
Odsłona tajemnic komórki bakteryjnej – zastosowanie nowych technik mikroskopii fluorescencyjnej
Magdalena Donczew 1 , Katarzyna Ginda 1 , Jolanta Zakrzewska-Czerwińska 2 , Dagmara Jakimowicz 2Streszczenie
W pracy przedstawiono jak rozwój znaczników fluorescencyjnych, począwszy od odkrycia białka zielonej fluorescencji i jego różnobarwnych odmian, aż do opracowania metod analizy oddziaływań międzycząsteczkowych w oparciu o różne warianty białek wpłynął na zrewidowanie poglądów dotyczących budowy i organizacji komórki bakteryjnej. Użycie nowych metod mikroskopowych umożliwiło lokalizację poszczególnych białek oraz fragmentów chromosomu, a także śledzenie ich migracji w czasie rzeczywistym. Badania te ujawniły przestrzenną organizację komórki bakteryjnej obejmującą swoistą subkomórkową lokalizację białek, obecność dynamicznych struktur cytoszkieletowych, uporządkowaną i aktywną segregację chromosomów oraz czasowo-przestrzenną regulację ekspresji genów.
Słowa kluczowe:mikroskopia fluorescencyjna • białka fluorescencyjne • FRET • FRAP • cytoszkielet bakteryjny • organizacja komórki bakteryjnej
Summary
We have reviewed how the development of fluorescent markers, triggered by the discovery of green fluorescence protein and its other color variants leading to the establishment of methods for studies of protein interactions with application of fluorescent proteins, affected the view of bacterial cell organization. Application of the new microscopic methods allowed localization of proteins and chromosomal regions, and observation of their migration in real time. These studies revealed the spatial organization of bacterial cells which includes specific subcellular localization of proteins, the presence of dynamic cytoskeletal structures, orchestrated and active segregation of chromosomes, and spatiotemporal gene regulation.
Key words:fluorescence microscopy • fluorescent proteins • FRET • FRAP • bacterial cytoskeleton • organization of bacterial cell
Wstęp
Ostatnie lata przyniosły radykalną zmianę poglądów na temat budowy i struktury komórki bakteryjnej. Przyczyniły się do tego przede wszystkim najnowsze techniki mikroskopowe, a szczególnie wykorzystanie zjawiska fluorescencji. Zastosowanie barwników fluorescencyjnych wiążących DNA (DAPI czy jodek propidyny), błonę komórkową i peptydoglikan (lipofilowy barwnik FM6-64 oraz znakowane wankomycyną lub aglutyniną) umożliwiło wizualizację struktur subkomórkowych, odpowiednio nukleoidu, błony i ściany komórkowej.Natomiast zastosowanie znakowanych przeciwciał pozwoliło na subkomórkową detekcję białek i ich kompleksów (immunolokalizacja). Jednak metoda ta jest obecnie wypierana przez coraz częstsze wykorzystanie białek fluorescencyjnych, jako znaczników do lokalizacji białek fuzyjnych. Mikroskopia fluorescencyjna pozwala zarówno na analizę preparatów utrwalonych, jak i obserwacje przyżyciowe umożliwiające śledzenie migracji poszczególnych białek oraz fragmentów chromosomu w czasie rzeczywistym. Wyniki uzyskane w tych eksperymentach zostały uzupełnione badaniami biochemicznymi oraz strukturalnymi. Dzięki nim wiadomo, że bakteria nie jest, jak długi czas uważano, „zbiornikiem” wypełnionym mieszaniną białek, ale komórką o ściśle uporządkowanej organizacji, w której umiejscowienie białek i ich kompleksów podlega precyzyjnej czasowo-przestrzennej kontroli sprawowanej przez struktury białkowe o charakterze cytoszkieletowym [50]. Są one tworzone przez homologi białek cytoszkieletu eukariotycznego i odpowiadają, podobnie jak u Eukariota, za utrzymanie kształtu komórki (CreS, homolog filamentów pośrednich nadający księżycowaty kształt Caulobacter crescentus czy MreB, homolog aktyny zapewniający wydłużony kształt bakteriom pałeczkowatym), za podział komórkowy (FtsZ, homolog tubuliny inicjujący powstanie przegrody podziałowej i MinD kontrolujące jej pozycje) oraz za właściwą segregację chromosomów (ParA, zapewniające aktywny rozdział chromosomów po replikacji) [32,41]. Badania wewnątrzkomórkowych struktur u bakterii mają nie tylko znaczenie poznawcze, ale również praktyczne, ponieważ dostarczają informacji na temat możliwych sposobów kontroli wzrostu i namnażania bakterii. Przykładem może być FtsZ, które jest obecnie badane jako potencjalny cel dla antybiotyków.
Początki badań nad lokalizacją białek bakteryjnych
Pierwsze doniesienia wskazujące na swoistą lokalizację określonych białek w komórce bakteryjnej pochodzą z lat 90. ubiegłego wieku. Używając mikroskopii immunoelektronowej wykazano, że białko FtsZ jest umiejscowione w środku komórki, czyli w miejscu powstawania przegrody podziałowej [7]. Natomiast dzięki zastosowaniu mikroskopii fluorescencyjnej w połączeniu z technikami immunofluorescencyjnymi wykazano swoistą lokalizację białek kompleksu chemoreceptora na biegunie komórki Escherichia coli [34]. Metody immunologiczne, chociaż nie ingerują w strukturę przestrzenną białka, są czasochłonne, a także kosztowne, ponieważ wymagają wysokiej jakości przeciwciał: pierwszorzędowego, które swoiście rozpoznaje badane białko oraz drugorzędowego, znakowanego np. barwnikiem fluorescencyjnym. Co więcej, komórki muszą zostać utrwalone, a struktura ich błony i ściany komórkowej naruszona tak, aby przeciwciało mogło przeniknąć do wnętrza. Uniemożliwia to prowadzenie obserwacji w czasie rzeczywistym z zastosowaniem żywych komórek.
Zastosowanie białka zielonej fluorescencji GFP (green fluorescent protein) w biologii komórki rozpoczęło rozwój technologii znaczników fluorescencyjnych. To nieduże (25,9 kDa) fluorescencyjne białko, występujące naturalnie u meduzy Aequorea victoria, odkryli już w 1962 r. Shimomura i wsp. [53], jednak dopiero 30 lat później zastosowano je jako znacznik biologiczny. GFP jest zbudowane głównie z beta-harmonijek tworzących strukturę beta-baryłki, w środku której znajduje się chromofor zbudowany z trzech aminokwasów Ser65-Tyr66-Gly67 poddanych posttranslacyjnej cyklizacji i oksydacji (ryc. 1).
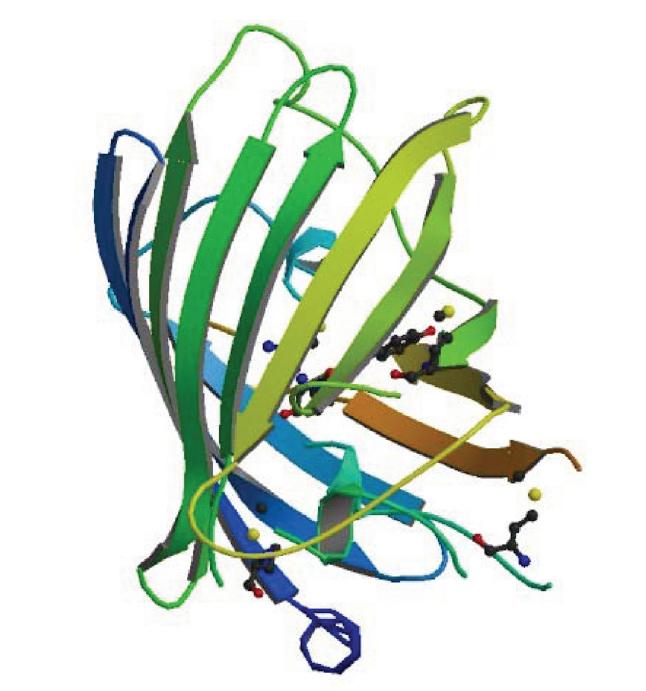
Ryc. 1. Struktura przestrzenna białka zielonej fluorescencji [ID: 1EMA, PDB]
Tworzenie chromoforu trwa około 2 godzin, a jego najwolniejszym i limitującym etapem jest oksydacja. Proces ten nie wymaga obecności żadnego enzymu, również do samej fluorescencji nie są potrzebne dodatkowe kofaktory bądź substraty. Fluorescencja jest zazwyczaj odporna na wygaszanie światłem. Główną zaletą zastosowania GFP jest możliwość obserwacji białek, kompleksów oraz określonych fragmentów chromosomów w żywej komórce w czasie rzeczywistym, a także to, że GFP i jego pochodne nie są toksyczne dla bakterii i organizmów eukariotycznych. Natomiast wadą GFP jest tendencja do tworzenia nierozpuszczalnych agregatów, zwłaszcza w wyższych temperaturach. Ponadto fuzja z GFP może zaburzać strukturę natywną analizowanego białka, a tym samym wpływać na zmianę jego funkcjonalności i lokalizacji. Najczęściej w celu zachowania funkcjonalności obu białek wstawia się pomiędzy nie krótki łącznik (składający się z kilku, kilkunastu aminokwasów), który umożliwia niezależne fałdowanie się obu białek. W 1994 r. Chalfie i wsp. po raz pierwszy poddali ekspresji gen gfp w komórkach bakteryjnych (Escherichia coli) oraz eukariotycznych (nicień Caenorhabditis elegans) i zastosowali go jako gen reporterowy do analizy aktywności promotorów [11]. Rok później zespół Losicka [3] wykorzystał GFP do lokalizacji białka SpoIIE, które jest zaangażowane w proces sporulacji Bacillus subtilis. Wykazano, że umiejscowienie GFP-SpoIIE zmienia się w czasie trwania cyklu komórkowego – przed rozpoczęciem tworzenia asymetrycznej przegrody obserwowano dwa skupiska fluorescencyjne, a po jej utworzeniu już tylko jedno, w miejscu powstawania przegrody.
Jednym z pierwszych interesujących doświadczeń z użyciem fuzyjnego białka GFP była wizualizacja w komórkach E. coli wspomnianego już białka FtsZ – strukturalnego homologu eukariotycznej tubuliny oraz FtsA. FtsZ wyznacza miejsce podziału komórki bakteryjnej poprzez polimeryzację w postaci wewnątrzkomórkowego pierścienia Z, związanego z błoną komórkową (ryc. 2), natomiast białko FtsA jest zasocjowane z powstającym pierścieniem Z [33]. Jednak obecność w komórce samych białek fuzyjnych nie była wystarczająca do prawidłowej lokalizacji i polimeryzacji FtsZ; funkcjonalne polimery białek fuzyjnych tworzyły się jedynie w obecności niezmodyfikowanych białek.
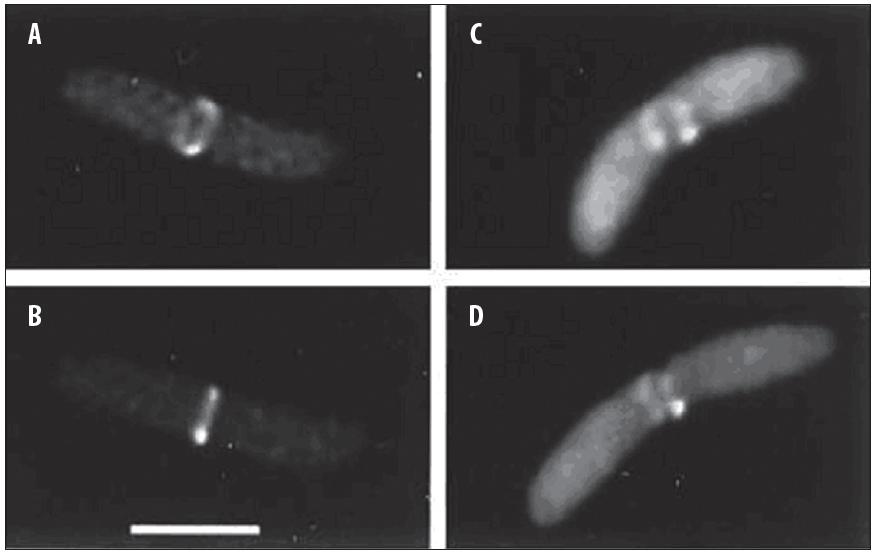
Ryc. 2. Tworzenie pierścienia Z przez białko FtsZ-GFP w centrum komórki E. coli [33, za zgodą]; A, B – obrazy przedstawiające komórkę z w pełni ukształtowanym pierścieniem Z; C, D – obrazy komórki zawierającej niekompletny pierścień Z; skala: 1 µm
W ciągu następnych paru lat, GFP wykorzystano do wizualizacji wielu innych białek [40]. Opisano umiejscowienie w komórkach E. coli lub B. subtilis białek zaangażowanych w główne etapy cyklu komórkowego, na przykład: uczestniczących w podziale komórkowym: MinC [23], MinD [36,47], MinE [22,46], FtsI [63], FtsH [62], FtsL [21,54] FtsQ [12], ZipA [22], odpowiadających za organizację chromosomu HU [64] i SMC [9], zaangażowanych w replikację DNA PolC [24,29] i SeqA [8,38] oraz uczestniczących w segregacji potomnych chromosomów ParA, ParB [31,35,44], FtsK [67] i DivIVA [18]. Określono również lokalizację w komórkach E. coli i Caulobacter crescentus białek CckA [25], Che [55], DivJ i PleC [65], DivK [26] uczestniczących w przekazywaniu sygnału oraz w komórkach B. subtilis CotE [61], DacF [30], SigE [28], SpoIIE [3,5], SpoIIGA [19], SpoIVA [30,42], SpoIVFB zaangażowanych w sporulację. Pierwsze zastosowania GFP ograniczały się głównie do modelowych organizmów – E. coli, B. subtilis, C. crescentus, ale później zaczęto je stosować do lokalizacji białek w komórkach innych gatunków bakterii szczególnie chorobotwórczych np. Mycobacterium tuberculosis, Vibrio cholerae, Helicobacter pylori [45,60,66]. Ciekawym zastosowaniem GFP była wizualizacja cytoszkieletowych białek, krescentyny i Ccrp, które nadają komórkom bakteryjnym charakterystyczny kształt: krescentyna odpowiada za księżycowaty kształt C. crescentus, natomiast białko Ccrp (coiled coil rich protein) za helikalny kształt H. pylori [4,60].
Nowe warianty i kolory białek fluorescencyjnych
Coraz szersze stosowanie GFP sprawiło, że zaczęto poszukiwać jego ulepszonych wariantów. Wprowadzane modyfikacje dotyczyły głównie: zwiększenia intensywności fluorescencji i fotostabilności białka, rozszerzenia zakresu spektralnego, zmniejszenia tendencji do oligomeryzacji i wrażliwości na pH, a także przyspieszenia dojrzewania chromoforu. Pierwszym udoskonalonym białkiem było EGFP (enhanced green fluorescent protein) [14,15], które uzyskano wymieniając jedynie dwa aminokwasy w obrębie chromoforu białka GFP. EGFP wykazuje znacznie wyższą (ponad 35 razy) intensywność fluorescencji w porównaniu do GFP oraz obniżoną tendencję do agregacji w 37°C. Później, głównie przez wprowadzanie przypadkowych mutacji w GFP, uzyskano białka fluorescencyjne emitujące w różnych zakresach spektralnych: niebieskim (blue fluorescent protein – BFP), niebiesko-zielonym (cyan fluorescent protein – CFP) oraz żółtym (yellow fluorescent protein – YFP). Wszystkie wymienione białka mają strukturę białka GFP, a jedyne różnice dotyczą chromoforu.
Największym wyzwaniem w konstrukcji znaczników fluorescencyjnych stało się otrzymanie białek emitujących w zakresie czerwonych i pomarańczowych regionów spektralnych. Wielokrotnie podejmowane próby konstrukcji tych białek w wyniku mutacji wprowadzanych w obrębie chromoforu białka GFP nie dawały żadnych rezultatów. Przełom nastąpił dopiero po odkryciu białka czerwonej fluorescencji (DsRed) pochodzącego z koralowca Discosoma striata [37]. Białko DsRed przyjmuje postać tetrameru. Struktura monomeru DsRed, mimo bardzo niskiej homologii aminokwasowej (~23%), wykazuje wiele podobieństw z konformacją przestrzenną białka GFP. W oparciu o białko DsRed w wyniku losowej i ukierunkowanej mutagenezy, uzyskano warianty, które utraciły zdolność do tetrameryzacji: dTomato, mRFP1, mStrawberry, mPlum i mCherry [49]. Wśród obecnie dostępnych białek fluoryzujących w czerwonym zakresie spektralnym najlepsze własności ma mCherry: charakteryzuje się wysoką fotostabilnością i intensywnością fluorescencji (około połowa intensywności EGFP) oraz może być stosowane w eksperymentach z długim czasem obrazowania [48].
Zastosowanie nowych wariantów białek fluorescencyjnych uruchomiło lawinę doniesień dotyczących już nie tylko swoistej lokalizacji pojedynczych białek, ale przede wszystkim umożliwiło jednoczesną wizualizację kilku białek w komórce. Dzięki wykorzystaniu białek fluorescencyjnych wykazano obecność w komórce bakteryjnej złożonych i dynamicznych struktur białkowych potwierdzając wstępne doniesienia o istnieniu cytoszkieletu bakteryjnego.
W wielu badaniach skupiono się na białkach będących homologami eukariotycznych białek cytoszkieletu – FtsZ, MreB i unikalnym dla bakterii systemie MinCDE, który kontroluje miejsce podziału komórki. Białko MinC, wraz z MinD, hamuje polimeryzację FtsZ i co za tym idzie tworzenie pierścienia Z w pobliżu biegunów komórki, natomiast MinD znosi inhibicję MinCD w centrum komórki. Wykorzystanie dwóch kolorów YFP i CFP umożliwiło np. jednoczesną lokalizację w komórkach E. coli białek MinD, które oscyluje pomiędzy biegunami komórki oraz MinE, które tworzy dynamiczny pierścień (ryc. 3). Dzięki zastosowaniu dwóch białek fluorescencyjnych pokazano, że dynamiczna lokalizacja MinD zależy od MinE, a także zidentyfikowano mutanty MinE, w których oscylacja MinD była zaburzona [51]. Jednoczesną lokalizację MinD-YFP wraz z FtsZ-CFP udało się także zbadać w komórkach mutanta E. coli pozbawionych białka MreB, które zapewnia utrzymanie wydłużonego kształtu komórki [52]. Okazało się, że oscylacja MinD i tworzenie pierścienia FtsZ nie są zaburzone w kulistych mutantach pozbawionych mreB.

Ryc. 3. Umiejscowienie białek MinD oraz MinE w szczepie E. coli wytwarzającym jednocześnie YFP-MinD oraz MinE-CFP [51, za zgodą]. A – Nomarski, B – fluorescencja YFP, C – fluorescencja CFP, D – nakładające się fluorescencje YFP i CFP; skala: 1 µm
Lokalizacja białek MreB i MreC w komórkach Caulobacter crescentus była jednym z pierwszych eksperymentów wykorzystujących znacznik mCherry i mRFP1 do badań biologii komórki bakteryjnej [17]. Na podstawie mikroskopowych obserwacji zaproponowano, że MreB polimeryzuje tworząc spiralny filament, który stanowi rusztowanie pozwalające na odpowiednie rozmieszczenie białek odpowiedzialnych za syntezę ściany komórkowej (ryc. 4). Co więcej, zauważono, że również MreC wykazuje zdolność do tworzenia spiralnych filamentów, które są niezbędne do prawidłowej lokalizacji prekursorów peptydoglikanu.
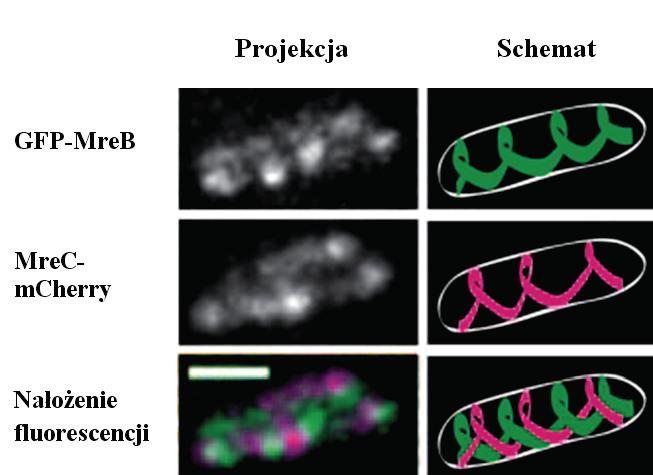
Ryc. 4. Mikroskopia fluorescencyjna białek fuzyjnych GFP-MreB i MreC-mCherry w komórkach C. crescentus [17]. Projekcja to złożenie obrazów z 10 płaszczyzn, schemat – interpretacja graficzna; skala 1 µm
Nowe metody znakowania białek
Jeden z nowych typów znaczników opiera się na wykorzystaniu dwóch domen zamka leucynowego eukariotycznych czynników transkrypcyjnych Jun i Fos (JunLZ, FosLZ). W systemie tym jedna z domen występuje w połączeniu z analizowanym białkiem, natomiast druga z białkiem fluorescencyjnym; utworzenie dimeru pomiędzy JunLZ i FosLZ umożliwia wizualizację analizowanego białka. Największą zaletą stosowania tej metody jest to, że w przeciwieństwie do GFP obie dimeryzujące domeny są dość krótkie (długość JunLZ i FosLZ wynosi odpowiednio 41 i 77, a GFP 240 aminokwasów) i dlatego w znacznie mniejszym stopniu zaburzają funkcjonalność analizowanego białka.
Oddziaływanie domen zamka leucynowego białek Jun i Fos posłużyło do określenia lokalizacji białka SpoIIIE w sporulujących komórkach B. subtilis [43]. SpoIIIE pełni funkcję translokazy wykorzystującej energię pochodzącą z ATP do przeniesienia nowo zreplikowanego chromosomu z komórki macierzystej do prespory. W prowadzonych badaniach próbowano ustalić wpływ obecności subdomeny γ (wchodzącej w skład domeny translokacyjnej białka SpoIIIE) na miejsce wiązania się translokazy do przegrody, a tym samym na kierunek przenoszenia nowo zreplikowanego chromosomu. Analizowano komórki szczepu B. subtilis, w których białko SpoIIIE poddano ekspresji w fuzji z JunLZ, natomiast wytwarzanie znacznika fluorescencyjnego, GFP-FosLZ odbywało się w presporze lub w komórce macierzystej w zależności od badanego szczepu. W przypadku niezmutowanego białka SpoIIIE fluorescencyjne skupiska obserwowano wyłącznie w szczepie, w którym GFP-FosLZ wytwarzane było w komórce macierzystej, a zatem SpoIIIE umiejscowione było po jednej stronie przegrody (ryc. 5A). Natomiast białko SpoIIIE pozbawione subdomeny γ umiejscowione było po obydwu stronach przegrody (ryc. 5B).
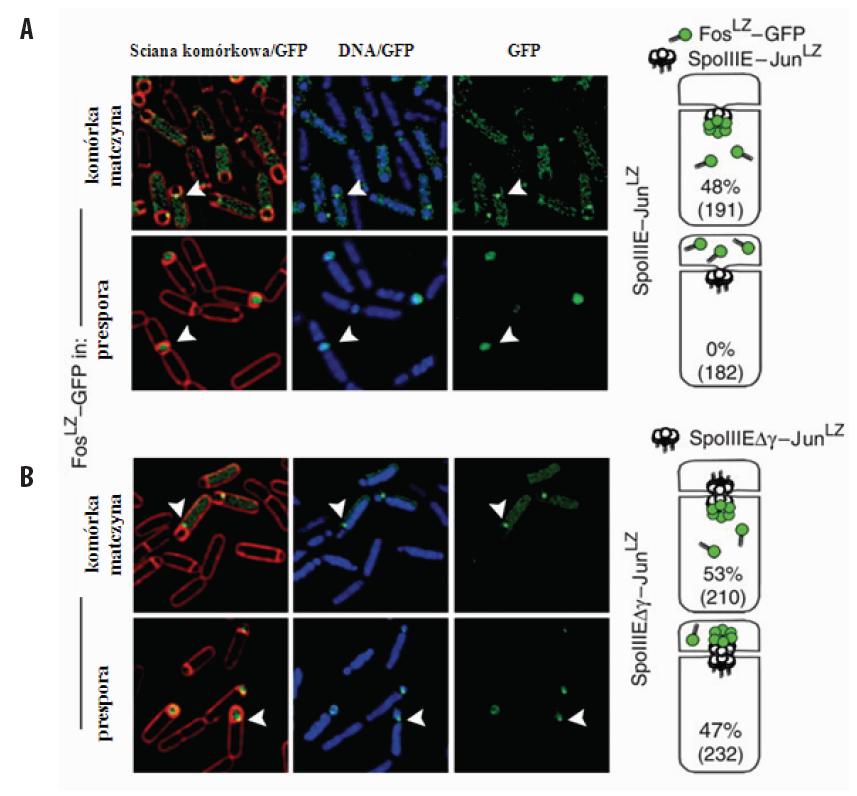
Ryc. 5. Umiejscowienie białka SpoIIIE w sporulujących komórkach B. subtilis [43, za zgodą]. Zdjęcia mikroskopowe oraz schematy (umieszczone z prawej strony) przedstawiają fluorescencję pochodzącą od białka FosLZ-GFP obecnego w szczepach wytwarzających odpowiednio: (A) niezmutowane białko SpoIIIE oraz (B) białko SpoIIIE pozbawione domeny γ. Grotami strzałek oznaczono skupiska GFP, procenty określają liczbę komórek, w których skupiska SpoIIIE-JunLZ obserwowane były po stronie komórki macierzystej lub prespory, n – liczba wszystkich komórek
Obecnie coraz częściej stosowane są syntetyczne barwniki stanowiące alternatywę dla białek fluorescencyjnych, np. FlasH i ReAsH. Barwniki te silnie wiążą sekwencje zawierające cztery reszty cysteinowe (Cys-Cys-X-X-Cys-Cys), a zatem badane białko musi mieć jedynie krótki ogon heksapeptydowy. Największą zaletą stosowania tego typu znaczników jest niewielka zmiana w strukturze trzeciorzędowej analizowanego białka, a także możliwość usunięcia barwnika. Barwniki te znalazły zastosowanie w lokalizacji białka FliC w komórkach E. coli [13]. FliC jest monomerem flagelliny, kurczliwego białka, które polimeryzując tworzy włókna – podstawę konstrukcji rzęski bakteryjnej.
Stosunkowo niedawno wprowadzono znaczniki o ściśle kontrolowanej i odwracalnej fluorescencji [39]; jednym z pierwszych takich białek było PA-GFP (photoactivable – GFP). Znacznik ten stanowi pewnego rodzaju przełącznik molekularny, który z łatwością przechodzi ze stanu „uśpionego” w stan fluorescencyjnie wzbudzony wskutek intensywnego naświetlania. Co więcej białko to może przechodzić nawet kilkaset takich cykli. Dotychczas białko PA-GFP wykorzystano w badaniach nad organizmami eukariotycznymi.
Badanie dynamicznej lokalizacji białek bakteryjnych
Jedną z technik pozwalających na badanie dynamiki/ruchu białek w komórkach jest FRAP (fluorescence recovery after photobleaching). Metoda ta polega na wygaszeniu fluorescencji we fragmencie struktury subkomórkowej zawierającego znakowane fluorescencyjnie białko. Do wygaszenia używa się lasera o wysokiej intensywności, a następnie, w określonym czasie, monitoruje się dyfuzję nowych cząstek fluorescencyjnych w miejscu, w którym wygaszono fluorescencję. Pozwala to na analizę własności kinetycznych i motorycznych białek, współczynnika dyfuzji czy szybkości transportu w żywej komórce. Przywrócenie fluorescencji w wygaszonym miejscu w krótkim czasie oznacza, że w badanej strukturze zachodzą aktywne procesy wymiany. Zastosowanie metody FRAP pozwoliło śledzić zmiany w umiejscowieniu białek w żywych komórkach i znacznie przyspieszyło badania nad dynamiką bakteryjnych białek cytoszkieletowych, np. białka FtsZ, które podobnie jak tubulina ma aktywność GTP-azową i wykazuje zdolność do tworzenia protofilamentów in vitro [2]. Badania metodą FRAP nad znakowanym fluorescencyjnie FtsZ w komórkach E. coli i B. subtilis wykazały, że struktura „pierścienia Z” tworzonego przez białko FtsZ jest niezwykle dynamiczna – połowiczny czas przywracania fluorescencji wynosi 8-9 s u tych bakterii. To oznacza, że następuje intensywna wymiana „wygaszonych” cząsteczek FtsZ-GFP na nowe, pochodzące prawdopodobnie z cytoplazmy (ryc. 6) [56].
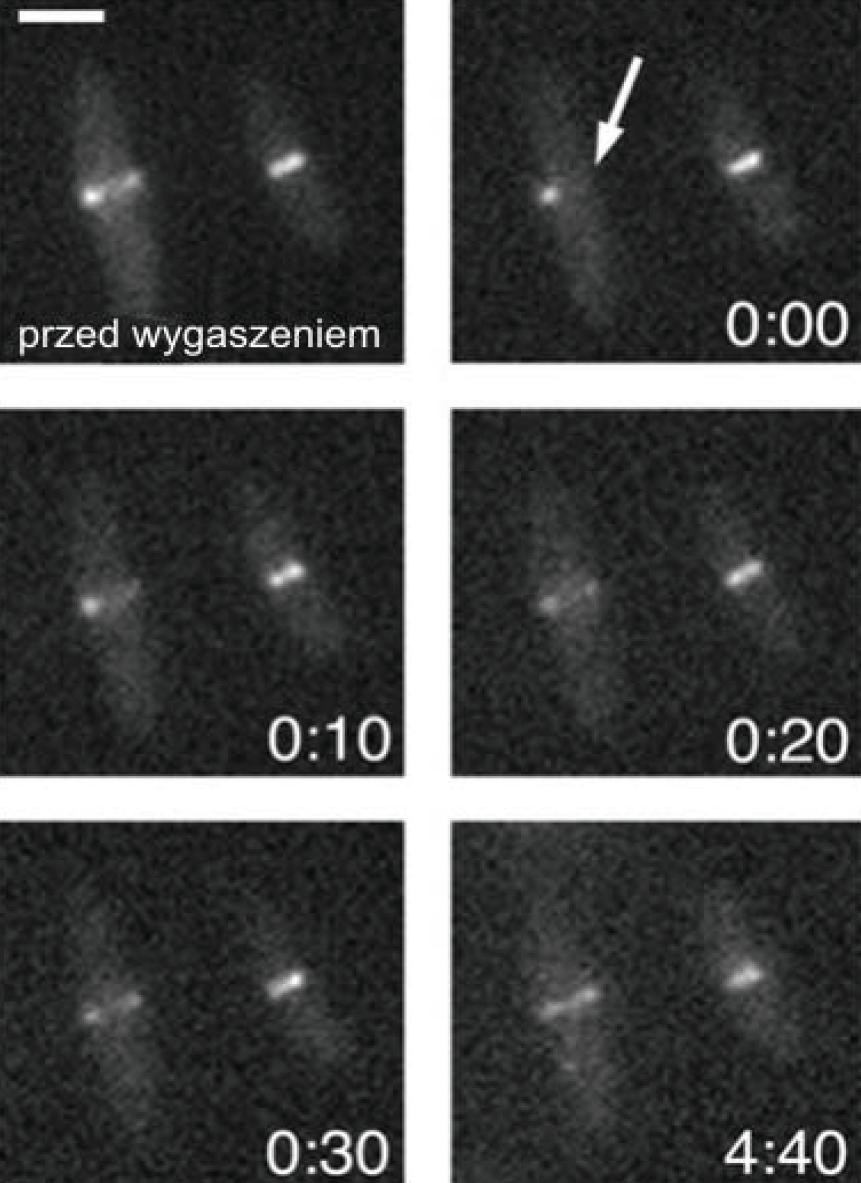
Ryc. 6. Przykład techniki FRAP wykorzystanej do obserwacji pierścieni FtsZ w komórkach E. coli [56, za zgodą]. Strzałką oznaczono fragment pierścienia Z, w którym fluorescencja została wygaszona. Dla każdego zdjęcia podano czas, jaki minął po wygaszeniu fluorescencji; skala: 2 µm
Badania z wykorzystaniem techniki FRAP pozwoliły także na analizę dynamicznego zachowania cząsteczek MreB w komórkach B. subtilis (ryc. 7); przywrócenie fluorescencji po wygaszeniu znakowanych GFP filamentów MreB następuje już po 2 min, co oznacza, że białko to tworzy struktury charakteryzujące się stosunkowo dużą dynamiką [16].

Ryc. 7. Analiza ruchu białka GFP-MreB w komórkach B. subtilis wykorzystująca technikę FRAP [16, za zgodą]. Obrazy mikroskopowe obserwowane: Pre – przed wygaszeniu laserem; 0 – w chwili wygaszenia; 1, 2, 3, 4, 5 – kolejno po wygaszeniu w odstępach 1-minutowych. Przerywanym kółkiem oznaczono region, w którym wygaszono fluorescencję; strzałkami miejsce w komórce po wygaszeniu fluorescencji; trójkątami dynamiczną lokalizację filamentów MreB-GFP; kreskami oznaczono końce komórek lub septy pomiędzy komórkami; skala: 2 µm
Znaczniki fluorescencyjne wbadaniu oddziaływań białek
Obecnie do najważniejszych metod wykorzystujących znaczniki fluorescencyjne w celu obserwacji oddziaływań białkowych w komórce w czasie rzeczywistym należą BiFC i FRET.
Pierwsza z nich, BiFC (bimolecular fluorescence complementation) jest techniką opartą na rekonstrukcji znacznika fluorescencyjnego wewnątrz komórki. Każde z badanych (potencjalnie oddziałujących ze sobą) białek (lub peptydów) podlega ekspresji w fuzji z połową funkcjonalnego białka fluorescencyjnego. Rekonstrukcja znacznika (najczęściej jest nim YFP lub GFP) nie zachodzi spontanicznie w komórce, lecz jest następstwem oddziaływania między białkami (lub peptydami), które są połączone z danym fragmentem chromoforu. Co więcej, pomiar intensywności fluorescencji technikami mikroskopii fluorescencyjnej pozwala na oszacowanie siły oddziaływań między białkami.
Technika BiFC stanowiła podstawę badań, których celem było określenie oddziaływań między białkami MreB i MbI w żywych komórkach B. subtilis [16]. Obserwacje prowadzono na komórkach szczepów kodujących kombinacje dwóch białek fuzyjnych: YN-MreB i YC-MbI. W przypadku szczepów wytwarząjących pojedyncze białka YN-MreB (ryc. 8A), YC-MbI (ryc. 8B) oraz YC-MbI i sam fragment YN (ryc. 8C) widoczne było jedynie tło fluorescencyjne. W obrazie mikroskopowym komórek wytwarzających jednocześnie białka YN-MreB oraz YC-MbI (ryc. 8D) można było dostrzec wyraźne sygnały fluorescencyjne w wielu miejscach wzdłuż ściany komórkowej. Badania te potwierdziły istnienie oddziaływań między analizowanymi białkami.
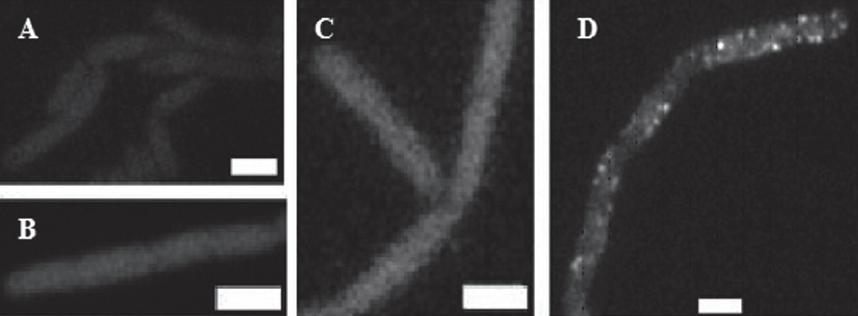
Ryc. 8. Analiza oddziaływań międzybiałkowych wykorzystująca technikę BiFC [16, za zgodą]. Szczepy B. subtilis wytwarzające białka: A – YN-MreB, B – YC-MbI, C – YC-MbI i YN, D – YN-MreB oraz YC-MbI; skala: 2 µm
FRET (Förster/fluorescence resonance energy transfer) podobnie jak BiFC jest techniką wykorzystującą znakowane fluorescencyjnie białka. Opiera się na zastosowaniu zjawiska przenoszenia energii między dwoma chromoforami będącymi w odległości nie większej niż 10 nm. Jeden z chromoforów, będący w stanie wzbudzonym (chromofor donorowy), przekazuje energię rezonansu drugiemu chromoforowi (chromofor akceptorowy). Wzbudzenie chromoforu będącego donorem następuje po naświetleniu go falą o długości odpowiadającej maksimum jego absorpcji. Potwierdzeniem istnienia oddziaływań międzybiałkowych jest emisja fali o długości odpowiadającej maksimum emisji chromoforu stanowiącego akceptor. Technika FRET została wykorzystana do analizy procesu chemotaksji w komórkach E. coli [58]. Szczególną uwagę skupiono na dwóch białkach o aktywności fosfatazy cytoplazmatycznej, CheY i CheZ oraz kinazie CheA, która wchodzi w skład kompleksu receptorowego znajdującego się wewnątrz błony komórkowej na jednym z biegunów komórek E. coli. Skonstruowano szczep, w którym jednocześnie poddano ekspresji białka CheZ i CheY w fuzji z odpowiednio CFP i YFP. Na podstawie otrzymanych wyników dowiedziono oddziaływania białek CheY i CheZ oraz możliwe stało się także określenie przestrzennego rozmieszczenia kompleksów oddziałujących białek wewnątrz komórki.
Znaczniki fluorescencyjne w badaniu dynamiki chromosomu
Zastosowanie białek w fuzji z GFP pozwoliło także na badanie dynamiki chromosomów w trakcie cyklu komórkowego. Czasoprzestrzenna organizacja replikowanego chromosomu opisywana jest głównie poprzez lokalizację i analizę dynamiki swoistych chromosomalnych loci oraz analizę globalnej topologii nowo syntetyzowanego DNA. Lokalizacja poszczególnych regionów chromosomów jest możliwa dzięki zastosowaniu systemu FROS (fluorescent repressor-operator system), który polega na wprowadzeniu w różne regiony chromosomu (np. w okolice regionu oriC – inicjującego replikację) kasety swoiście wiązanej przez białko połączone z fluorescencyjnym znacznikiem.
Jako kasety wykorzystuje się powtórzone (kilkasetkrotnie) sekwencje operatorowe tetR i/lub lacO. Ekspresja w komórce białek represorowych TerR i/lub LacI w fuzji z białkami fluorescencyjnymi (np. TetR-YFP oraz LacI-CFP), pozwala na wizualizację kompleksu nukleoproteinowego utworzonego w miejscu wprowadzenia kasety. FROS, w przeciwieństwie do wcześniej stosowanej techniki FISH (fluorescence in situ hybridization, lokalizacja określonych miejsc na chromosomie za pomocą hybrydyzacji ze znakowanymi sondami), umożliwia badania przyżyciowe. System ten wykorzystano m.in. do lokalizacji kilkudziesięciu różnych loci chromosomalnych podczas cyklu komórkowego C. crescentus wprowadzając za pomocą transpozonów kasety w różne miejsca chromosomu. Badania te wykazały istnienie mechanizmu, który warunkuje uporządkowany transport kolejnych nowo zreplikowanych fragmentów chromosomu z miejsca replikacji w stronę przeciwległego bieguna komórki [59]. W przypadku V. cholerae, Gram-ujemnych pałeczek, które mają dwa koliste chromosomy (I i II) system FROS umożliwił jednocześnie śledzenie lokalizacji regionów oriC obu chromosomów (oriCI i oriCII) w trakcie procesu replikacji. Analiza mikroskopowa pozwoliła stwierdzić, że powielone regiony oriCI są umiejscowione najczęściej w pobliżu biegunów komórki, podczas gdy nowo zreplikowane regiony oriCII są umiejscowione w pozycji 1/4 i 3/4 komórki (ryc. 9). Proces przemieszczania się regionów oriC jest skoordynowany z cyklem komórkowym V. cholerae. Zaproponowano ciekawy mechanizm rozdziału nowo zreplikowanych regionów oriC – oriCI ulega niesymetrycznej segregacji z pozycji polarnej, z jednym oriCI pozostającym na biegunie i drugim, który ulega przemieszczeniu na przeciwległy biegun komórki, natomiast oriCII podlega symetrycznej segregacji z centrum komórki do pozycji 1/4 i 3/4 [20].
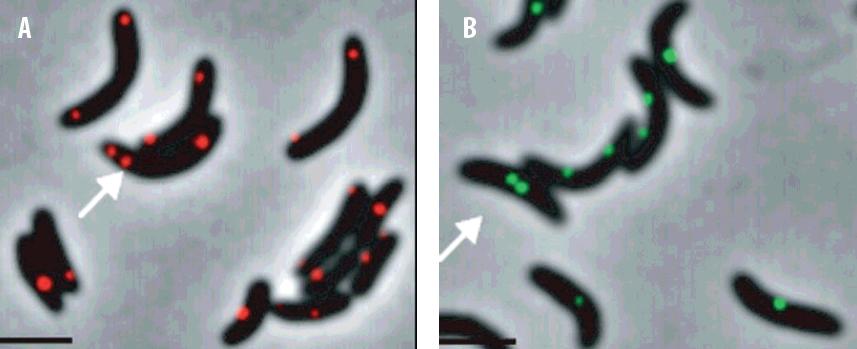
Ryc. 9. Wizualizacja regionów oriCI (A) i oriCII (B) w komórkach Vibrio cholerae systemem FROS (fluorescent repressor-operator system) [20]. A – fluorescencja pochodząca od LacI-CFP, swoiście wiązanego do sekwencji operatorowych lacO znajdujących się przy oriCIVC, B – fluorescencja pochodząca od TetR-CFP, swoiście wiązanego do sekwencji operatorowych tetO znajdujących się przy oriCIIVC, Strzałkami oznaczono komórki, w których dwa nowo zreplikowane regiony oriC znajdują się blisko siebie; skala: 2 µm
Zastosowanie białek fluorescencyjnych w badaniach ekspresji genów
Badania przeprowadzone przez Chalfiego i wsp. po raz pierwszy ukazały użyteczność GFP w analizie aktywności promotorów [11]. Zaletą stosowania gfp jako genu reporterowego jest łatwość detekcji tego białka i fluorescencja niezależna od kofaktorów. Zastosowanie białek fluorescencyjnych może też pozwolić na jednoczesne badanie kilku promotorów przez zastosowanie wariantów białka emitującego w różnych długościach fali. Problemem podczas oznaczeń ilościowych zwłaszcza w badaniach prowadzonych in situ, gdzie nie jest możliwa ścisła kontrola warunków eksperymentu jest długi czas fałdowania GFP i tworzenia się chromoforu. Może to uniemożliwić pomiary aktywności promotora w czasie, ponieważ utrudnione jest określenie czasu rozpoczęcia transkrypcji.
Zastosowanie GFP w badaniach ekspresji genów bakteryjnych pozwoliło na weryfikację tez o czasowo-przestrzennej regulacji ekspresji genów bakteryjnych. Pierwsze doniesienie o wykorzystaniu GFP, jako genu reporterowego w bakteriach pojawiło się w 1995 r. Zespół Losicka [61] badając sporulację B. subtilis wykazał swoistą dla komórki macierzystej lub dla prespory aktywność promotorów zależnych odpowiednio od podjednostek sigma polimerazy RNA sigE lub sigF. Zróżnicowanie przestrzenne aktywności tych dwóch czynników sigma pozwala na różnicowanie się dwóch kompartmentów komórki (komórki macierzystej i prespory). Pod kontrolą SigE znajduje się m.in. gen cotE, którego produkt odpowiedzialny jest za opłaszczenie endospory. Wprowadzenie genu gfp pod kontrolą promotora genu cotE wykazało, że ekspresja cotE występuje w komórce macierzystej – fluorescencja pochodząca od białka GFP obserwowana była jedynie w tym kompartmencie. Gen gfp wklonowany do plazmidu pBET131 pod kontrolą promotora regulowanego przez sigF (aktywny jedynie w presporze), wykorzystano w badaniach segregacji plazmidów niskiej kopii w komórkach B. subtilis [6]. Obserwowana fluorescencja wskazywała na obecność plazmidu pBET131w presporze (ryc. 10). W ostatnich 15 latach GFP i inne białka fluorescencyjne były wykorzystywane jako geny reporterowe dla wielu różnych mikroorganizmów np. Helicobacter pylori, Borrelia burgdorferi, Streptomyces coelicolor, Anabaena do badania regulacji rozmaitych procesów, takich jak na przykład różnicowanie, wirulencja, transport czy biosynteza metabolitów wtórnych [1,10,27,57].
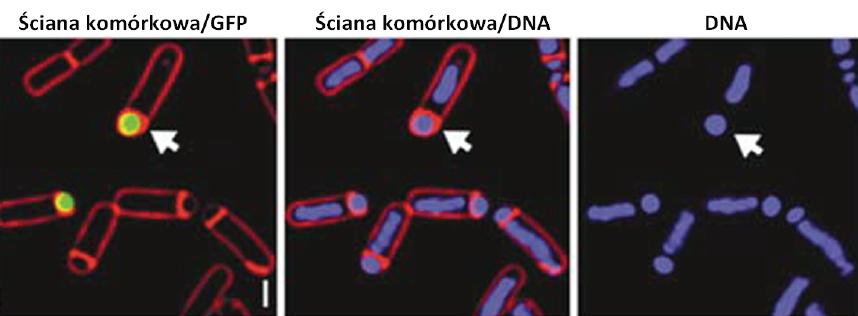
Ryc. 10. Wykorzystanie gfp jako genu reporterowego w komórkach B. subtilis w celu zbadania segregacji plazmidu pBET131 do prespory [6, za zgodą]. Strzałkami oznaczono presporę; skala: 1 µm
Podsumowanie
Badania biologii komórki rozwinęły się dzięki uhonorowanym w 2008 r. Nagrodą Nobla badaniom Osama Shimomura (odkrywca GFP), Martina Chalfie (zastosował GFP, jako fluorescencyjny znacznik) oraz Rogera Tsiena (zmodyfikował GFP otrzymując ulepszone warianty). Wprowadzenie w ostatnich kilku latach nowych, różnorodnych znaczników fluorescencyjnych oraz nowoczesnych technik obrazowania pozwala obserwować w czasie rzeczywistym umiejscowienie, przemieszczanie i działanie jednocześnie kilku białek i/lub określonych struktur komórkowych oraz regionów chromosomów wewnątrz żywej komórki. Nowe techniki wizualizacji umożliwiają również śledzenie zmian lokalizacji określonych struktur komórkowych pod wpływem różnorodnych czynników.
Wyniki badań, w których zastosowano techniki fluorescencyjne całkowicie zrewolucjonizowały nasze poglądy na temat komórki bakteryjnej. Do niedawna pokutował pogląd, że w przeciwieństwie do komórek eukariotycznych, w komórce bakteryjnej nie można wyróżnić określonych struktur subkomórkowych (odpowiedników eukariotycznych organelli) – wyobrażano sobie, że wnętrze komórki bakteryjnej to amorficzny twór złożony z różnorodnych makrocząsteczek. Z takim obrazem komórki bakteryjnej można jeszcze spotkać się w wielu podręcznikach. Obecnie wiemy, że bakterie mają cytoszkielet, który uczestniczy w aktywnej segregacji potomnych chromosomów i cytokinezie oraz utrzymywaniu określonego kształtu komórki bakteryjnej. Należy podkreślić, że oprócz omówionych tu zastosowań, techniki obrazowania są wykorzystywane w mikrobiologii lekarskiej i środowiskowej.
PIŚMIENNICTWO
[1] Aldea M.R., Mella-Herrera R.A., Golden J.W.: Sigma factor genes sigC, sigE, and sigG are upregulated in heterocysts of the cyanobacterium Anabaena sp. strain PCC 7120. J. Bacteriol., 2007; 189: 8392-8396
[PubMed] [Full Text HTML] [Full Text PDF]
[2] Anderson D.E., Gueiros-Filho F.J., Erickson H.P.: Assembly dynamics of FtsZ rings in Bacillus subtilis and Escherichia coli and effects of FtsZ-regulating proteins. J. Bacteriol., 2004; 186: 5775-5781
[PubMed] [Full Text HTML] [Full Text PDF]
[3] Arigoni F., Pogliano K., Webb C.D., Stragier P., Losick R.: Localization of protein implicated in establishment of cell type to sites of asymmetric division. Science, 1995; 270: 637-640
[PubMed]
[4] Ausmees N., Kuhn J.R., Jacobs-Wagner C.: The bacterial cytoskeleton: an intermediate filament-like function in cell shape. Cell, 2003; 115: 705-713
[PubMed]
[5] Barák I., Behari J., Olmedo G., Guzmán P., Brown D.P., Castro E., Walker D., Westpheling J., Youngman P.: Structure and function of the Bacillus SpoIIE protein and its localization to sites of sporulation septum assembly. Mol. Microbiol., 1996; 19: 1047-1060
[PubMed]
[6] Becker E., Herrera N.C., Gunderson F.Q., Derman A.I., Dance A.L., Sims J., Larsen R.A., Pogliano J.: DNA segregation by the bacterial actin AlfA during Bacillus subtilis growth and development. EMBO J., 2006; 25: 5919-5931
[PubMed] [Full Text HTML] [Full Text PDF]
[7] Bi E.F., Lutkenhaus J.: FtsZ ring structure associated with division in Escherichia coli. Nature, 1991; 354: 161-164
[PubMed]
[8] Brendler T., Sawitzke J., Sergueev K., Austin S.: A case for sliding SeqA tracts at anchored replication forks during Escherichia coli chromosome replication and segregation. EMBO J., 2000; 19: 6249-6258
[PubMed] [Full Text HTML] [Full Text PDF]
[9] Britton R.A., Lin D.C., Grossman A.D.: Characterization of a prokaryotic SMC protein involved in chromosome partitioning. Genes Dev., 1998; 12: 1254-1259
[PubMed] [Full Text HTML] [Full Text PDF]
[10] Carroll J.A., Stewart P.E., Rosa P., Elias A.F., Garon C.F.: An enhanced GFP reporter system to monitor gene expression in Borrelia burgdorferi. Microbiology, 2003; 149: 1819-1828
[PubMed] [Full Text HTML] [Full Text PDF]
[11] Chalfie M., Tu Y., Euskirchen G., Ward W.W., Prasher D.C.: Green fluorescent protein as a marker for gene expression. Science, 1994; 263: 802-805
[PubMed]
[12] Chen J.C., Weiss D.S., Ghigo J.M., Beckwith J.: Septal localization of FtsQ, an essential cell division protein in Escherichia coli. J. Bacteriol., 1999; 181: 521-530
[PubMed] [Full Text HTML] [Full Text PDF]
[13] Copeland M.F., Flickinger S.T., Tuson H.H., Weibel D.B.: Studying the dynamics of flagella in multicellular communities of Escherichia coli by using biarsenical dyes. Appl. Environ. Microbiol., 2010; 76: 1241-1250
[PubMed] [Full Text HTML] [Full Text PDF]
[14] Cormack B.P., Valdivia R.H., Falkow S.: FACS-optimized mutants of the green fluorescent protein (GFP). Gene, 1996; 173: 33-38
[PubMed]
[15] Cubitt A.B., Woollenweber L.A., Heim R.: Understanding structure-function relationship in the Aequorea victoria green fluorescent protein. Methods Cell Biol., 1999; 58: 19-30
[PubMed]
[16] Defeu Soufo H.J., Graumann P.L.: Dynamic localization and interaction with other Bacillus subtilis actin-like proteins are important for the function of MreB. Mol. Microbiol., 2006; 62: 1340-1356
[PubMed] [Full Text HTML] [Full Text PDF]
[17] Dye N.A., Pincus Z., Theriot J., Shapiro L., Gitai Z.: Two independent spiral structures control cell shape in Caulobacter. Proc. Natl. Acad. Sci. USA, 2005; 102: 18608-18613
[PubMed] [Full Text HTML] [Full Text PDF]
[18] Edwards D.H., Thomaides H.B., Errington J.: Promiscuous targeting of Bacillus subtilis cell division protein DivIVA to division sites in Escherichia coli and fission yeast. EMBO J., 2000; 19: 2719-2727
[PubMed] [Full Text HTML] [Full Text PDF]
[19] Fawcett P., Melnikov A., Youngman P.: The Bacillus SpoIIGA protein is targeted to sites of spore septum formation in a SpoIIE-independent manner. Mol. Microbiol., 1998; 28: 931-943
[PubMed] [Full Text HTML] [Full Text PDF]
[20] Fogel M.A., Waldor M.K.: A dynamic, mitotic-like mechanism for bacterial chromosome segregation. Genes Dev., 2006; 20: 3269-3282
[PubMed] [Full Text HTML] [Full Text PDF]
[21] Ghigo J.M., Weiss D.S., Chen J.C., Yarrow J.C., Beckwith J.: Localization of FtsL to the Escherichia coli septal ring. Mol. Microbiol., 1999; 31: 725-737
[PubMed] [Full Text HTML] [Full Text PDF]
[22] Hale C.A., de Boer P.A.: Direct binding of FtsZ to ZipA, an essential component of the septal ring structure that mediates cell division in E. coli. Cell, 1997; 88: 175-185
[PubMed]
[23] Hu Z., Lutkenhaus J.: Topological regulation of cell division in Escherichia coli involves rapid pole to pole oscillation of the division inhibitor MinC under the control of MinD and MinE. Mol. Microbiol., 1999; 34: 82-90
[PubMed]
[24] Imai Y., Ogasawara N., Ishigo-Oka D., Kadoya R., Daito T., Moriya S.: Subcellular localization of Dna-initiation proteins of Bacillus subtilis: evidence that chromosome replication begins at either edge of the nucleoids. Mol. Microbiol., 2000; 36: 1037-1048
[PubMed] [Full Text HTML] [Full Text PDF]
[25] Jacobs C., Domian I.J., Maddock J.R., Shapiro L.: Cell cycle-dependent polar localization of an essential bacterial histidine kinase that controls DNA replication and cell division. Cell, 1999; 97: 111-120
[PubMed]
[26] Jacobs C., Hung D., Shapiro L.: Dynamic localization of a cytoplasmic signal transduction response regulator controls morphogenesis during the Caulobacter cell cycle. Proc. Natl. Acad. Sci. USA, 2001; 98: 4095-4100
[PubMed] [Full Text HTML] [Full Text PDF]
[27] Josenhans C., Friedrich S., Suerbaum S.: Green fluorescent protein as a novel marker and reporter system in Helicobacter sp. FEMS Microbiol. Lett., 1998; 161: 263-273
[PubMed]
[28] Ju J., Luo T., Haldenwang W.G.: Forespore expression and processing of the SigE transcription factor in wild-type and mutant Bacillus subtilis. J. Bacteriol., 1998; 180: 1673-1681
[PubMed] [Full Text HTML] [Full Text PDF]
[29] Lemon K.P., Grossman A.D.: Localization of bacterial DNA polymerase: evidence for a factory model of replication. Science, 1998; 282: 1516-1519
[PubMed]
[30] Lewis P.J., Errington J.: Use of green fluorescent protein for detection of cell-specific gene expression and subcellular protein localization during sporulation in Bacillus subtilis. Microbiology, 1996; 142: 733-740
[PubMed] [Full Text PDF]
[31] Lin D.C., Levin P.A., Grossman A.D.: Bipolar localization of a chromosome partition protein in Bacillus subtilis. Proc. Natl. Acad. Sci. USA, 1997; 94: 4721-4726
[PubMed] [Full Text HTML] [Full Text PDF]
[32] Löwe J., Amos L.A.: Evolution of cytomotive filaments: the cytoskeleton from prokaryotes to eukaryotes. Int. J. Biochem. Cell Biol., 2009; 41: 323-329
[PubMed]
[33] Ma X., Ehrhardt D.W., Margolin W.: Colocalization of cell division proteins FtsZ and FtsA to cytoskeletal structures in living Escherichia coli cells by using green fluorescent protein. Proc. Natl. Acad. Sci. USA, 1996; 93: 12998-13003
[PubMed] [Full Text HTML] [Full Text PDF]
[34] Maddock J.R., Shapiro L.: Polar location of the chemoreceptor complex in the Escherichia coli cell. Science, 1993; 259: 1717-1723
[PubMed]
[35] Marston A.L., Errington J.: Dynamic movement of the ParA-like Soj protein of B. subtilis and its dual role in nucleoid organization and developmental regulation. Mol. Cell, 1999; 4: 673-682
[PubMed]
[36] Marston A.L., Thomaides H.B., Edwards D.H., Sharpe M.E., Errington J.: Polar localization of the MinD protein of Bacillus subtilis and its role in selection of the mid-cell division site. Genes Dev., 1998; 12: 3419-3430
[PubMed] [Full Text HTML] [Full Text PDF]
[37] Mizuno H., Sawano A., Eli P., Hama H., Miyawaki A.: Red fluorescent protein from Discosoma as a fusion tag and a partner for fluorescence resonance energy transfer. Biochemistry, 2001; 40: 2502-2510
[PubMed]
[38] Onogi T., Niki H., Yamazoe M., Hiraga S.: The assembly and migration of SeqA-Gfp fusion in living cells of Escherichia coli. Mol. Microbiol., 1999; 31: 1775-1782
[PubMed] [Full Text HTML] [Full Text PDF]
[39] Patterson G.H., Lippincott-Schwartz J.: A photoactivatable GFP for selective photolabeling of proteins and cells. Science, 2002; 297: 1873-1877
[PubMed]
[40] Phillips G.J.: Green fluorescent protein – a bright idea for the study of bacterial protein localization. FEMS Microbiol. Lett., 2001; 204: 9-18
[PubMed]
[41] Pogliano J.: The bacterial cytoskeleton. Curr. Opin. Cell Biol., 2008; 20: 19-27
[PubMed]
[42] Price K.D., Losick R.: A four-dimensional view of assembly of a morphogenetic protein during sporulation in Bacillus subtilis. J. Bacteriol., 1999; 181: 781-790
[PubMed] [Full Text HTML] [Full Text PDF]
[43] Ptacin J.L., Nollmann M., Becker E.C., Cozzarelli N.R., Pogliano K., Bustamante C.: Sequence-directed DNA export guides chromosome translocation during sporulation in Bacillus subtilis. Nat. Struct. Mol. Biol., 2008; 15: 485-493
[PubMed] [Full Text PDF]
[44] Quisel J.D., Lin D.C., Grossman A.D.: Control of development by altered localization of a transcription factor in B. subtilis. Mol. Cell, 1999; 4: 665-672
[PubMed]
[45] Rajagopalan M., Maloney E., Dziadek J., Poplawska M., Lofton H., Chauhan A., Madiraju M.V.: Genetic evidence that mycobacterial FtsZ and FtsW proteins interact, and colocalize to the division site in Mycobacterium smegmatis. FEMS Microbiol. Lett., 2005; 250: 9-17
[PubMed]
[46] Raskin D.M., de Boer P.A.: The MinE ring: an FtsZ-independent cell structure required for selection of the correct division site in E. coli. Cell, 1997; 91: 685-694
[PubMed]
[47] Raskin D.M., de Boer P.A.: Rapid pole-to-pole oscillation of a protein required for directing division to the middle of Escherichia coli. Proc. Natl. Acad. Sci. USA, 1999; 96: 4971-4976
[PubMed] [Full Text HTML] [Full Text PDF]
[48] Shaner N.C., Campbell R.E., Steinbach P.A., Giepmans B.N., Palmer A.E., Tsien R.Y.: Improved monomeric red, orange and yellow fluorescent proteins derived from Discosoma sp. red fluorescent protein. Nat. Biotechnol., 2004; 22: 1567-1572
[PubMed]
[49] Shaner N.C., Patterson G.H., Davidson M.W.: Advances in fluorescent protein technology. J. Cell Sci., 2007; 120: 4247-4260
[PubMed] [Full Text HTML] [Full Text PDF]
[50] Shapiro L., McAdams H.H., Losick R.: Why and how bacteria localize proteins. Science, 2009; 326: 1225-1228
[PubMed]
[51] Shih Y.L., Fu X., King G.F., Le T., Rothfield L.: Division site placement in E. coli: mutations that prevent formation of the MinE ring lead to loss of the normal midcell arrest of growth of polar MinD membrane domains. EMBO J., 2002; 21: 3347-3357
[PubMed] [Full Text HTML] [Full Text PDF]
[52] Shih Y.L., Le T., Rothfield L.: Division site selection in Escherichia coli involves dynamic redistribution of Min proteins within coiled structures that extend between the two cell poles. Proc. Natl. Acad. Sci. USA, 2003; 100: 7865-7870
[PubMed] [Full Text HTML] [Full Text PDF]
[53] Shimomura O., Johnson F.H., Saiga Y.: Extraction, purification and properties of aequorin, a bioluminescent protein from luminous hydromedusan, Aequorea. J. Cell. Comp. Physiol., 1962; 59: 223-239
[PubMed]
[54] Sievers J., Errington J.: The Bacillus subtilis cell division protein FtsL localizes to sites of septation and interacts with DivIC. Mol. Microbiol., 2000; 36: 846-855
[PubMed] [Full Text HTML] [Full Text PDF]
[55] Sourjik V., Berg H.C.: Localization of components of the chemotaxis machinery of Escherichia coli using fluorescent protein fusions. Mol. Microbiol., 2000; 37: 740-751
[PubMed] [Full Text HTML] [Full Text PDF]
[56] Stricker J., Maddox P., Salmon E.D., Erickson H.P.: Rapid assembly dynamics of the Escherichia coli FtsZ-ring demonstrated by fluorescence recovery after photobleaching. Proc. Natl. Acad. Sci. USA, 2002; 99: 3171-3175
[PubMed] [Full Text HTML] [Full Text PDF]
[57] Sun J., Kelemen G.H., Fernández-Abalos J.M., Bibb M.J.: Green fluorescent protein as a reporter for spatial and temporal gene expression in Streptomyces coelicolor A3(2). Microbiology, 1999; 145: 2221-2227
[PubMed] [Full Text HTML] [Full Text PDF]
[58] Vaknin A., Berg H.C.: Single-cell FRET imaging of phosphatase activity in the Escherichia coli chemotaxis system. Proc. Natl. Acad. Sci. USA, 2004; 101: 17072-17077
[PubMed] [Full Text HTML] [Full Text PDF]
[59] Viollier P.H., Thanbichler M., McGrath P.T., West L., Meewan M., McAdams H.H., Shapiro L.: Rapid and sequential movement of individual chromosomal loci to specific subcellular locations during bacterial DNA replication. Proc. Natl. Acad. Sci. USA, 2004; 101: 9257-9262
[PubMed] [Full Text HTML] [Full Text PDF]
[60] Waidner B., Specht M., Dempwolff F., Haeberer K., Schaetzle S., Speth V., Kist M., Graumann P.L.: A novel system of cytoskeletal elements in the human pathogen Helicobacter pylori. PLoS Pathog., 2009; 5: e1000669
[PubMed] [Full Text HTML] [Full Text PDF]
[61] Webb C.D., Decatur A., Teleman A., Losick R.: Use of green fluorescent protein for visualization of cell-specific gene expression and subcellular protein localization during sporulation in Bacillus subtilis. J. Bacteriol., 1995; 177: 5906-5911
[PubMed] [Full Text PDF]
[62] Wehrl W., Niederweis M., Schumann W.: The FtsH protein accumulates at the septum of Bacillus subtilis during cell division and sporulation. J. Bacteriol., 2000; 182: 3870-3873
[PubMed] [Full Text HTML] [Full Text PDF]
[63] Weiss D.S., Chen J.C., Ghigo J.M., Boyd D., Beckwith J.: Localization of FtsI (PBP3) to the septal ring requires its membrane anchor, the Z ring, FtsA, FtsQ, and FtsL. J. Bacteriol., 1999; 181: 508-520
[PubMed] [Full Text HTML] [Full Text PDF]
[64] Wery M., Woldringh C.L., Rouviere-Yaniv J.: HU-GFP and DAPI co-localize on the Escherichia coli nucleoid. Biochimie, 2001; 83: 193-200
[PubMed]
[65] Wheeler R.T., Shapiro L.: Differential localization of two histidine kinases controlling bacterial cell differentiation. Mol. Cell, 1999; 4: 683-694
[PubMed]
[66] Yamaichi Y., Duigou S., Shakhnovich E.A., Waldor M.K.: Targeting the replication initiator of the second Vibrio chromosome: towards generation of vibrionaceae-specific antimicrobial agents. PLoS Pathog., 2009; 5: e1000663
[PubMed]
[67] Yu X.C., Tran A.H., Sun Q., Margolin W.: Localization of cell division protein FtsK to the Escherichia coli septum and identification of a potential N-terminal targeting domain. J. Bacteriol., 1998; 180: 1296-1304
[PubMed]
Autorki deklarują brak potencjalnych konfliktów interesów.