobrano komórki, rodzaju tkanki, jak i od dobowego, sezonowego lub endokrynnego cyklu narządu. Starzenie się komórek w hodowli jest odzwierciedleniem starzenia się populacji komórkowych in vivo. Dłuższą żywotnością odznaczają się linie komórkowe wyprowadzone z komórek zarodków lub z komórek macierzystych (stem cells). Znacznie krótszą żywotność wykazują linie komórkowe pochodzące od osobnika dorosłego. Linie komórkowe, które mają ograniczony czas życia, nazywa się finite cell lines. Rosną one przez konkretną liczbę pokoleń, osiągając zwykle 20–80 podwojeń populacji, potem komórek nie da się już dłużej pasażować i linia komórkowa zamiera [78,151].
3.1.1. Pierwotne linie komórek mikrogleju
W badaniach nad etiopatogenezą AD modelem badawczym o istotnym znaczeniu są pierwotne linie komórek mikrogleju. Są to komórki glejowe pochodzenia szpikowego występujące w ośrodkowym układzie nerwowym, które po pobudzeniu nabierają zdolności do fagocytozy. Komórki te są tkankowo swoiste i biorą udział w odpowiedzi immunologicznej. Istnieje wiele hipotez wyjaśniających pochodzenie komórek mikrogleju. Część badaczy uważa, że są to komórki wywodzące się z linii monocytarnej pochodzenia mezodermalnego. Dowodem na to były doświadczenia ze znakowaniem monocytów krwi obwodowej niemowląt, które następnie identyfikowano w tkance nerwowej mózgu. W cytoplazmie tych komórek występowały również fragmenty lizozymu oraz enzymów (esteraza czy peroksydaza). Na powierzchni komórek mikrogleju obecne są markery F4/80, Mac-1, ED1, lektyny (GSA I-B4) charakterystyczne dla monocytów i makrofagów oraz receptor fragmentu Fc. Alternatywnym źródłem komórek mikrogleju wydaje się neuroektoderma. Wywodzą się z niej również glioblasty będące prekursorami astrocytów i oligodendrocytów. Komórki prekursorowe oligodendrocytów i astrocytów, oraz mikroglej mają bardzo zbliżone cechy histochemiczne. Zakłada się, że mikroglej wykształcił się jako odrębna linia z komórek macierzystych neurogleju. Istnieje również pogląd przyjmujący heterogenne pochodzenia komórek mikrogleju. Część jest pochodzenia mezodermalnego, a część neuroektodermalnego. W mózgu mikroglej przyjmuje różnorakie postaci i funkcje uwarunkowane stanem fizjologicznym tkanki [45,129].
Mikroglej może stanowić nawet 20% całkowitej populacji komórek nieneuronalnych w mózgu. Jego podstawowymi zadaniami są:
• fagocytoza komórek apoptotycznych,
• reakcja na obecność patogennego antygenu,
• kontrolowanie środowiska tkanki.
Komórki mikrogleju ulegają transformacji z postaci spoczynkowej do aktywowanej. Polega ona m.in. na powiększeniu ciała komórki, obkurczeniu rozgałęzień, ekspresji białek adhezyjnych, reorganizacji cytoszkieletu oraz ekspresji MHC klasy I i II [129,151].
Po naruszeniu integralności tkanki mózgowej lub zaburzeniu homeostazy jonowej uruchamiane jest wiele mechanizmów mających za zadanie ochronę przed patogenami, a także naprawę uszkodzeń. Następuje aktywacja ekspresji genów odpowiedzialnych za stymulację astrocytów i mikrogleju oraz indukcja wydzielania receptorów błonowych, które wzmagają odpowiedź. Należą do nich:
• receptory rozpoznające cząsteczki związane z patogenem,
• receptory komplementu (np. CR1, CR3, CR4),
• receptory cytokin (np. TNFRI, TNFRII, IL-1RI, IL-12R),
• receptory chemokin (np. CCR2, CCR3, CXCR4, CX3CR1),
• receptory ułatwiające interakcje z układem odpornościowym, np. z limfocytami T czy immunoglobulinami (np. FcRI – RIII) [105].
Stymulowany mikroglej zaczyna wytwarzać:
• cytokiny (IL-1, TNF-α, IL-6, IL-12, IL-15, IL-18),
• chemokiny (fraktalkina, IP-10, MIP-1α, MIP-1β, MCP-1, RANTES, MDC),
• związki cytotoksyczne (iNOS, ROS),
• prostanoidy (PGD2, PGE2, tromboksan B2) [10].
Wynikiem interakcji z uszkodzonymi komórkami nerwowymi, oddziaływań z pobudzonymi astrocytami oraz autostymulacji komórek mikrogleju jest podwyższona proliferacja, migracja oraz fagocytoza. Następuje ekspresja CD40 oraz MHC klasy I i II. Mikroglej transformuje do komórki prezentującej antygen (APC). Ze względu na rolę i zaangażowanie mikrogleju w ochronie tkanki nerwowej mózgu wydaje się, iż komórki te mogą być głównym celem terapii chorób o podłożu zapalnym w tym AD [53,127].
3.1.2. Linie komórkowe – HL-60, THP-1, PC12
Ograniczony dostęp do mózgów, jak i wyprowadzonych, pierwotnych linii komórkowych skłonił badaczy do poszukiwania linii komórkowych pozwalających na obserwowanie zjawisk występujących w AD.
HL-60 (komórki ludzkiej białaczki promielocytarnej) zostały wyprowadzone z komórek pochodzących od pacjenta chorego na ostrą białaczkę promielocytarną. Jest to linia dzieląca się, nieadherentna z czasem podwojenia 36–48 godzin. Proliferacja zachodzi z udziałem receptorów insuliny i transferytyny, które są ekspresjonowane na powierzchni komórek. Spontaniczne różnicowanie się do dojrzałych granulocytów może być wywoływane poprzez DMSO lub kwas retinowy. Inne czynniki, takie jak: 1,25 dwuhydroksywitamina D3, TPA lub GM-CSF mogą indukować różnicowanie się komórek odpowiednio do: monocytów, makrofago- lub eozynofilopodobnych komórek. Na komórkach HL-60 można badać zmiany zachodzące podczas różnicowania się komórek szpiku [33]. Genom HL-60 zawiera amplifikowany protoonkogen c-myc. Stężenie mRNA c-myc jest duże w niezróżnicowanych komórkach i raptownie spada już w początkowych etapach procesu różnicowania komórek [17]. Komórki HL-60 są stosowane jako modele do badań nad mechanizmem migracji monocytów/makrofagów przez barierę krew–mózg u chorych na AD. Ponieważ, tak jak komórki PC12, wytwarzają sekrecyjną postać APP stosuje się je do badań nadmiernego wytwarzania lub nieprawidłowego katabolizmu APP. Zidentyfikowano w nich izoformę natywnego, 110–135 kDa kompleksu białkowego APPβ występującego we wszystkich strukturach dojrzałego mózgu [17,33,41].
THP-1 – ludzka linia komórek białaczki – wyprowadzona z krwi chłopca chorego na ostrą białaczkę monocytarną. Komórki te zawierają receptory Fc oraz C3b, lecz nie mają immunoglobulin powierzchniowych oraz cytoplazmatycznych. Monocytarny charakter tej linii został określony na podstawie obecności aktywnej naftylo-butyryloesterazy hamowanej przez NaF, wytwarzania lizozymu, aktywności fagocytarnej oraz zdolności aktywacji odpowiedzi limfocytów T na ConA. Komórki THP-1 nie mają jądrowego antygenu związanego z wirusem Epsteina-Barr [159]. Linia ta została użyta jako model komórek mikrogleju w badaniach nad hamowaniem hiperaktywności tych komórek przez Zingiber officinale i Alpinia galanga oraz inhibicją ekspresji genów czynników prozapalnych [64]. Używając komórek THP-1 przeprowadzono również badania nad zależnością między Aβ, metaloproteinazą 2 (MMP-2) oraz TGF-1β – immunosupresyjną cytokiną powiązaną z akumulacją Aβ w modelach mysich oraz biorącą udział w procesie niszczenia złogów amyloidowych przez aktywowane komórki mikrogleju. Stosując komórki THP-1 badano również korelację pomiędzy TNF-α, cAMP, białkiem p38 oraz zbudowanym ze 105 reszt aminokwasowych, C-końcowym fragmentem APP – białkiem prawdopodobnie biorącym udział w patologicznych zmianach występujących w AD [30]. Zaobserwowano, że fibrylarne postaci Aβ aktywują kaskadę sygnałową zależną od kinazy tyrozynowej w komórkach THP-1 oraz w komórkach mysiego mikrogleju. Pociąga to za sobą zwiększone wydzielanie czynników neurotoksycznych, wytwarzanie cytokin prozapalnych oraz ROS [34]. Prowadzono również badania nad możliwością regulacji wydzielania przez komórki THP-1 aktywowane Aβ, takich czynników jak: IL-1α, IL-1β, TNF-α, IL-6, MCP-1 [156].
Komórki PC12 to linia wyprowadzona z guza chromochłonnego nadnercza szczura. Podział oraz indukcja różnicowania tej linii komórkowej hamowane są przez NGF, co pozwala na używanie komórek PC12 jako modelowej linii różnicowania się neuronów oraz w badaniach mechanizmów działania NGF. Pod jego wpływem komórki PC12 zmieniają się fenotypowo z chromochłonnych na komórki podobne do neuronów układu współczulnego. PC12 traktowane Aβ25–35 wykazywały podwyższony poziom ROS oraz białka p53 [11]. W komórkach PC12 traktowanych chlorokininą (związek niszczący błony komórkowe, endosomowe i lizosomowe), na powierzchni komórek oraz endosomów akumulował się gangliozyd GM1 biorący udział w aktywacji agregacji Aβ [173].
3.2. Zwierzęta transgeniczne
Wydaje się, że spośród wielu modeli wprowadzonych do badań biomedycznych, zwierzęta transgeniczne spełniają wymagania, aby do badania różnych schorzeń nękających człowieka uzyskać modele wykazujące taki sam mechanizm procesu chorobowego. Udane próby przenoszenia materiału genetycznego między różnymi komórkami zachęciły do podjęcia badań nad opracowaniem metody pozwalającej na otrzymanie osobnika, który we wszystkich komórkach posiadałby obcy materiał genetyczny. Dotąd opracowując i stosując różne nowe metody wyprowadzono wiele różnych linii myszy lub szczurów transgenicznych. Zwierzęta jednego gatunku cechujące się jednorodnością genetyczną i homozygotycznością zostały nazwane szczepem wsobnym. Podszczepy to zwierzęta należące do szczepu wsobnego, ale różniące się od niego na poziomie genetycznym. Mutacje genetyczne są wywoływane przez czynniki przypadkowe lub w ramach planowanych działań. Dwie linie szczepu wsobnego różniące się jednym zmutowanym genem określane są jako pary koizogeniczne. Przez jednoczesne zastosowanie genetycznych reakcji krzyżowych oraz selekcji wyprowadza się szczepy kongeniczne różniące się krótkim odcinkiem jednego chromosomu. Szczepami kongenicznymi opornymi CR (congenic resistant) nazwane zostały dwie linie zwierząt transgenicznych zawierające mutacje na chromosomie mającym locus zgodności tkankowej. Zwierzęta z takich dwóch szczepów odrzucają wzajemnie swoje przeszczepy tkankowe. Serię szczepów wsobnych rekombinacyjnych (recombinant inbred strains – RIS) stanowi zbiór szczepów wyhodowanych z losowo skojarzonych par zwierząt. Pary te są wybrane z drugiego pokolenia mieszańców z krzyżówki między dwoma znanymi szczepami wsobnymi charakteryzującymi się odpowiednio brakiem lub ekspresją badanej cechy. Każdy nowy szczep RIS jest homozygotyczny w stosunku do jednego z dwóch alleli wniesionych przez przodków we wszystkich loci, w których przodkowie się różnili. W ten sposób w wyniku losowej rekombinacji niesprzężonych loci każdy szczep z danej serii RIS ma własną szczególną kombinację materiału genetycznego pochodzącego od obu szczepów rodzicielskich. Szczepy kongeniczne rekombinacyjne (recombinant congenic strains – RCS) zostały pomyślane jako narzędzia do badania uwarunkowanych genetycznie cech ilościowych kontrolowanych przez kilka współdziałających (addytywnych) genów. Do takich cech zalicza się m.in. oporność i podatność na nowotwory [61,95].
Istnieje wiele linii myszy transgenicznych, które charakteryzują się występowaniem złogów Aβ oraz płytek starczych (tab. 3).
Tabela 3. Wybrane mysie linie modelowe – APP/Aβ [95]

W mózgach myszy PDAPP (APPV717F mice) występuje postać rozpuszczalna Aβ, a także uformowane złogi amyloidowe. Dochodzi do utraty i zaburzenia sygnału w synapsach oraz zwiększenia liczby astrocytów i mikrogleju otaczającego złogi. Myszy mają zaburzony proces rozpoznawania obiektów, pamięć przestrzenną oraz procesy uczenia się. Pomimo wykrywania hiperfosforylacji białka tau w tym szczepie nie obserwuje się NFT. Linia ta jest szeroko rozpowszechniona i używana do badań nad szczepionkami przeciwko Aβ.
W kolejnym modelu – Tg2576 wraz z wiekiem zwiększa się poziom Aβ40 oraz Aβ42, co prowadzi do powstania dużych złogów Aβ w korze mózgowej, hipokampie i móżdżku. Myszy tej linii charakteryzują się neofobią – pogłębiającym się strachem przez nieznanymi przedmiotami, zaburzeniami pamięci oraz przedwczesną śmiercią. Zwiększona ekspresja SOD1, hemoksygenazy 1 oraz duże ilości 4HNE wskazują na występowanie stresu oksydacyjnego. U myszy tych nie obserwuje się zaawansowanych zmian w białku tau oraz zmian zapalnych. Nie mają one jednak NFT, więc nie są kompletnym modelem AD. Linie PDAPP oraz Tg2576 stanowią podstawę w projektowaniu różnego rodzaju mutantów cechujących się izoformami białek APP lub Aβ, lub nadekspresją związanych z nimi genów.
Szczep APP23 charakteryzuję się prawie 7-krotną nadekspresją APP, obecnością rozpuszczalnych płytek w korze nowej i hipokampie, astrocytozą oraz mikroglejozą. Żadna z dotychczas wyprowadzonych linii będących modelami mutacji APP lub Aβ nie charakteryzuje się obumieraniem neuronów typowym dla AD.
Mutacje na chromosomie 17q21.1, mimo że nie mają bezpośredniego powiązania z powstawaniem AD biorą udział w rozwoju pokrewnych demencji, choroby Picka, postępującego porażenia nadjądrowego czy degeneracji kory mózgowej. Transkrypty kodowanego na tym chromosomie, genu MAPT (microtubule-associated protein tau) są na różne sposoby ekspresjonowane w ośrodkowym układzie nerwowym. Proces ten zależy od stopnia dojrzałości oraz rodzaju komórek nerwowych.
Mutanty MAPT wykazują obecność NFT typową dla patologii neurofibrylarnych oraz znaczny ubytek neuronów w ogniskach zapalnych mózgu. Mechanizm odpowiedzialny za śmierć neuronów obserwowaną u myszy transgenicznych, nie jest jeszcze poznany. Prowadzone badania wskazują, iż proces degeneracji neuronów nie jest zapoczątkowany przez apoptozę (tab. 4) [60,61,95].
Tabela 4. Wybrane mysie linie modelowe – białko tau/białka powiązane z białkiem tau [95]
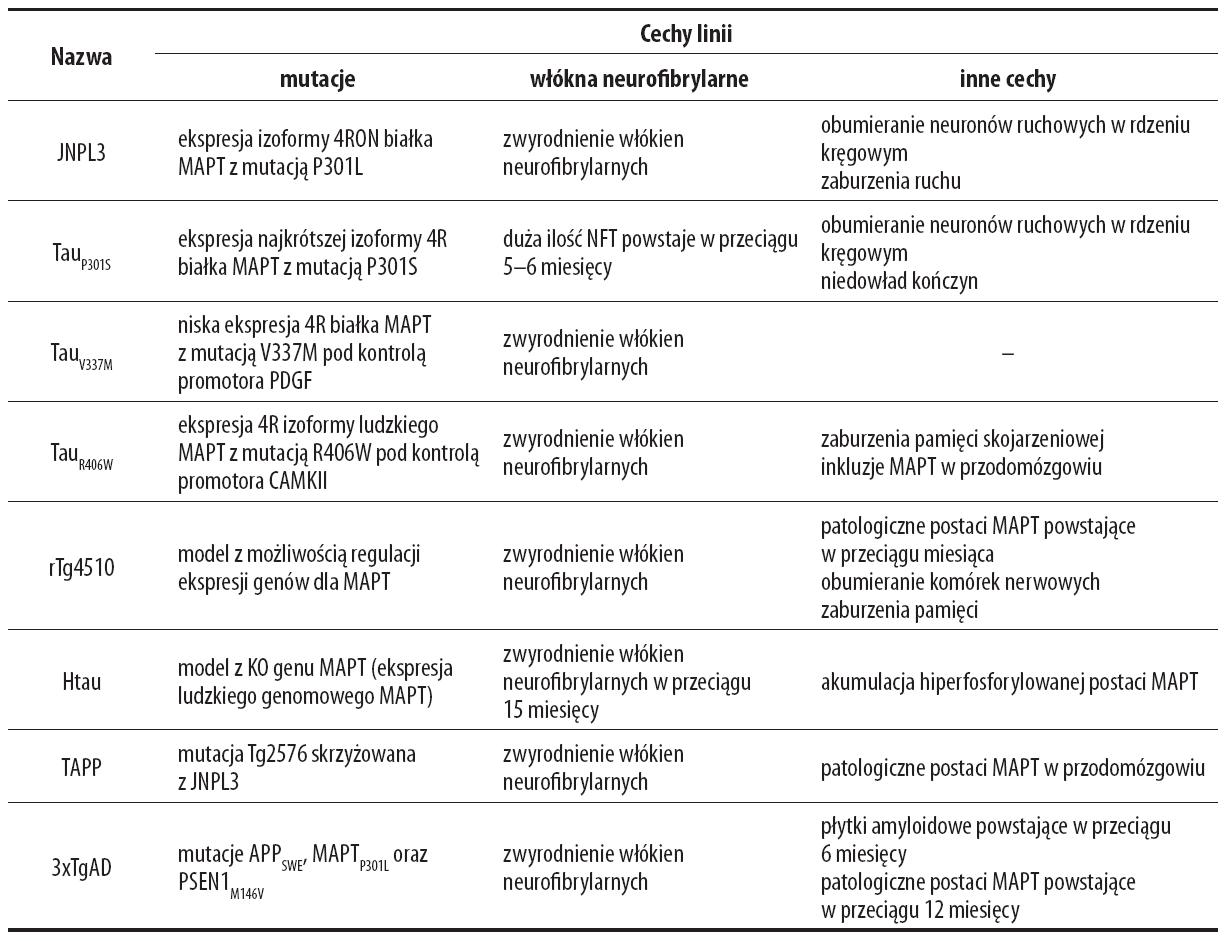
Mysi model rTg4510 odzwierciedlający zaburzenia w metabolizmie białka tau charakteryzuje się:
• ekspresją ludzkiego białka tau P301L ograniczoną do przodomózgowia,
• obecnością srebrochłonnych NFT rozwijających się w ciągu 4 miesięcy,
• upośledzeniem pamięci rozwijającym się w ciągu 2,5 miesiąca,
• całkowitą atrofią mózgu w ciągu 5,5 miesiąca.
Wyprowadzono również linię charakteryzującą się powstawaniem zarówno płytek amyloidowych jak i NFT (JNPL3), która również cechowała się wysokim poziomem obumierania neuronów w węchomózgowiu. Linia JNPL3 stworzona ze skrzyżowania linii PSEN1M146V, APPSWE oraz MAPT wykazuje pogłębiające się z wiekiem: wewnątrzneuronalne występowanie Aβ, pojedyncze złogi amyloidowe, ubytki białka MAPT, dysfunkcję synaps oraz zaburzenia pamięci. Możliwość stworzenia modelu mysiego odzwierciedlającego wszystkie zmiany patologiczne związane z AD wydaje się bardzo skomplikowana i ograniczona żywotnością oraz wielowymiarowym mechanizmem oddziaływań Aβ z innymi zmianami patologicznymi cechującymi AD [61,95].
Mimo iż podstawowymi modelami stosowanymi w badaniach mechanizmów AD są modele mysie, prowadzi się również doświadczenia na muszkach owocówkach (Drosophila melanogaster), nicieniach (Caenorhabditis elegans) czy minogach morskich (Petromyzon marinus). Zaletami D. melanogaster są: niewielki rozmiar organizmów, krótki czas życia oraz niskie koszta prowadzenia badań. Model ten jest używany przede wszystkim do badania zmian apoptotycznych oraz zaburzeń mitozy w AD. Ekspresje dzikiego typu oraz mutacje ludzkiego białka tau u C. elegans oraz D. melanogaster objawiają się patologicznymi zaburzeniami na poziomie synaptycznym oraz zmianami w zachowaniu. P. marinus charakteryzuje się obecnością w móżdżku sześciu, dużych, mocno rozgałęzionych neuronów, które są wdzięcznym modelem do badań [60].
3.3. Banki mózgów
Doświadczenia ostatnich dziesięciu lat wskazują, że podstawą dogłębnego zrozumienia funkcjonowania ośrodkowego układu nerwowego jest badanie tkanek mózgowych. Wyniki doświadczeń prowadzonych post mortem odegrały znaczącą rolę w opracowaniu testu genetycznego na chorobę Huntingtona i Parkinsona. Również neurochemiczne i anatomiczne badania mózgu stwarzają nowe możliwości zrozumienia mechanizmów leżących u podstaw psychozy oraz innych chorób mózgu. By móc analizować stan patologiczny, niezbędne jest porównanie wyników z kontrolnymi, zdrowymi tkankami mózgowymi pochodzącymi od dawców, którzy nie mieli urazów głowy, ataku serca, zaników pamięci, delirium oraz nie byli uzależnieni od leków, papierosów czy alkoholu. Do badań neurobiologicznych tkanki są kolekcjonowane od: • dawców bez zaburzeń neurologicznych czy neuropsychiatrycznych, • dawców ze zdiagnozowanymi zaburzeniami, • schizofreników lub chorych na manie depresyjne, • rodziców, dzieci oraz rodzeństwa chorych.
Istnieją banki specjalizujące się w przechowywaniu tkanek dawców z wybranymi chorobami neurologicznymi. W bankach mózgów są przechowywane tkanki chorych na: AD, stwardnienie zanikowe boczne, autyzm, demencję, depresję, dystonię DYT-1, choroby Huntingtona i Parkinsona, postępujące porażenie nadjądrowe, zespół Retta czy Turetta itp. [107,119,160].
Mimo stosowania wielu różnorodnych modeli badawczych często trudno jest znaleźć jednoznaczną odpowiedź nie tylko na pytanie jak przebiega proces neurodegeneracji, ale również, w jaki sposób i jakimi czynnikami można go spowolnić, zatrzymać lub mu zapobiegać oraz w sposób jednoznaczny wykazać skuteczność proponowanej terapii. Jest to wciąż aktualne wyzwanie do prowadzenia interdyscyplinarnych badań.
PIŚMIENNICTWO
[1] Aarli J.A.: Role of cytokines in neurological disorders. Curr. Med. Chem., 2003; 10: 1931-1937
[PubMed]
[2] Adessi C., Frossard M.J., Boissard C., Fraga S., Bieler S., Ruckle T., Vilbois F., Robinson S.M., Mutter M., Banks W.A., Soto C.: Pharmacological profiles of peptide drug candidates for the treatment of Alzheimer’s disease. J. Biol. Chem., 2003; 278: 13905-13911
[PubMed] [Full Text HTML] [Full Text PDF]
[3] Akiyama H., Barger S., Barnum S., Bradt B., Bauer J., Cole G.M., Cooper N.R., Eikelenboom P., Emmerling M., Fiebich B.L., Finch C.E., Frautschy S., Griffin W.S., Hampel H., Hull M., Landreth G., Lue L., Mrak R., Mackenzie I.R., McGeer P.L., O’Banion M.K., Pachter J., Pasinetti G., Plata-Salaman C., Rogers J., Rydel R., Shen Y., Streit W., Strohmeyer R., Tooyoma I., Van Muiswinkel F.L., Veerhuis R., Walker D., Webster S., Wegrzyniak B., Wenk G., Wyss-Coray T.: Inflammation and Alzheimer’s disease. Neurobiol. Aging, 2000; 21: 383-421
[PubMed]
[4] Alkam T., Nitta A., Mizoguchi H., Itoh A., Nabeshima T.: A natural scavenger of peroxynitrites, rosmarinic acid, protects against impairment of memory induced by Aβ(25-35). Behav. Brain Res., 2007; 180: 139-145
[PubMed]
[5] Allan S.M., Rothwell N.J.: Cytokines and acute neurodegeneration. Nat. Rev. Neurosci., 2001; 2: 734-744
[PubMed]
[6] Ancelin M.L., Christen Y., Ritchie K.: Is antioxidant therapy a viable alternative for mild cognitive impairment? Examination of the evidence. Dement. Geriatr. Cogn. Disord., 2007; 24: 1-19
[PubMed]
[7] Arbel M., Yacoby I., Solomon B.: Inhibition of amyloid precursor protein processing by β-secretase through site-directed antibodies. Proc. Natl. Acad. Sci. USA, 2005; 102: 7718-7723
[PubMed] [Full Text HTML] [Full Text PDF]
[8] Arosio B., Trabattoni D., Galimberti L., Bucciarelli P., Fasano F., Calabresi C., Cazzullo C.L., Vergani C., Annoni G., Clerici M.: Interleukin-10 and interleukin-6 gene polymorphisms as risk factors for Alzheimer’s disease. Neurobiol. Aging, 2004; 25: 1009-1015
[PubMed]
[9] Aslan M., Ozben T.: Reactive oxygen and nitrogen species in Alzheimer’s disease. Curr. Alzheimer Res., 2004; 1: 111-119
[PubMed]
[10] Babas T., Munoz D., Mankowski J.L., Tarwater P.M., Clements J.E., Zink M.C.: Role of microglial cells in selective replication of simian immunodeficiency virus genotypes in the brain. J. Virol., 2003; 77: 208-216
[PubMed] [Full Text HTML] [Full Text PDF]
[11] Bacsi A., Stanton G.J., Hughes T.K., Kruzel M., Boldogh I.: Colostrinin-driven neurite outgrowth requires p53 activation in PC12 cells. Cell. Mol. Neurobiol., 2005; 25: 1123-1139
[PubMed]
[12] Beckman M.: Untangling Alzheimer’s by paring plaques bolsters amyloid theory. Science, 2004; 305: 762
[PubMed]
[13] Beloosesky Y., Salman H., Bergman M., Bessler H., Djaldetti M.: Cytokine levels and phagocytic activity in patients with Alzheimer’s disease. Gerontology, 2002; 48: 128-132
[PubMed]
[14] Benveniste E.N.: Inflammatory cytokines within the central nervous system: sources, function, and mechanism of action. Am. J. Physiol. Cell Physiol., 1992; 263: C1-C16
[PubMed]
[15] Bilikiewicz A., Gaus W.: Colostrinin (a naturally occurring, proline-rich, polypeptide mixture) in the treatment of Alzheimer’s disease. J. Alzheimers Dis., 2004; 6: 17-26
[PubMed]
[16] Birks J.: Cholinesterase inhibitors for Alzheimer’s disease. Cochrane Database Syst. Rev., 2006; 1: CD005593
[PubMed] [Full Text PDF]
[17] Birnie G.D.: The HL60 cell line: a model system for studying human myeloid cell differentiation. Br. J. Cancer Suppl., 1988; 9: 41-45
[PubMed]
[18] Blasko I., Apochal A., Boeck G., Hartmann T., Grubeck-Loebenstein B., Ransmayr G.: Ibuprofen decreases cytokine-induced amyloid β production in neuronal cells. Neurobiol. Dis., 2001; 8: 1094-1101
[PubMed]
[19] Blasko I., Grubeck-Loebenstein B.: Role of the immune system in the pathogenesis, prevention and treatment of Alzheimer’s disease. Drugs Aging, 2003; 20: 101-113
[PubMed]
[20] Bonotis K., Krikki E., Holeva V., Aggouridaki C., Costa V., Baloyannis S.: Systemic immune aberrations in Alzheimer’s disease patients. J. Neuroimmunol., 2008; 193:183-187
[PubMed]
[21] Braddock M.: Safely slowing down the decline in Alzheimer’s disease: gene therapy shows potential. Expert Opin. Investig. Drugs, 2005; 14: 913-915
[PubMed]
[22] Brera B., Serrano A., de Ceballos M.L.: β-amyloid peptides are cytotoxic to astrocytes in culture: a role for oxidative stress. Neurobiol. Dis., 2000; 7: 395-405
[PubMed]
[23] Brzyska M., Elbaum D.: Dysregulation of calcium in Alzheimer’s disease. Acta Neurobiol. Exp., 2003; 63: 171-183
[PubMed]
[24] Bucki R., Górski J.: Współczesne poglądy dotyczące funkcji i regulacji stężenia Ca2+ w jądrach komórkowych. Post. Hig. Med. Dośw., 2001; 55: 157-175
[PubMed]
[25] Bullock R.: Galantamine: use in Alzheimer’s disease and related disorders. Expert Rev. Neurother., 2004; 4: 153-163
[PubMed]
[26] Calabrese V., Bates T.E., Stella A.M.: NO synthase and NO-dependent signal pathways in brain aging and neurodegenerative disorders: the role of oxidant/antioxidant balance. Neurochem. Res., 2000; 25: 1315-1341
[PubMed]
[27] Caselli R.J., Beach T.G., Yaari R., Reiman E.M.: Alzheimer’s disease a century later. J. Clin. Psychiatry, 2006; 67: 1784-1800
[PubMed]
[28] Chen C.Y., Jang J.H., Park M.H., Hwang S.J., Surh Y.J., Park O.J.: Attenuation of Aβ-induced apoptosis of plant extract (Saengshik) mediated by the inhibition of mitochondrial dysfunction and antioxidative effect. Ann. NY Acad. Sci., 2007; 1095: 399-411
[PubMed]
[29] Cho H.S., Hyman B.T., Greenberg S.M., Rebeck G.W.: Quantitation of apoE domains in Alzheimer disease brain suggests a role for apoE in Aβ aggregation. J. Neuropathol. Exp. Neurol., 2001; 60: 342-349
[PubMed]
[30] Chong Y.H., Shin Y.J., Suh Y.H.: Cyclic AMP inhibition of tumor necrosis factor α production induced by amyloidogenic C-terminal peptide of Alzheimer’s amyloid precursor protein in macrophages: involvement of multiple intracellular pathways and cyclic AMP response element binding protein. Mol. Pharmacol., 2003; 63: 690-698
[PubMed] [Full Text HTML] [Full Text PDF]
[31] Christensen D.D.: Alzheimer’s disease: progress in the development of anti-amyloid disease-modifying therapies. CNS Spectr., 2007; 12: 113-123
[PubMed]
[32] Chu L.W., Yik P.Y., Mok W., Chung C.P.: A 2-year open-label study of galantamine therapy in Chinese Alzheimer’s disease patients in Hong Kong. Int. J. Clin. Pract., 2007; 61: 403-410
[PubMed] [Full Text HTML] [Full Text PDF]
[33] Collins S.J.: The HL-60 promyelocytic leukemia cell line: proliferation, differentiation, and cellular oncogene expression. Blood, 1987; 70: 1233-1244
[PubMed] [Full Text PDF]
[34] Combs C.K., Karlo J.C., Kao S.C., Landreth G.E.: β-amyloid stimulation of microglia and monocytes results in TNFα-dependent expression of inducible nitric oxide synthase and neuronal apoptosis. J. Neurosci., 2001; 21: 1179-1188
[PubMed] [Full Text HTML] [Full Text PDF]
[35] Corzo L., Zas R., Rodriguez S., Fernández-Novoa L., Cacabelos R.: Decreased levels of serum nitric oxide in different forms of dementia. Neurosci. Lett., 2007; 420: 263-267
[PubMed]
[36] Craft J.M., Watterson D.M., Van Eldik L.J.: Human amyloid β-induced neuroinflammation is an early event in neurodegeneration. Glia, 2006; 53: 484-490
[PubMed]
[37] De Felice F.G., Velasco P.T., Lambert M.P., Viola K., Fernandez S.J., Ferreira S.T., Klein W.L.: A β oligomers induce neuronal oxidative stress through an N-methyl-D-aspartate receptor-dependent mechanism that is blocked by the Alzheimer drug memantine. J. Biol. Chem., 2007; 282: 11590-11601
[PubMed] [Full Text HTML] [Full Text PDF]
[38] Dobryszycka W., Gąsiorowski K., Leszek J.: Metaboliczne podstawy choroby Alzheimera. W: Demencje wieku podeszłego. Patomechanizm i strategie leczenia. Wyd. Continuo, 2004; 9-45
[39] Dobryszycka W., Leszek J., Rymaszewska J.: Blaszki starcze. W: Choroba Alzheimera. Patogeneza, diagnostyka, leczenie. Wyd. Continuo, 2002; 13-25
[40] Eikelenboom P., Veerhuis R., Scheper W., Rozemuller A.J., van Gool W.A., Hoozemans J.J.: The significance of neuroinflammation in understanding Alzheimer’s disease. J. Neural. Transm., 2006; 113: 1685-1695
[PubMed]
[41] Eperon S., Jungi T.W.: The use of human monocytoid lines as indicators of endotoxin. J. Immunol. Methods, 1996; 194: 121-129
[PubMed]
[42] Farlow M.R., Cummings J.L.: Effective pharmacologic management of Alzheimer’s disease. Am. J. Med., 2007; 120: 388-397
[PubMed]
[43] Feigin A., Eidelberg D.: Gene transfer therapy for neurodegenerative disorders. Mov. Disord., 2007; 22: 1223-1228
[PubMed]
[44] Feldman H.H., Ferris S., Winblad B., Sfikas N., Mancione L., He Y., Tekin S., Burns A., Cummings J., del Ser T., Inzitari D., Orgogozo J.M., Sauer H., Scheltens P., Scarpini E., Herrmann N., Farlow M., Potkin S., Charles H.C., Fox N.C., Lane R.: Effect of rivastigmine on delay to diagnosis of Alzheimer’s disease from mild cognitive impairment: the InDDEx study. Lancet Neurol., 2007; 6: 501-512
[PubMed]
[45] Fiala M., Cribbs D.H., Rosenthal M., Bernard G.: Phagocytosis of amyloid-β and inflammation: two faces of innate immunity in Alzheimer’s disease. J. Alzheimers Dis., 2007; 11: 457-463
[PubMed]
[46] Filipcik P., Cente M., Ferencik M., Hulin I., Novak M.: The role of oxidative stress in the pathogenesis of Alzheimer’s disease. Bratisl. Lek. Listy, 2006; 107: 384-394
[PubMed]
[47] Flirski M., Sobow T.: Biochemical markers and risk factors of Alzheimer’s disease. Curr. Alzheimer Res., 2005; 2: 47-64
[PubMed]
[48] Fraser P.E., Yang D.S., Yu G., Lévesque L., Nishimura M., Arawaka S., Serpell L.C., Rogaeva E., St. George-Hyslop P.: Presenilin structure, function and role in Alzheimer disease. Biochim. Biophys. Acta, 2000; 1502: 1-15
[PubMed]
[49] Gadient R.A., Otten U.H.: Interleukin-6 (IL-6) – a molecule with both beneficial and destructive potentials. Prog. Neurobiol., 1997; 52: 379-390
[PubMed]
[50] Gambi F., Reale M., Iarlori C., Salone A., Toma L., Paladini C., De Luca G., Feliciani C., Salvatore M., Salerno R.M., Theoharides T.C., Conti P., Exton M., Gambi D.: Alzheimer patients treated with an AchE inhibitor show higher IL-4 and lower IL-1b levels and expression in peripheral blood mononuclear cells. J. Clin. Psychopharmacol., 2004; 24: 314-321
[PubMed]
[51] Gamblin T.C., Chen F., Zambrano A., Abraha A., Lagalwar S., Guillozet A.L., Lu M., Fu Y., Garcia-Sierra F., LaPointe N., Miller R., Berry R.W., Binder L.I., Cryns V.L.: Caspase cleavage of tau: linking amyloid and neurofibrillary tangles in Alzheimer’s disease. Proc. Natl. Acad. Sci. USA, 2003; 100: 10032-10037
[PubMed] [Full Text HTML] [Full Text PDF]
[52] Gandy S., Petanceska S.: Regulation of Alzheimer β-amyloid precursor trafficking and metabolism. Biochim. Biophys. Acta, 2000; 1502: 44-52
[PubMed]
[53] Gebicke-Haerter P.J.: Microglia in neurodegeneration: molecular aspects. Microsc. Res. Tech., 2001; 54: 47-58
[PubMed]
[54] Geldmacher D.S.: Donepezil (Aricept) for treatment of Alzheimer’s disease and other dementing conditions. Expert Rev. Neurother., 2004; 4: 5-16
[PubMed]
[55] Gelinas D.S., DaSilva K., Fenili D., St. George-Hyslop P., McLaurin J.: Immunotherapy for Alzheimer’s disease. Proc. Natl. Acad. Sci. USA, 2004; 101 (Suppl. 2): 14657-14662
[PubMed] [Full Text HTML] [Full Text PDF]
[56] Gilman S., Koller M., Black R.S., Jenkins L., Griffith S.G., Fox N.C., Eisner L., Kirby L., Rovira M.B., Forette F., Orgogozo J.M.: Clinical effects of Aβ immunization (AN1792) in patients with AD in an interrupted trial. Neurology, 2005; 64: 1553-1562
[PubMed]
[57] Glabe C.: Intracellular mechanisms of amyloid accumulation and pathogenesis in Alzheimer’s disease. J. Mol. Neurosci., 2001; 17: 137-145
[PubMed]
[58] Golde T.E.: Alzheimer disease therapy: can the amyloid cascade be halted? J. Clin. Invest., 2003; 111: 11-18
[PubMed] [Full Text HTML] [Full Text PDF]
[59] Golde T.E.: The Aβ hypothesis: leading us to rationally-designed therapeutic strategies for the treatment or prevention of Alzheimer disease. Brain Pathol., 2005; 15: 84-87
[PubMed]
[60] Götz J.: Tau and transgenic animal models. Brain Res. Brain Res. Rev., 2001; 35: 266-286
[PubMed]
[61] Götz J., Deters N., Doldissen A., Bokhari L., Ke Y., Wiesner A., Schonrock N., Ittner L.M.: A decade of tau transgenic animal models and beyond. Brain Pathol., 2007; 17: 91-103
[PubMed]
[62] Griffin W.S., Stanley L.C., Ling C., White L., MacLeod V., Perrot L.J., White C.L.3rd, Araoz C.: Brain interleukin 1 and S-100 immunoreactivity are elevated in Down syndrome and Alzheimer disease. Proc. Natl. Acad. Sci. USA, 1989; 86: 7611-7615
[PubMed] [Full Text PDF]
[63] Griffin W.S., Liu L., Li Y., Mrak R.E., Barger S.W.: Interleukin-1 mediates Alzheimer and Lewy body pathologies. J. Neuroinflammation, 2006; 3: 5
[PubMed]
[64] Grzanna R., Phan P., Polotsky A., Lindmark L., Frondoza C.G.: Ginger extract inhibits β-amyloid peptide-induced cytokine and chemokine expression in cultured THP-1 monocytes. J. Altern. Complement. Med., 2004; 10: 1009-1013
[PubMed]
[65] Guo J.P., Arai T., Miklossy J., McGeer P.L.: Aβ and tau form soluble complexes that may promote self aggregation of both into the insoluble forms observed in Alzheimer’s disease. Proc. Natl. Acad. Sci. USA, 2006; 103: 1953-1958
[PubMed] [Full Text HTML] [Full Text PDF]
[66] Hardy J.: Amyloid, the presenilins and Alzheimer’s disease. Trends Neurosci., 1997; 20: 154-159
[PubMed]
[67] Hartmann R., Meisel H.: Food-derived peptides with biological activity: from research to food applications. Curr. Opin. Biotechnol., 2007; 18: 163-169
[PubMed]
[68] Heneka M.T., O’Banion M.K.: Inflammatory processes in Alzheimer’s disease. J. Neuroimmunol., 2007; 184: 69-91
[PubMed]
[69] Hock C., Konietzko U., Streffer J.R., Tracy J., Signorell A., Müller-Tillmanns B., Lemke U., Henke K., Moritz E., Garcia E., Wollmer M.A, Umbricht D., de Quervain D.J., Hofmann M., Maddalena A., Papassotiropoulos A., Nitsch R.M.: Antibodies against β-amyloid slow cognitive decline in Alzheimer’s disease. Neuron, 2003; 38: 547-554
[PubMed]
[70] Huang Y., Liu X.Q., Wyss-Coray T., Brecht W.J., Sanan D.A., Mahley R.W.: Apolipoprotein E fragments present in Alzheimer’s disease brains induce neurofibrillary tangle-like intracellular inclusions in neurons. Proc. Natl. Acad. Sci. USA, 2001; 98: 8838-8843
[PubMed] [Full Text HTML] [Full Text PDF]
[71] Hyman B.T., Growdon J.H.: Can the immune system fight Alzheimer disease? Nat. Med., 2006; 12: 755-756
[PubMed]
[72] Ichikawa M., Sugita M., Takahashi M., Satomi M., Takeshita T., Araki T., Takahashi H.: Breast milk macrophages spontaneously produce granulocyte-macrophage colony-stimulating factor and differentiate into dendritic cells in the presence of exogenous interleukin-4 alone. Immunology, 2003; 108: 189-195
[PubMed] [Full Text HTML] [Full Text PDF]
[73] Ischiropoulos H., Beckman J.S.: Oxidative stress and nitration in neurodegeneration: cause, effect, or association? J. Clin. Invest., 2003; 111: 163-169
[PubMed] [Full Text HTML] [Full Text PDF]
[74] Kelso A.: Cytokines: principles and prospects. Immunol. Cell Biol., 1998; 76: 300-317
[PubMed]
[75] Kim D.S., Kim J.Y., Han Y.S.: Alzheimer’s disease drug discovery from herbs: neuroprotectivity from βeta-amyloid (1-42) insult. J. Altern. Complement. Med., 2007; 13: 333-340
[PubMed]
[76] Kitamura T., Sugimori K., Sudo S., Kobayashi K.: Relationship between microtubule-binding repeats and morphology of neurofibrillary tangle in Alzheimer’s disease. Acta Neurol. Scand., 2005; 112: 327-334
[PubMed]
[77] Kivipelto M., Solomon A., Winblad B.: Alzheimer’s Disease: back to the future. Acta Neurol. Scand. Suppl., 2006; 114: 119-120
[PubMed]
[78] Kolarski W.: Podstawowe typy komórek i tkanek. W: Strukturalne podstawy biologii komórki. Wyd. PWN, 2003; 55-60
[79] Kowalska A.: Hipoteza kaskady β-amyloidu – sekwencja wydarzeń prowadzących do neurodegeneracji w chorobie Alzheimera. Neurol. Neurochir. Pol., 2004; 38: 405-411
[PubMed] [Full Text PDF]
[80] Lee H.G., Castellani R.J., Zhu X., Perry G., Smith M.A.: Amyloid-β in Alzeimer’s disease: the horse or the cart? Pathogenic or protective? Int. J. Exp. Pathol., 2005; 86: 133-138
[PubMed]
[81] Lemere C.A.: A beneficial role for IL-1 β in Alzheimer disease? J. Clin. Invest., 2007, 117:1483-1485
[PubMed] [Full Text HTML] [Full Text PDF]
[82] Leong S.K., Ruan R.S., Zhang Z.: A critical assessment of the neurodestructive and neuroprotective effects of nitric oxide. Ann. NY Acad. Sci., 2002; 962: 161-181
[PubMed]
[83] Leszek J., Inglot A.D., Janusz M., Byczkiewicz F., Kiejna A., Georgiades J.A., Lisowski J.: Colostrinin proline-rich polypeptide complex from ovine colostrum – a long-term study of its efficacy in Alzheimer’s disease. Med. Sci. Monit., 2002; 8: PI93-PI96
[PubMed]
[84] Leszek J., Inglot A.D., Janusz M., Lisowski J., Krukowska K., Georgiades J.A.: Colostrinin®: a proline-rich polypeptide (PRP) complex isolated from ovine colostrum for treatment of Alzheimer’s disease. A double-blind, placebo-controlled study. Arch. Immunol. Ther. Exp., 1999; 47: 377-385
[PubMed]
[85] Licastro F., Pedrini S., Caputo L., Annoni G., Davis L.J., Ferri C., Casadei V., Grimaldi L.M.: Increased plasma levels of interleukin-1, interleukin-6 and α1-antichymotrypsin in patients with Alzheimer’s disease: peripheral inflammation or signals from the brain? J. Neuroimmunol., 2000; 103: 97-102
[PubMed]
[86] Lopez J.C.: Neurodegeneration…β-secretase unmasked. Nat. Rev. Neurosci., 2001; 2: 222-223
[87] Lynch T., Cherny R.A., Bush A.I.: Oxidative processes in Alzheimer’s disease: the role of Ab-metal interactions. Exp. Gerontol., 2000; 35: 445-451
[PubMed]
[88] Maccioni R.B., Munoz J.P., Barbeito L.: The molecular bases of Alzheimer’s disease and other neurodegenerative disorders. Arch. Med. Res., 2001; 32: 367-381
[PubMed]
[89] Makrides V., Massie M.R., Feinstein S.C., Lew J.: Evidence for two distinct binding sites for tau on microtubules. Proc. Natl. Acad. Sci. USA, 2004; 101: 6746-6751
[PubMed]
[90] Markesbery W.R., Carney J.M.: Oxidative alterations in Alzheimer’s disease. Brain Pathol., 1999; 9: 133-146
[PubMed]
[91] Marlatt M.W., Webber K.M., Moreira P.I., Lee H.G., Casadesus G., Honda K., Zhu X., Perry G., Smith M.A.: Therapeutic opportunities in Alzheimer disease: one for all or all for one? Curr. Med. Chem., 2005; 12: 1137-1147
[PubMed]
[92] Marx J.: Alzheimer’s disease. A new take on tau. Science, 2007; 316: 1416-1417
[PubMed]
[93] Masters C.L., Cappai R., Barnham K.J., Villemagne V.L.: Molecular mechanisms for Alzheimer’s disease: implications for neuroimaging and therapeutics. J. Neurochem., 2006; 97: 1700-1725
[PubMed] [Full Text HTML] [Full Text PDF]
[94] Mattson M.P.: Pathways towards and away from Alzheimer’s disease. Nature, 2004; 430: 631-639
[PubMed]
[95] McGowan E., Eriksen J., Hutton M.: A decade of modeling Alzheimer’s disease in transgenic mice. Trends Genet., 2006; 22: 281-289
[PubMed]
[96] Medeiros R., Prediger R.D., Passos G.F., Pandolfo P., Duarte F.S., Franco J.L., Dafre A.L., Di Giunta G., Figueiredo C.P., Takahashi R.N., Campos M.M., Calixto J.B. Connecting TNF-α signaling pathways to iNOS expression in a mouse model of Alzheimer’s disease: relevance for the behavioral and synaptic deficits induced by amyloid β protein. J. Neurosci., 2007; 27: 5394-5404
[PubMed] [Full Text HTML] [Full Text PDF]
[97] Monsonego A., Weiner H.L.: Immunotherapeutic approaches to Alzheimer’s disease. Science, 2003; 302: 834-838
[PubMed]
[98] Mooren F.C., Kinne R.K.: Cellular calcium in health and disease. Biochim. Biophys. Acta, 1998; 1406: 127-151
[PubMed]
[99] Morinaga A., Hirohata M., Ono K., Yamada M.: Estrogen has anti-amyloidogenic effects on Alzheimer’s β-amyloid fibrils in vitro. Biochem. Biophys. Res. Commun., 2007; 359: 697-702
[PubMed]
[100] Morris J.C.: Is Alzheimer’s disease inevitable with age?: Lessons from clinicopathologic studies of healthy aging and very mild Alzheimer’s disease. J. Clin. Invest., 1999; 104: 1171-1173
[PubMed] [Full Text HTML] [Full Text PDF]
[101] Mrak R.E., Griffin W.S.: Interleukin-1, neuroinflammation, and Alzheimer’s disease. Neurobiol. Aging, 2001; 22: 903-908
[PubMed]
[102] Munoz L., Ranaivo H.R., Roy S.M., Hu W., Craft J.M., McNamara L.K., Chico L.W., Van Eldik L.J., Watterson D.M.: A novel p38α MAPK inhibitor suppresses brain proinflammatory cytokine up-regulation and attenuates synaptic dysfunction and behavioral deficits in an Alzheimer’s disease mouse model. J. Neuroinflammation, 2007; 4: 21
[PubMed] [Full Text HTML] [Full Text PDF]
[103] Murphy R.M.: Peptide aggregation in neurodegenerative disease. Annu. Rev. Biomed. Eng., 2002; 4: 155-174
[PubMed]
[104] Nagele R.G., D’Andrea M.R., Lee H., Venkataraman V., Wang H.Y.: Astrocytes accumulate Aβ42 and give rise to astrocytic amyloid plaques in Alzheimer disease brains. Brain Res., 2003; 971: 197-209
[PubMed]
[105] Nakamura Y.: Regulating factors for microglial activation. Biol. Pharm. Bull., 2002; 25: 945-953
[PubMed] [Full Text PDF]
[106] Napryeyenko O., Borzenko I.: Ginkgo biloba special extract in dementia with neuropsychiatric features. A randomised, placebo-controlled, double-blind clinical trial. Arzneimittelforschung, 2007; 57: 4-11
[PubMed]
[107] New York Brain Bank at Columbia University (19.06.2008) http://nybb.hs.columbia.edu
[108] Octave J.N., Essalmani R., Tasiaux B., Menager J., Czech C., Mercken L.: The role of presenilin-1 in the γ-secretase cleavage of the amyloid precursor protein of Alzheimer’s disease. J. Biol. Chem., 2000; 275: 1525-1528
[PubMed] [Full Text HTML] [Full Text PDF]
[109] Oliveira A.A. Jr., Hodges H.M.: Alzheimer’s disease and neural transplantation as prospective cell therapy. Curr. Alzheimer Res., 2005; 2: 79-95
[PubMed]
[110] Orellana D.I., Quintanilla R.A., Maccioni R.B.: Neuroprotective effect of TNF α against the β-amyloid neurotoxicity mediated by CDK5 kinase. Biochim. Biophys. Acta, 2007; 1773: 254-263
[PubMed]
[111] Ostrowski M., Grzanka A., Izdebska M.: Rola aktyny w chorobie Alzheimera. Post. Hig. Med. Dośw., 2005; 59: 224-228
[PubMed] [Full Text HTML] [Full Text PDF]
[112] Ott B.R., Blake L.M., Kagan E., Resnick M.: Open label, multicenter, 28-week extension study of the safety and tolerability of memantine in patients with mild to moderate Alzheimer’s disease. J. Neurol., 2007; 254: 351-358
[PubMed]
[113] Panda D., Samuel J.C., Massie M., Feinstein S.C., Wilson L.: Differential regulation of microtubule dynamics by three- and four-repeat tau: implications for the onset of neurodegenerative disease. Proc. Natl. Acad. Sci. USA, 2003; 100: 9548-9553
[PubMed] [Full Text HTML] [Full Text PDF]
[114] Pappolla M.A., Smith M.A., Bryant-Thomas T., Bazan N., Petanceska S., Perry G., Thal L.J., Sano M., Refolo L.M.: Cholesterol, oxidative stress, and Alzheimer’s disease: expanding the horizons of pathogenesis. Free Radic. Biol. Med., 2002; 33: 173-181
[PubMed]
[115] Pearson H.A., Peers C.: Physiological roles for amyloid β peptides. J. Physiol., 2006; 575: 5-10
[PubMed] [Full Text HTML] [Full Text PDF]
[116] Pinnix I., Musunuru U., Tun H., Sridharan A., Golde T., Eckman C., Ziani-Cherif C., Ontstead L., Sambamurti K.: A novel γ-secretase assay based on detection of the putative C- terminal fragment-γ of amyloid β protein precursor. J. Biol. Chem., 2001; 276: 481-487
[PubMed] [Full Text HTML] [Full Text PDF]
[117] Potyk D.: Treatments for Alzheimer disease. South. Med. J., 2005; 98: 628-635
[PubMed]
[118] Prada C.M., Garcia-Alloza M., Betensky R.A., Zhang-Nunes S.X., Greenberg S.M., Bacskai B.J., Frosch M.P.: Antibody-mediated clearance of amyloid-β peptide from cerebral amyloid angiopathy revealed by quantitative in vivo imaging. J. Neurosci., 2007; 27:1973-1980
[PubMed] [Full Text HTML] [Full Text PDF]
[119] Primate Brain Bank (19.06.2008)
http://www-vf.bio.uu.nl/~webmanager/PBB/PBB-Start.html
[120] Quan N., Herkenham M.: Connecting cytokines and brain: a review of current issues. Histol. Histopathol., 2002; 17: 273-288
[PubMed]
[121] Rajanikant G.K., Zemke D., Kassab M., Majid A.: The therapeutic potential of statins in neurological disorders. Curr. Med. Chem., 2007; 14: 103-112
[PubMed]
[122] Ray S., Britschgi M., Herbert C., Takeda-Uchimura Y., Boxer A., Blennow K., Friedman L.F., Galasko D.R., Jutel M., Karydas A., Kaye J.A., Leszek J., Miller B.L., Minthon L., Quinn J.F., Rabinovici G.D., Robinson W.H., Sabbagh M.N., So Y.T., Sparks D.L., Tabaton M., Tinklenberg J., Yesavage J.A., Tibshirani R., Wyss-Coray T.: Classification and prediction of clinical Alzheimer’s diagnosis based on plasma signaling proteins. Nat. Med., 2007; 13: 1359-1362
[PubMed]
[123] Remarque E.J., Bollen E.L., Weverling-Rijnsburger A.W., Laterveer J.C., Blauw G.J., Westendorp R.G.: Patients with Alzheimer’s disease display a pro-inflammatory phenotype. Exp. Gerontol., 2001; 36: 171-176
[PubMed]
[124] Riekse R.G., Li G., Petrie E.C., Leverenz J.B., Vavrek D., Vuletic S., Albers J.J., Montine T.J., Lee V.M., Lee M., Seubert P., Galasko D., Schellenberg G.D., Hazzard W.R., Peskind E.R.: Effect of statins on Alzheimer’s disease biomarkers in cerebrospinal fluid. J. Alzheimers Dis., 2006; 10: 399-406
[PubMed]
[125] Robak T.: Biologia i farmakologia cytokin. Wydawnictwo PWN, Warszawa, 1995; 13-73,122-133, 156-161, 255-262
[126] Roberson E.D., Scearce-Levie K., Palop J.J., Yan F., Cheng I.H., Wu T., Gerstein H., Yu G.Q., Mucke L.: Reducing endogenous tau ameliorates amyloid β-induced deficits in an Alzheimer’s disease mouse model. Science, 2007; 316: 750-754
[PubMed]
[127] Rogers J., Strohmeyer R., Kovelowski C.J., Li R.: Microglia and inflammatory mechanisms in the clearance of amyloid β peptide. Glia, 2002; 40: 260-269
[PubMed]
[128] Sala G., Galimberti G., Canevari C., Raggi M.E., Isella V., Facheris M., Appollonio I., Ferrarese C.: Peripheral cytokine release in Alzheimer patients: correlation with disease severity. Neurobiol. Aging, 2003; 24: 909-914
[PubMed]
[129] Saura J.: Microglial cells in astroglial cultures: a cautionary note. J. Neuroinflammation, 2007; 4: 26
[PubMed] [Full Text HTML] [Full Text PDF]
[130] Saurwein-Teissl M., Blasko I., Zisterer K., Neuman B., Lang B., Grubeck-Loebenstein B.: An imbalance between pro- and anti-inflammatory cytokines, a characteristic feature of old age. Cytokine, 2000, 12: 1160-1161
[PubMed]
[131] Sayre L.M., Smith M.A., Perry G.: Chemistry and biochemistry of oxidative stress in neurodegenerative disease. Curr. Med. Chem., 2001; 8: 721-738
[PubMed]
[132] Selkoe D.J.: Alzheimer’s disease: genes, proteins, and therapy. Physiol. Rev., 2001; 81: 741-766
[PubMed] [Full Text HTML] [Full Text PDF]
[133] Selkoe D.J.: Presenilin, Notch, and the genesis and treatment of Alzheimer’s disease. Proc. Natl. Acad. Sci. USA, 2001; 98: 11039-11041
[PubMed] [Full Text HTML] [Full Text PDF]
[134] Selkoe D.J.: Alzheimer’s disease is a synaptic failure. Science, 2002; 298: 789-791
[PubMed]
[135] Selkoe D.J., Schenk D.: Alzheimer’s disease: molecular understanding predicts amyloid-based therapeutics. Annu. Rev. Pharmacol. Toxicol., 2003; 43: 545-584
[PubMed]
[136] Seltzer B.: Donepezil: an update. Expert Opin. Pharmacother., 2007; 8: 1011-1023
[PubMed]
[137] Senior K.: Dosing in phase II trial of Alzheimer’s vaccine suspended. Lancet Neurol., 2002; 1: 3
[PubMed]
[138] Serpell L.C.: Alzheimer’s amyloid fibrils: structure and assembly. Biochim. Biophys. Acta, 2000; 1502: 16-30
[PubMed]
[139] Shaftel S.S., Kyrkanides S., Olschowka J.A., Miller J.N., Johnson R.E., O’Banion M.K.: Sustained hippocampal IL-1β overexpression mediates chronic neuroinflammation and ameliorates Alzheimer plaque pathology. J. Clin. Invest., 2007; 117:1595-1604
[PubMed] [Full Text HTML] [Full Text PDF]
[140] Shimazaki M., Nakano H., Kobayashi K.: Correlation between tau phosphorylation sites and tangle morphology in Alzheimer disease. Psychogeriatrics, 2005; 5: 22-35
[141] Siemers E., Skinner M., Dean R.A., Gonzales C., Satterwhite J., Farlow M., Ness D., May P.C.: Safety, tolerability, and changes in amyloid β concentrations after administration of a γ-secretase inhibitor in volunteers. Clin. Neuropharmacol., 2005; 28: 126-132
[PubMed]
[142] Sisodia S.S., Kim S.H., Thinakaran G.: Function and dysfunction of the presenilins. Am. J. Hum. Genet., 1999; 65: 7-12
[PubMed] [Full Text PDF]
[143] Small G.W., Kaufer D., Mendiondo M.S., Quarg P., Spiegel R.: Cognitive performance in Alzheimer’s disease patients receiving rivastigmine for up to 5 years. Int. J. Clin. Pract., 2005; 59: 473-477
[PubMed] [Full Text HTML] [Full Text PDF]
[144] Smith C., Graham D.I., Murray L.S., Nicoll J.A.: Tau immunochemistry in acute brain injury. Neuropathol. Appl. Neurobiol., 2003; 29: 496-502
[PubMed]
[145] Smith M.A., Joseph J.A., Perry G.: Arson. Tracking the culprit in Alzheimer’s disease. Ann. NY Acad. Sci., 2000; 924: 35-38
[PubMed]
[146] Solomon B.: Clinical immunologic approaches for the treatment of Alzheimer’s disease. Expert Opin. Investig. Drugs, 2007; 16: 819-828
[PubMed]
[147] Soto C.: Unfolding the role of protein misfolding in neurodegenerative diseases. Nat. Rev. Neurosci., 2003; 4: 49-60
[PubMed]
[148] Soto C., Sigurdsson E.M., Morelli L., Kumar R.A., Castano E.M., Frangione B.: β-sheet breaker peptides inhibit fibrillogenesis in a rat brain model of amyloidosis: implications for Alzheimer’s therapy. Nat. Med., 1998; 4: 822-826
[PubMed]
[149] Speciale L., Calabrese E., Saresella M., Tinelli C., Mariani C., Sanvito L., Longhi R., Ferrante P.: Lymphocyte subset patterns and cytokine production in Alzheimer’s disease patients. Neurobiol. Aging, 2007; 28: 1163-1169
[PubMed]
[150] Srebro Z., Wiliński B., Sura P.: Stres oksydacyjny w chorobie Alzheimera. Folia Med. Cracov., 2000; 41: 165-170
[PubMed]
[151] Stokłosowa S.: Hodowla komórek i tkanek. Wydawnictwo Naukowe PWN, 2004
[152] Strle K., Zhou J.H., Shen W.H., Broussard S.R., Johnson R.W., Freund G.G., Dantzer R., Kelley K.W.: Interleukin-10 in the brain. Crit. Rev. Immunol., 2001; 21: 427-449
[PubMed]
[153] Sugita-Konishi Y., Pestka J.J.: Differential upregulation of TNF-α, IL-6, and IL-8 production by deoxynivalenol (vomitoxin) and other 8-ketotrichothecenes in a human macrophage model. J. Toxicol. Environ. Health A, 2001; 64: 619-636
[PubMed]
[154] Suh Y.H., Checler F.: Amyloid precursor protein, presenilins, and α-synuclein: molecular pathogenesis and pharmacological applications in Alzheimer’s disease. Pharmacol. Rev., 2002; 54: 469-525
[PubMed] [Full Text HTML] [Full Text PDF]
[155] Sylwanowicz W.: Mały atlas anatomiczny. PZWL, 1969
[156] Szczepanik A.M., Funes S., Petko W., Ringheim G.E.: IL-4, IL-10 and IL-13 modulate A β(1-42)-induced cytokine and chemokine production in primary murine microglia and a human monocyte cell line. J. Neuroimmunol., 2001; 113: 49-62
[PubMed]
[157] Szczyrbowska M., Leszek J.: Wybrane zagadnienia immunologiczne u pacjentów w podeszłym wieku. W: Choroby Otępienne. Teoria i praktyka. Wyd. Continuo, 2003; 243-257
[158] Thomas R.S., Liddell J.E., Murphy L.S., Pache D.M., Kidd E.J.: An antibody to the β-secretase cleavage site on amyloid-β-protein precursor inhibits amyloid-β production. J. Alzheimers Dis., 2006; 10: 379-390
[PubMed]
[159] Tsuchiya S., Yamabe M., Yamaguchi Y., Kobayashi Y., Konno T., Tada K.: Establishment and characterization of a human acute monocytic leukemia cell line (THP-1). Int. J. Cancer, 1980; 26: 171-176
[PubMed]
[160] Verwer R.W., Dubelaar E.J., Hermens W.T., Swaab D.F.: Tissue cultures from adult human postmortem subcortical brain areas. J. Cell. Mol. Med., 2002; 6: 429- 432
[Abstract] [Full Text PDF]
[161] Vetulani J.: Perspektywy terapii choroby Alzheimera. Psychogeriatria Pol., 2004; 1: 253-278
[162] Wallin A.K., Andreasen N., Eriksson S., Batsman S., Nasman B., Ekdahl A., Kilander L., Grut M., Rydén M., Wallin A., Jonsson M., Olofsson H., Londos E., Wattmo C., Eriksdotter Jonhagen M., Minthon L. Swedish Alzheimer Treatment Study Group: Donepezil in Alzheimer’s disease: what to expect after 3 years of treatment in a routine clinical setting. Dement. Geriatr. Cogn. Disord., 2007; 23: 150-160
[PubMed]
[163] Walter J., Kaether C., Steiner H., Haass C.: The cell biology of Alzheimer’s disease: uncovering the secrets of secretases. Curr. Opin. Neurobiol., 2001; 11: 585-590
[PubMed]
[164] Weber M.S., Prod’homme T., Steinman L., Zamvil S.S.: Drug insight: using statins to treat neuroinflammatory disease. Nat. Clin. Pract. Neurol., 2005; 1: 106-112
[PubMed]
[165] Weiner H.L., Frenkel D.: Immunology and immunotherapy of Alzheimer’s disease. Nat. Rev. Immunol., 2006; 6: 404-416
[PubMed]
[166] Weksler M.E., Gouras G., Relkin N.R., Szabo P.: The immune system, amyloid-β peptide, and Alzheimer’s disease. Immunol. Rev., 2005; 205: 244-256
[PubMed]
[167] Williams D.R.: Tauopathies: classification and clinical update on neurodegenerative diseases associated with microtubule-associated protein tau. Intern. Med. J., 2006; 36: 652-660
[PubMed]
[168] Woodhouse A., Dickson T.C., Vickers J.C.: Vaccination strategies for Alzheimer’s disease: A new hope? Drugs Aging, 2007; 24: 107-119
[PubMed]
[169] Wu J., Parungo C., Wu G., Kang P.M., Laham R.J., Sellke F.W., Simons M., Li J.: PR39 inhibits apoptosis in hypoxic endothelial cells: role of inhibitor apoptosis protein-2. Circulation, 2004; 109: 1660-1667
[PubMed] [Full Text HTML] [Full Text PDF]
[170] Wyska E., Rosiak M.: Zastosowanie łańcuchowej reakcji polimerazy w czasie rzeczywistym (real-time PCR) w badaniach farmakokinetycznych. Post. Hig. Med. Dośw., 2006; 60: 660-666
[PubMed] [Full Text HTML] [Full Text PDF]
[171] Yanagisawa K.: Cholesterol and pathological processes in Alzheimer’s disease. J. Neurosci. Res., 2002; 70: 361-366
[PubMed]
[172] Yeon S.W., Jeon Y.J., Hwang E.M., Kim T.Y.: Effects of peptides derived from BACE1 catalytic domain on APP processing. Peptides, 2007; 28: 838-844
[PubMed]
[173] Yuyama K., Yamamoto N., Yanagisawa K.: Chloroquine-induced endocytic pathway abnormalities: Cellular model of GM1 ganglioside-induced Aβ fibrillogenesis in Alzheimer’s disease. FEBS Lett., 2006; 580: 6972-6976
[PubMed]
[174] Žerovnik E.: Amyloid-fibril formation. Proposed mechanisms and relevance to conformational disease. Eur. J. Biochem., 2002; 269: 3362-3371
[PubMed] [Full Text HTML] [Full Text PDF]