
Charakterystyka subpopulacji limfocytów T
Paulina Niedźwiedzka-Rystwej 1 , Beata Tokarz-Deptuła 1 , Wiesław Deptuła 2Streszczenie
W pracy omówiono subpopulacje limfocytów T, to jest komórek pomocniczych, cytotoksycznych, regulatorowych, pamięci i innych oraz odmian tych limfocytów. Obecnie wśród komórek T-pomocniczych wyodrębnia się: limfocyty Th0, Th1, Th2, Th9, Th17, Th22, TFH oraz nTh2, wśród limfocytów T-cytotoksycznych komórki Tc, a ze względu m.in. na pełnione funkcje należy zaliczyć komórki NKT, Tγδ i T CD8αα (IEL), w tym nIEL i iIEL, limfocytów T-regulatorowych: limfocyty nTreg, iTreg, TR1, iTR35, limfocyty T CD8, w tym CD8+CD122+ i CD8+CD28- oraz T CD11c+CD8+. Natomiast wśród limfocytów T-pamięci: komórki Tcm oraz Tem, a wśród limfocytów T innych wyodrębnia się komórki: Tn, w tym T αβ CD4+ i T αβ CD8+ oraz limfocyty T wykończone/zmęczone i anergiczne.
Słowa kluczowe:limfocyty T • subpopulacje
Summary
The paper describes the characteristics, receptor profile and functions of T lymphocyte subpopulations (helper, cytotoxic, regulatory, memory and others). Among T helper cells one can enumerate Th0, Th1, Th2, Th9, Th17, Th22, TFH and nTh2, while T cytotoxic cells include Tc, NKT, Tγδ, and T CD8αα (IEL). Among regulatory cells there are nTreg, iTreg, TR1, and iTR35, as well as T lymphocytes with CD8, such as CD8+CD122+, CD8+CD28-, and CD11c+CD8+. And among memory T cells there are Tcm and Tem. Moreover, there are some so-called other T cells, such as Tn (T αβ CD4+ and T αβ CD8+), T exhausted and T anergic.
Key words:T lymphocytes • subpopulations
Wstęp
Dotąd podstawą klasyfikacji limfocytów T było występowanie na ich powierzchni receptorów TCR, antygenów zgodności tkankowej klasy I (MHC I) oraz receptorów różnicowania – CD [18]. Na tej podstawie wyodrębniono limfocyty pomocnicze (Th) o receptorze CD4, cytotoksyczne (Tc) o receptorze CD8 oraz regulatorowe (Treg) o niejednoznacznym garniturze receptorowym – od znacznika CD4, poprzez CD8 i CD25 aż do receptora FOXP3, a także limfocyty pamięci (Tm) o receptorze CD3 oraz inne limfocyty T, a także odmiany limfocytów T, np. komórki CIK o receptorze CD3 (tab. 1, 2). Ponadto, w zależności od różnic w budowie receptora TCR, wyróżniono wśród tych komórek, limfocyty Tαb i Tγd (tab. 1). Obecny podział limfocytów T oparto nie tylko o receptory powierzchniowe, ale głównie o wydzielane przez nie substancje i pełnione przez nie funkcje, co doprowadziło do wyodrębnienia wśród limfocytów T, komórek pomocniczych, cytotoksycznych, regulatorowych, pamięci oraz innych. Podział ten nie uwzględnił niektórych dotychczas opisywanych subpopulacji limfocytów T oraz ich odmian (tab.1).
Tabela 1. Subpopulacje limfocytów T i ich odmiany
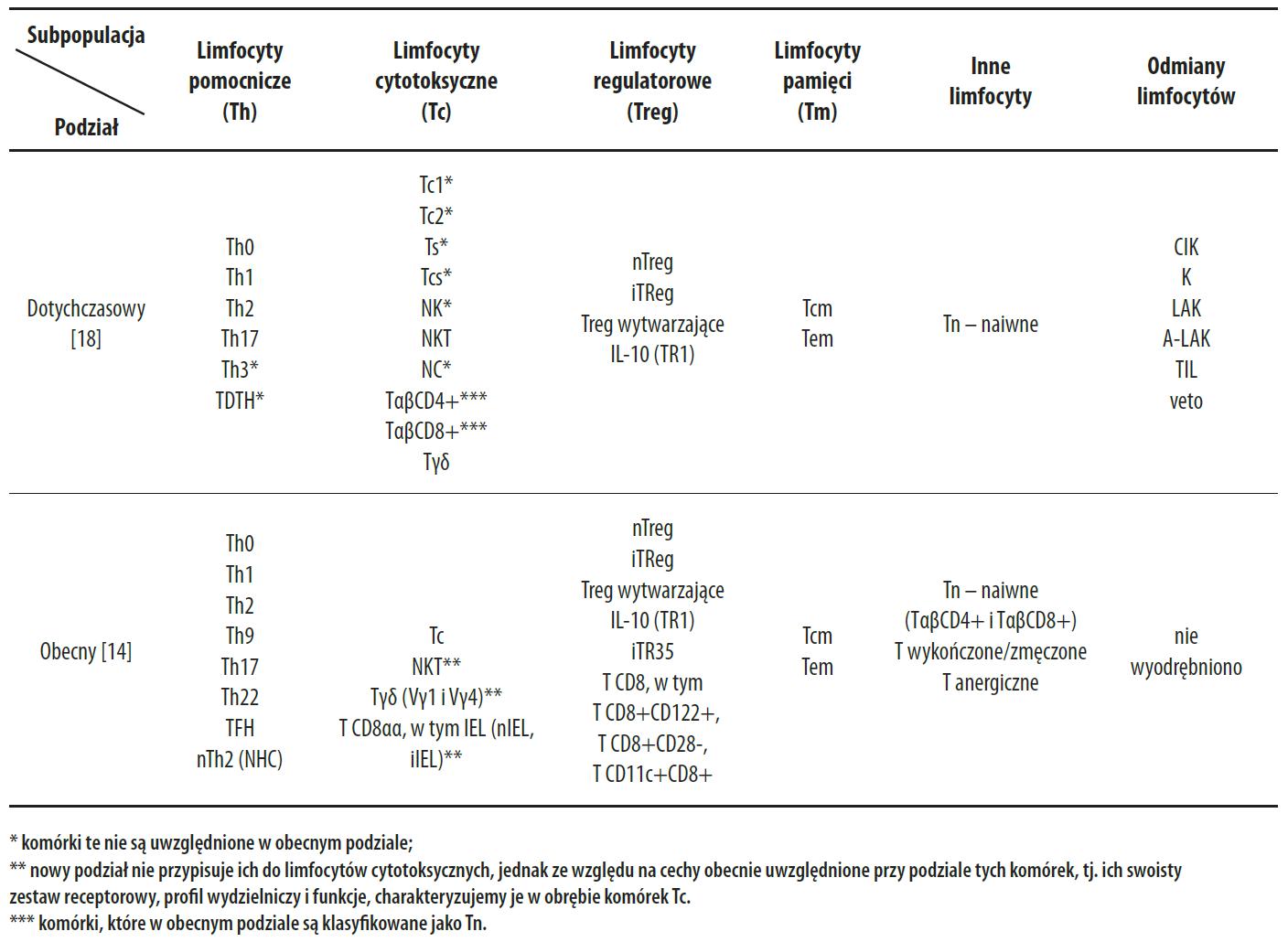
Tabela 2. Rola subpopulacji limfocytów T

Limfocyty pomocnicze (Th)
W dotychczasowym podziale limfocytów Th, wyróżniano komórki Th0, Th1, Th2, Th3, Th17 i TDTH (tab.1). Obecnie wśród nich wyodrębniono limfocyty Th0, Th1, Th2, Th9, Th17, Th22, TFH i nTh2, lecz nie wymienia się limfocytów Th3 i TDTH (tab.1).
Omawiając limfocyty pomocnicze Th0, trzeba stwierdzić, że mają one receptor CD4, wydzielają IL-2, -3, -4, -5, -10, IFN-γ, GM-CSF i w wyniku stymulacji mogą się przekształcić w dowolny typ komórek Th [18]. Natomiast limfocyty Th1 i Th2 mają receptor CD4 i CD3, z tym że komórki Th1 wydzielając IL-2 i IFN-γ uczestniczą w odpowiedzi typu komórkowego przeciwko patogenom wewnątrzkomórkowym. To one przede wszystkim wpływają na utrzymanie równowagi między limfocytami Treg, a Th17 [27] i łatwiej „wchodzą” w proces apoptozy niż komórki Th17 [64]. Natomiast limfocyty Th2 wydzielając IL-4, -5, -10, -13, są podstawowymi komórkami warunkującymi mechanizmy odpowiedzi humoralnej przeciwko zewnątrzkomórkowym patogenom (tab. 2). Biorą one udział w infekcjach przeciwpasożytniczych oraz chorobach alergicznych, np. astmie [28,67]. Wykazano także, że limfopoetyna stromalna grasicy (TSLP – thymic stromal lymphopoietin), IL-25 oraz IL-33, aktywując komórki nabłonkowe, wzmacniają także odpowiedź mediowaną przez limfocyty Th2 [28,60]. Dowiedziono również [52], że w różnicowaniu komórek Th1 i Th2, istotną rolę odgrywa system dopełniacza, który jak dotychczas był uważany za główny element w funkcjonowaniu limfocytów B. W przypadku komórek Th9 (tab.1) wykazano, że zawierają na swojej powierzchni receptor CD3 i CD4, wydzielają IL-9, -10, TGF-β i biorą udział w reakcjach alergicznych oraz w odporności przeciwko zewnątrzkomórkowym patogenom – głównie nicieniom [14,47]. Komórki te, wydzielając IL-25, oddziałują autokrynnie podczas reakcji alergicznych. Podobnie jak limfocyty Th2, komórki Th9 wydzielają TGF-β i ich działanie jest sprzężone, gdyż syntetyzowana przez nie interleukina 9, aktywuje komórki Th2 [44,47]. Natomiast limfocyty Th17 mają receptor CD4 oraz CD3, wydzielają IL-17, IL- 6, TNF i GM-CSF, stąd określane są często jako limfocyty T-regulatorowe [46]. Ważny dla ich funkcjonowania jest receptor jądrowy RORα i RORγt (retinoid-related orphan receptor), gdyż jego brak hamuje wytwarzania IL- 17, powodując tym samym defekt rozwoju komórek Th17 [33,54]. Limfocyty te wykazują działanie przeciwko różnym patogenom, w tym bakteryjnym, np. Klebsiella pneumoniae i Citrobacter rodentium, a także grzybiczym, takim jak Candida albicans, zwłaszcza w obrębie błon śluzowych [22,49]. Ich rolę wykazano także w ośrodkowym układzie nerwowym w eksperymentalnym zakażeniu mózgu i rdzenia kręgowego u myszy [34] oraz w chorobach alergicznych i autoimmunologicznych, a także w stanach zapalnych oraz w kształtowaniu odporności przeciwnowotworowej, głównie w raku jajnika [22,56]. Komórki Th17 wytwarzają IL-21 i czynnik TGF-β, zamiast IL-6, których immunosupresyjne działanie wpływa na kształtowanie odpowiedzi przeciwzapalnej [22,70]. Jest to szczególnie widoczne w reaktywności makrofagów aktywowanych limfocytami Th17 [30]. Czynnikiem regulującym różnicowanie limfocytów Th17 jest ścieżka kinazy mTor [13], a ich aktywność kształtowana jest głównie przez czynnik IRF5 (interferon-regulatory factor), który aktywuje w nich geny kodujące IFN, TNF, IL-12 i IL-23 [30]. Wykazano, że mimo iż rola IL-23 w dojrzewaniu i różnicowaniu komórek Th17 jest dyskusyjna, cytokina ta wpływa na zwiększenie liczby dojrzałych komórek Th17, przez co oddziałuje na kształtowanie odporności [22]. Dowiedziono także [35], że IL-2 działa supresyjnie na rozwój i różnicowanie komórek Th17, a IL-27 wpływa hamująco na ich działanie [54]. W przypadku limfocytów Th22 zarejestrowano na ich powierzchni receptor CD3, CD4 i CCR10 i wykazano, że są podobne do limfocytów Th17, bo mają receptor CCR6 i CCR4, choć brak u nich typowego dla tych ostatnich limfocytów, receptora CD161 [66]. Do prawidłowego funkcjonowania tych komórek konieczne jest występowanie czynnika transkrypcyjnego AHR (aryl hydrocarbon receptor), który ma zdolność interakcji z wieloma ligandami, w tym z toksynami środowiskowymi [65]. Limfocyty Th22 wydzielają IL-22 i biorą udział w chorobach skóry, np. w łuszczycy [14,66]. Inną subpopulacją wśród limfocytów T pomocniczych są limfocyty folikularne – TFH, zawierające na swej powierzchni wiele receptorów, w tym m.in: CD3, CD4, CXCR5, SLAM, OX40L, CD40L, ICOS, IL-21R, PD1 i BTLA [10,11]. Wyizolowano je z ludzkich migdałków, jako subpopulację limfocytów T z receptorem CD4+ [11]. Wydzielają one IL-21 i ich funkcja związana jest z aktywnością limfocytów B, przez co bardzo często są łączone z limfocytami T-regulatorowymi [10,11,14,46]. Czynnikiem transkrypcyjnym wzmagającym ich aktywność jest Bcl-6 (Bcl-2-oncogene B cell leukemia) [70], zaś czynnikiem definiującym jest sekrecja czynnika CXCR5 i PD1 (programmed death 1) [71]. Również te ostatnie substancje warunkują komórki folikularne – GC i non-GC, czyli związane i niezwiązane z centroblastami węzłów chłonnych. Szczególną cechą tych komórek jest występowanie w pęcherzykach pierwotnych w węzłach chłonnych będących skupiskiem limfocytów B (B cell follicles) [11]. Inną subpopulacją limfocytów Th są niedawno opisane komórki nTh2, nazywane pierwotnie NHC (natural helper cell) [41,42,43,63], które występują w skupiskach limfoidalnych, związanych z tkanką tłuszczową FALC (FAT-associated lymphoid clusters), zwłaszcza okolic nerek i narządów płciowych [29]. Zawierają one charakterystyczne dla progenitorowych komórek NK receptor c-Kit (stem-cell receptor) oraz Sca-1 (stem cell antygen -1), ale nie wykazują typowego dla nich receptora TCR [43,63].
Wytwarzają IL-4, -5, -6, -7, -10 i -13 i ich rola związana jest z lokalną odpornością błon śluzowych przewodu pokarmowego (tab. 2), choć także z udziałem w odporności przeciwpasożytniczej u myszy, wywołanej Nippostrongylus brasiliensis [29,41,42,43,63]. Wpływają na proliferację limfocytów B w kępkach Peyera, a także stymulują komórki kubkowe do wydzielania śluzu [41,42,43,63]. Wydzielana przez te komórki IL-5 wzmaga proliferację limfocytów B1 [43]. Charakteryzując te komórki, należy wspomnieć o innych nowo opisanych komórkach układu odpornościowego, które podobnie jak nTh2, biorą udział w polaryzacji odpowiedzi immunologicznej na korzyść odpowiedzi mediowanej limfocytami Th2. Komórki te to nuocyty, wielopotencjalne komórki pregenitorowe typu 2 (MMPtype2) i wrodzone komórki pomocnicze typu 2 (Ih2) [45,58,59]. Nuocyty to komórki wydzielające IL-13 i w mniejszym stopniu IL-5 i IL-4, a ich rola łączy się głównie z obroną organizmu przed infekcjami pasożytniczymi [28,45,58], zaś wielopotencjalne komórki progenitorowe MMPtype2, jako jedyne z tych subpopulacji, mają zdolność przekształcania się w inne komórki UO, ale ich funkcja to głównie rola przeciwpasożytnicza [58,59]. Podobną funkcją cechują się wrodzone komórki pomocnicze typu 2 (Ih2) mające na swej powierzchni markery CD44 i CD90.2 oraz wydzielające IL-25 i IL-33 [59].
Omawiając grupę limfocytów T-pomocniczych, należy wspomnieć o limfocytach Th3 oraz komórkach TDTH, których w obecnym podziale nie uwzględnia się (tab.1). Limfocyty Th3 – zwane także Tr3, mają receptor CD4, wytwarzają duże ilości TGF-β oraz nieduże ilości IL-4 i -10, które są wykorzystywane do ich wzrostu i różnicowania. Ich rola wiąże się z utrzymywaniem tolerancji pokarmowej, ponieważ poprzez wytwarzanie czynnika TGF-β, oddziałują na funkcjonowanie limfocytów nTreg [38]. Natomiast limfocyty TDTH, to komórki T z receptorem CD4, biorące udział w nadwrażliwości typu późnego, np. w przypadku zakażenia ssaków prątkiem gruźlicy, a w stosunku do których, nie określono ich swoistego profilu wydzielniczego (17).
Limfocyty cytotoksyczne
Wśród limfocytów T-cytotoksycznych (Tc), w dotychczasowym podziale (tab.1) wyróżniono limfocyty Tc1, Tc2, Ts (supresorowe), Tcs (kontrasupresorowe), komórki NK (natural killer), limfocyty NKT i NC, a także limfocyty TαbCD8+, Tαb4+ i Tγd (tab.1). Obecny podział wśród tych limfocytów wyodrębnia jedynie limfocyty Tc, choć jak się wydaje, ze względu na garnitur receptorowy, a głównie ich profil wydzielniczy i funkcje, należy w obrębie tej grupy limfocytów zaliczyć komórki NKT oraz limfocyty Tγd i limfocyty T CD8αa (IEL), w tym nIEL i iIEL. Komórki Tc to komórki mające receptory CD8 i CD3, wydzielające perforyny, granzymy i IFN-γ i wykazujące zdolność cytotoksyczności i zabijania patogenów. Natomiast komórki NKT u myszy zawierają receptory NK1.1, SLAMF1, SLAMF6, TGF-βR, Vα14 i Jα18, a u ludzi Vα24 i Jα18 [6,14]. Syntetyzują one IL-4, IFN-γ oraz IL-17A [14] i wykazują funkcje pro- i antyzapalne oraz modulujące reakcje odpornościowe w chorobach nowotworowych, schorzeniach autoimmunologicznych, alergicznych oraz infekcjach bakteryjnych, wirusowych i pasożytniczych (tab. 2). Komórki te do prawidłowego rozwoju wymagają obecności białka E, które uważane jest za istotne w procesie dojrzewania limfocytów T [6]. Również IL-9 komórek NKT obecnych w błonie śluzowej nosogardzieli u ludzi, stanowi czynnik wzrostu [47]. W przypadku subpopulacji limfocytów Tγd, wykazano, że mają one charakterystyczny receptor TCR γd, ale nie mają, jak prawie wszystkie limfocyty T, receptora TCRαβ [14,18]. Zawierają one także receptor CD3 i syntetyzują IFN-γ, IL-17A, IL-17F oraz IL-22, a ich rola związana jest głównie z reakcjami odporności naturalnej, choć także i nabytej, głównie na powierzchniach nabłonkowych, gdzie spełniają rolę prozapalną i antyzapalną [14] (tab. 2). Komórki te odgrywają znaczącą rolę w zakażeniach pasożytniczych Nippostrongylus brasiliensis w jelicie cienkim u ssaków [23], a przez oddziaływanie na nie limfocytów nTreg, wpływają na zachowanie homeostazy immunologicznej w jelitach [50]. Natomiast opisane w obrębie limfocytów Tγd dwie grupy komórek, tj. Vγ1 i Vγ4 to komórki mało znane, choć przypisuje się im rolę w odporności przeciwnowotworowej [20]. W przypadku limfocytów T CD8αa stwierdzono, że mają one albo receptor TCR-αb lub TCR-γd, co nie zdarza się w innych subpopulacjach komórek T, a dodatkowo mają znacznik CD8αa i B220 [14]. Wydzielają IL-10 i TGF-β, a ich funkcja związana jest głównie z regulacyjnymi procesami odporności lokalnej w jelitach [14]. Przyjmuje się [14,16], że do komórek T CD8αa, należy zaliczyć także śródnabłonkowe limfocyty IEL (intraepithelial lymphocyte), występujące głównie w jelitach u ssaków i aktywowane tylko przez receptor CD2 [16], choć są dane donoszące, że limfocyty te, podobnie jak pozostałe komórki T, aktywowane są przez receptor CD3 [14]. Komórki IEL mają dodatkowo receptor CD103 (αE integrynę), który współdziała z E-kadheryną na komórkach nabłonkowych jelita [7]. Komórki IEL „współpracują” z limfocytami nTreg w zwalczaniu zarazków, które dostały się do ustroju drogą oralną, choć u ludzi udowodniono in vitro [51], że są one zdolne do zwalczania komórek nowotworowych. Wśród nich, zarówno w jelicie cienkim, jak i grubym wyodrębniono naturalne limfocyty IEL (nIEL) oraz indukowane komórki IEL (iIEL), o dużej aktywności przeciwzarazkowej i różniące się tym, że nIEL są obecne od narodzin i zanikają z wiekiem, natomiast iIEL pojawiają się dopiero w czasie życia osobniczego [7].
Charakteryzując subpopulacje limfocytów T-cytotoksycznych, należy dodać, że obecny podział nie uwzględnia takich komórek jak: Tc1, Tc2, Ts, Tcs, NK, NC, zaś limfocyty TαbCD8+ i TαβCD4+ przypisuje do limfocytów Tn. Podać należy, że komórki Tc1 i Tc2 to limfocyty o receptorze CD8, różniące się sekrecją swoistych interleukin, jako że limfocyty Tc1 wydzielają głównie IL-4, zaś Tc2 – IL-5 [18]. Natomiast limfocyty Ts mają także receptor CD8 i obecnie są włączone w szeroko pojętą populację limfocytów regulatorowych [18]. W przypadku limfocytów Tcs przyjęto, że ich swoistym receptorem jest znacznik CD3 i CD8, z tym że tak jak w przypadku limfocytów Ts nie określono ich profilu wydzielniczego, choć przyjęto, że komórki te chronią limfocyty Th przed „sygnałem” supresyjnym [37]. Natomiast komórki NK to naturalni bójcy, o receptorach CD2, CD16, CD56, wydzielające IFN-α, IL-2, IL-3, GM-CSF, CSF, TNF i biorące udział w odpowiedzi przeciwnowotworowej [18]. Komórki NC to limfocyty o silnych właściwościach cytotoksycznych i bliżej nieokreślonych receptorach, wydzielające również IL-2, biorące udział w procesach zapalnych [18]. Natomiast komórki TαbCD4+ i TαbCD8+ to limfocyty, które zostały sklasyfikowane jako limfocyty Tn.
Limfocyty regulatorowe
W obrębie subpopulacji limfocytów T regulatorowych (Treg) (tab. 1), dotychczasowy podział uwzględniał istnienie naturalnych limfocytów regulatorowych – nTreg, indukowanych – iTreg oraz Treg wytwarzających IL-10 (TR1). Obecny podział oprócz dotychczasowych trzech subpopulacji (nTreg, iTreg oraz TR1), w tej grupie komórek prezentuje także limfocyty iTR35 oraz limfocyty T CD8, w tym CD8+CD122+, CD8+CD28- i CD11c+CD8+.
Komórki nTreg to limfocyty o receptorze CD4, CD25, a także CD3, CTLA4 i GITR oraz FOXP3, wydzielające IL- 10, TGF-β i IL-35 [14,21,40] o działaniu regulatorowym, w tym supresyjnym, m.in. na odpowiedź limfocytów Th1 i Th2 w czasie powszechnie występujących infekcji dróg oddechowych u ssaków, w tym i u ludzi [12]. Limfocyty te oddziałują hamująco na limfocyty B, bo mogą indukować śmierć tych komórek oraz hamować syntezę immunoglobulin [72]. Ich rolę wykazano u ludzi dotkniętych cukrzycą typu 1 oraz u kobiet w ciąży, gdyż w związku z obecnością antygenów ojcowskich zarejestrowano ich zwiększoną liczbę we krwi doczesnej, jak i obwodowej [2]. Na prawidłowe funkcjonowanie i działanie czynnika FOXP3 stanowiącego podstawę ich funkcjonowania, wpływa ligaza PIAS1 (protein inhibitor of the activated signal transducer of transcription STAT1), która warunkuje odpowiednie funkcjonowanie promotora genu Foxp3, którego aktywacja jest zależna od czynnika Blimp-1 (B-lymphocyte-induced maturation protein 1) – czynnika obligatoryjnego dla dojrzewania i różnicowania się komórek nTreg [36,48]. Na działanie limfocytów nTreg wpływa także obecność katepsyny E [53] oraz receptorów TLR, głównie TLR2, 4, 5, 7, 8, 9 i 10 [19]. Znany jest także mechanizm immunosupresyjnej aktywności komórek nTreg, warunkowany wydzielaniem cytokin blokujących, które hamują zjawisko cytotoksyczności komórek układu immunologicznego oraz ograniczają odpowiedź immunologiczną poprzez oddziaływanie na komórki APC [5]. Natomiast iTreg mają receptory CD3, CD4, CD25, CTLA4, GITR, wydzielają FOXP3, FOXO1, FOXO3, STAT5, SMAD2, SMAD3, SMAD4, a ich rola wiąże się z indukcją zjawisk immunosupresyjnych (tab.2). Natomiast komórki Treg wytwarzające IL-10, określane są jako TR1 nie wykazują ekspresji swoistego dla limfocytów regulatorowych znacznika FOXP3, zaś za czynnik ekspresyjny typowy dla nich uznano ROG, tj. represor czynnika transkrypcyjnego GATA-3 (trans-acting T-cell-specific transcription factor) [38]. Komórki te mają także receptor CD3 i CD4, ale wydzielają tylko IL-10 i ich działanie związane jest z immunotolerancją, a także udziałem w procesach zapalnych, alergicznych oraz zakażeniach bakteryjnych i wirusowych. Zarejestrowano także [55], że interleukina IL-27, będąca czynnikiem działającym supresyjnie na limfocyty Th17, wpływa aktywizująco na komórki TR1. Natomiast syntetyzowana przez te komórki IL-10 uzyskuje najwyższą koncentrację w 24 godzinie po ich aktywacji [57]. Ich niewielka zdolność proliferacyjna wzrasta w obecności IL-2 i IL-15 [57]. Limfocyty te „współdziałają” z limfocytami nTreg, gdyż te ostatnie komórki wykazują aktywność w pierwszej fazie infekcji, zaś limfocyty TR1 we wtórnej fazie zakażenia. Te ostatnie biorą także udział w reakcjach alergicznych np. na nikiel oraz alergeny pochodzenia zwierzęcego – alergeny kotów [57]. W subpopulacji iTR35, wykazano, że są one mediowane przez IL-35, lecz nie wydzielają cytokin charakterystycznych dla limfocytów T-regulatorowych, takich jak IL-10, czy TFG-β, nie mają też znacznika Foxp3, jednakże wykazują silne właściwości supresorowe w zakażeniach i chorobach nowotworowych (tab.2). Upatruje się w nich potencjalne źródło manipulacji terapeutycznych, dotyczących aktywności komórek regulujących, co stwarza możliwość lepszego leczenia chorób nowotworowych i autoimmunologicznych [3,8,9]. Natomiast subpopulacja komórek T CD8 ma swoisty znacznik CD8 i wykazuje hamujące działanie wobec limfocytów TFH, przez co wpływa na tolerancję immunologiczną i na procesy autoimmunologiczne [26], głównie u myszy [61]. Wśród tych komórek wyróżniono limfocyty T CD8+CD122+, które oprócz charakterystycznego znacznika CD8, wykazują także ekspresję CD122 i oddziałują na homeostazę immunologiczną w chorobach autoimmunologicznych [15,39]. Ponadto wykazano [15], że ta subpopulacja limfocytów chroni myszy przed chorobami jelita grubego. Natomiast subpopulacja regulatorowych komórek T CD8+CD28-, ma jedynie znacznik CD8 i wykazuje zdolność do obniżania poziomu ekspresji znaczników CD40, CD80 na komórkach Th oraz CD86 na komórkach DC [24]. Tymczasem subpopulacja komórek regulatorowych T CD11c+CD8+, charakteryzuje się znacznikiem CD11c i CD8 i wykazuje korzystne działanie w przypadkach zapalenia stawów u ludzi, mimo że mechanizm ich działania oraz wydzielane przez nie substancje, a także ich funkcje efektorowe, są mało poznane [68]. U myszy komórki te stanowią prawie 3% wszystkich limfocytów krwi obwodowej i wykazują supresyjne oddziaływanie na limfocyty T z receptorem CD4+ [68]. Trzeba także stwierdzić, że w stosunku do limfocytów T CD8 (tab. 1), a mianowicie T CD8+CD122+, T CD8+CD28- i T CD11c+CD8+, nie opisano w piśmiennictwie substancji wydzielanych przez nie [15,24,39,68].
Limfocyty pamięci
Spośród subpopulacji limfocytów T pamięci, zarówno według dotychczasowego, jak i obecnego podziału (tab.1), wyodrębnia się centralne komórki pamięci (Tcm) oraz efektorowe komórki pamięci (Tem). Komórki Tcm zawierają na swej powierzchni receptory CCR7hi, CD44, CD62Lhi, CD3, IL-7R (CD127), wydzielają IL-2, CD40L oraz niewielkie ilości IL-4, IFN-γ i IL-17A [11], a ich rola łączy się z odpowiedzią na antygeny głównie we wtórnych narządach limfoidalnych. Natomiast komórki Tem mają na swojej powierzchni receptory CD62Llow, CD44, CD3, IL-7R (CD127), IL-15R i CCR7low [14], wytwarzają wiele cytokin zapalnych i ich najistotniejsza rola związana jest z aktywowaniem odpowiedzi immunologicznej zachodzącej w limfoidalnych narządach obwodowych. Komórki te nie przechodzą przez węzły chłonne, ale napływają do miejsc, gdzie toczy się stan zapalny [18]. Funkcjonowanie komórek Tem zależy od obecności przetrwałego w ustroju antygenu, a napływając do miejsca stanu zapalnego, oddziałują one przez wydzielanie cytokin lub działanie cytotoksyczne.
Inne limfocyty T
Według dotychczasowego podziału (tab.1) wśród limfocytów T są limfocyty T naiwne (Tn), tzw. dziewicze, które uwzględnia również najnowszy podział, a które obecnie dzielą się na dwie subpopulacje, tj. komórki T αb CD4+ z receptorem dla CD3, CD4, CCR7, CD62Lhi, IL-7R (CD127) i niepełniące funkcji wydzielniczej, ale wykazujące zdolność patrolowania kompleksów MHC II na komórkach APC [14] oraz komórki T ab CD8+, zawierające niemalże taki sam zestaw receptorowy, jak komórki T αβ CD4+, z jednym wyjątkiem: miejsce receptora CD4 zastępuje znacznik CD8 i które mają za zadanie patrolowanie kompleksów MHC I na komórkach APC [14]. Warto dodać, że komórki te w poprzednim podziale charakteryzowano w obrębie limfocytów T cytotoksycznych (tab. 1). Ponadto w obrębie innych limfocytów T, wyodrębniono limfocyty T wykończone/zmęczone oraz limfocyty T anergiczne. Limfocyty T wykończone/zmęczone mają receptory CD3, CD8, PD1, TIM3, 1B11, LAG3 i ich funkcja związana jest z odpowiedzią mediowaną przez receptor TCR [14]. Natomiast limfocyty T anergiczne to komórki z receptorem CD3 oraz BTLA (B and T lymphocyte attenuator), biorące udział w reakcjach autoimmunologicznych [14], których aktywacja poprzez receptor TCR, następuje w przypadku braku sygnałów kostymulujących [14].
Należy dodać, że dotychczas wśród limfocytów T, wyodrębniano odmiany limfocytów T, których nie uwzględnia nowy podział, a do których zaliczano komórki CIK, K, LAK, A-LAK, TIL i veto (tab.1). Przyjęto, że komórki CIK mają receptor CD3 i CD56 i wykazują, podobnie jak komórki K, dużą aktywność cytotoksyczną. Natomiast komórki LAK i A-LAK wykazują także silne właściwości cytotoksyczne, ale po stymulacji IL-2. Tymczasem komórki TIL to limfocyty T o dużym znaczeniu klinicznym, ponieważ mają właściwości infiltrujące i niszczące nowotwory, zaś komórki veto to limfocyty blokujące przejście limfocytów T w komórki efektorowe [18]. Należy stwierdzić, że w stosunku do odmian limfocytów T, tj. komórek LAK, A-LAK, TIL oraz veto, nie określono ich swoistych receptorów oraz profilu wydzielniczego.
Warto także dodać, że niemal każda z wymienionych subpopulacji limfocytów T w obrębie komórek pomocniczych, cytotoksycznych, regulatorowych, pamięci oraz innych – w tym także w obrębie ich odmian – w chwili czynnego uczestniczenia w odpowiedzi immunologicznej, w wyniku spełniania przypisanych im funkcji, stają się limfocytem efektorowym (Teff) i stąd w piśmiennictwie występuje pojęcie limfocyty T efektorowe [18]. W piśmiennictwie pojawiła się także wzmianka o komórkach ILC (innate lymphoid cells), których prekursorami mogą być komórki NK lub LTi (lymphoid tissue inducer), a które wykazują cechy komórek immunokompetentnych, jakimi są limfocyty, jednak nie można ich zakwalifikować do limfocytów T, ze względu na brak swoistych receptorów antygenowych, ani do limfocytów B, ze względu na ich niezależność od genów RAG (recombination activating gene), które są odpowiedzialne za rekombinację genów VDJ immunoglobulin [1,62,69]. Komórki te charakteryzują się ekspresją receptora sierocego Rorγt (orphan receptor), a ponadto w różnych stadiach rozwoju mogą także cechować się ekspresją znacznika CD117 i CD127 [2]. Wykazano także [1,62,69], że komórki te nie mają znaczników charakterystycznych dla komórek mieloidalnych i DC. Stwierdzono także [1,62,69], że komórki ILC wykazują ekspresję cząsteczki CD4, stąd poprzednio określano je jako LTi (lymphoid tissue inducer), ze względu na ich rolę w rozwoju węzłów chłonnych i kępek Peyera. Przyjmuje się [1,62,69], że funkcja tych komórek związana jest zarówno z odpornością naturalną, jak i nabytą, zwłaszcza w infekcjach błon śluzowych. Ostatnio zaproponowano podział komórek ILC na trzy grupy – pierwszą stanowią ILC wytwarzające IFN-γ, do drugiej można zaliczyć ILC wytwarzające IL-5 i -13 i zależne od receptorów GATA3 i ROR-α, a w trzeciej znajdują się ILC wytwarzające IL-17 i -22, zależne od receptora ROR-γt [62,69].
Najnowsze dane wskazują także na istnienie limfocytów T o odmiennym od innych komórek T fenotypie, które występują w błonie śluzowej – są to komórki MAIT (mucosa-associated inwariant T). Ich rola związana jest ze zwalczaniem infekcji bakteryjnych i grzybiczych poprzez wiązanie się ich receptora MR1 do metabolitów ryboflawinowych bakterii i grzybów [17,32].
Omawiając limfocyty T, należy dodać, że w praktyce laboratoryjnej, funkcjonuje również pojęcie „limfocyty atypowe”, które są „niezłośliwymi”, dużymi, choć zmienionymi morfologicznie komórkami T, o kształcie przypominającym muszelkę [25]. Prawdopodobnie limfocyty te powstają jako „skrzyżowanie” komórki plazmatycznej z limfocytem i wtedy określane są limfoplazmocytem lub limfoidalną komórką plazmatyczną – limfocytem plazmocytoidalnym [25]. Ich występowanie zarejestrowano u ludzi głównie w zakażeniach wirusem Epsteina-Barr, cytomegalii, różyczki, opryszczki, ospy wietrznej i świnki, a także w zakażeniach bakteryjnych, takich jak kiła, gruźlica, zakażenie pałeczką Listerii, Mycoplasma pneumoniae i Shigella [25].
PIŚMIENNICTWO
[1] Aparicio-Domingo P., Cupedo T.: Rorγt+ innate lymphoid cells in intestinal homeostasis and immunity. J. Innate Immun., 2011; 3: 577-584
[PubMed] [Full Text PDF]
[2] Battaglia M., Roncarolo M.G.: Immune intervention with T regulatory cells: past lessons and future perspectives for type 1 diabetes. Semin. Immunol., 2011; 23: 182-194
[PubMed] [Full Text HTML] [Full Text PDF]
[3] Belkaid Y., Chen W.: Regulatory ripples. Nat. Immunol., 2010; 11: 1077-1078
[PubMed] [Full Text HTML] [Full Text PDF]
[4] Bielawska-Pohl A., Duś D.: When killers become helpers – the ambivalent role of NK cells. Adv. Clin. Exp. Med., 2009; 18: 537-542
[Abstract] [Full Text PDF]
[5] Budna J., Kaczmarek M., Sikora J.: Znaczenie komórek T regulatorowych w rozwoju tolerancji na nowotwór. Post. Biol. Kom., 2011; 38: 283-295
[Abstract] [Full Text PDF]
[6] Chan A.C., Berzins S.P., Godfrey D.I.: Transcriptional regulation of lymphocyte development. Developing NKT cells need their (E) protein. Immunol. Cell Biol., 2010; 88: 507-509
[PubMed] [Full Text HTML] [Full Text PDF]
[7] Cheroutre H., Lambolez F., Mucida D.: The light and dark sides of intestinal intraepithelial lymphocytes. Nat. Rev. Immunol., 2011; 11: 445-456
[PubMed]
[8] Collison L.W., Chaturvedi V., Henderson A.L., Giacomin P.R., Guy C., Bankoti J., Finkelstein D., Forbes K., Workman C.J., Brown S.A., Rehg J.E., Jones M.L., Ni H.T., Artis D., Turk M.J., Vignali D.A.: IL-35-mediated induction of a potent regulatory T cell population. Nat. Immunol., 2010; 11: 1093-1101
[PubMed] [Full Text HTML] [Full Text PDF]
[9] Collison L.W., Workman C.J., Kuo T.T., Boyd K., Wang Y., Vignali K.M., Cross R., Sehy D., Blumberg R.S., Vignali D.A.: The inhibitory cytokine IL-35 contributes to regulatory T-cell function. Nature, 2007; 450: 566-569
[PubMed]
[10] Deenick E.K., Ma C.S.: The regulation and role of T follicular helper cells in immunity. Immunology, 2011; 134: 361-367
[PubMed] [Full Text HTML] [Full Text PDF]
[11] Deenick E.K., Ma C.S., Brink R., Tangye S.G.: Regulation of T follicular helper cell formation and function by antigen presenting cells. Curr. Opin. Immunol., 2011; 23: 111-118
[PubMed] [Full Text HTML] [Full Text PDF]
[12] Dehzad N., Bopp T., Reuter S., Klein M., Martin H., Ulges A., Stassen M., Schild H., Buhl R., Schmitt E., Taube C.: Regulatory T cells more effectively suppress Th1-induced airway inflammation compared with Th2. J. Immunol., 2011; 186: 2238-2244
[PubMed] [Full Text HTML] [Full Text PDF]
[13] Delgoffe G.M., Pollizzi K.N., Waickman A.T., Heikamp E., Meyers D.J., Horton M.R., Xiao B., Worley P.F., Powell J.D.: The kinase mTor regulates the differentiation of helper T cells through the selective activation of signaling by mTORC1 and mTORC2. Nat. Immunol., 2011; 12: 295-303
[PubMed] [Full Text HTML] [Full Text PDF]
[14] Dong C., Martinez G.J.: T cells: the usual subsets. Nat. Rev. Immunol., 2010; 11 (poster) http://www.nature.com/nri/posters/tcellsubsets/nri1009_tcellsubsets_poster.pdf
[Abstract]
[15] Endharti A.T., Okuno Y., Shi Z., Misawa N., Toyokuni S., Ito M., Isobe K., Suzuki H.: CD8+CD122+ regulatory T cells (Tregs) and CD4+ Tregs cooperatively prevent and cure CD4+ cell-induced colitis. J. Immunol., 2011; 186: 41-52
[PubMed] [Full Text HTML] [Full Text PDF]
[16] Gieryńska M., Kalinowska-Gacek E.: Błony śluzowe – stan gotowości immunologicznej. Część II. Życie Wet., 2009; 84: 115-122
[Full Text PDF]
[17] Gold M.C., Lewinsohn D.M.: Co-dependents: MR1-restricted MAIT cells and their antimicrobial function. Nat. Rev Microbiol., 2013; 11: 14-19
[PubMed] [Full Text HTML] [Full Text PDF]
[18] Gołąb J., Jakóbisiak M., Lasek W., Stokłosa T.: Immunologia. Wyd. Nauk. PWN Warszawa 2007
[19] Grygorowicz M.A., Kozłowska E.: Udział receptorów TLR rozpoznających wzorce molekularne organizmów patogennych w modulowaniu aktywności regulatorowych limfocytów T CD4+CD25+FoxP3+. Post. Mikrobiol., 2011; 50: 141-154
[Abstract] [Full Text PDF]
[20] Hao J., Dong S., Xia S., He W., Jia H., Zhang S., Wei J., O’Brien R.L., Born W.K., Wu Z., Wang P., Han J., Hong Z., Zhao L., Yin Z.: Regulatory role of Vγ1 γδ T cells in tumor immunity through IL-4 production. J. Immunol., 2011; 187: 4979-4986
[PubMed] [Full Text HTML] [Full Text PDF]
[21] Hirota K., Duarte J.H., Veldhoen M., Hornsby E., Li Y., Cua D.J., Ahlfors H., Wilhelm C., Tolaini M., Menzel U., Garefalaki A., Potocnik A.J., Stockinger B.: Fate mapping of IL-17-producing T cells in inflammatory responses. Nat. Immunol., 2011; 12: 255-263
[PubMed] [Full Text HTML] [Full Text PDF]
[22] Hirota K., Martin B., Veldhoen M.: Development, regulation and functional capacities of Th17 cells. Semin. Immunopathol., 2010; 32: 3-16
[PubMed]
[23] Inagaki-Ohara K., Sakamoto Y., Dohi T., Smith A.L.: γδ T cells play a protective role during infection with Nippostrongylus brasilensis by promoting goblet cells function in the small intestine. Immunology, 2011; 134: 448-458
[PubMed] [Full Text HTML]
[24] Jiang S., Tugulea S., Pennesi G., Liu Z., Mulder A., Lederman S., Harris P., Cortesini R., Suciu-Foca N.: Induction of MHC-class I restricted human suppressor T cell by peptide priming in vitro. Hum. Immunol., 1998; 59: 690-699
[PubMed] [Full Text HTML] [Full Text PDF]
[25] Kasprzycka E., Żak J., Ratomski K., Wysocka J.: Limfocyty atypowe. Pol. Merkur. Lekarski, 2008; 24: 443-445
[PubMed]
[26] Kim H.J., Verbinnen B., Tang X., Lu L., Cantor H.: Inhibition of follicular T-helper cells by CD8+ regulatory T cells is essential for self tolerance. Nature, 2010; 467: 328-332
[PubMed]
[27] Kosmaczewska A., Świerkot J., Ciszak L., Wiland P.: Rola subpopulacji limfocytów pomocniczych Th1, Th17 i Treg w patogenezie reumatoidalnego zapalenia stawów z uwzględnieniem przeciwzapalnego działania cytokin Th1. Postępy Hig. Med. Dośw., 2011; 65: 397-403
[PubMed] [Full Text HTML] [Full Text PDF]
[28] Koyasu S., Moro K.: Type 2 innate immune responses and the natural helper cell. Immunology, 2011; 132: 475-481
[PubMed] [Full Text HTML] [Full Text PDF]
[29] Koyasu S., Moro K., Tanabe M., Takeuchi T.: Natural helper cells: a new player in the innate immune response against helminth infection. Adv. Immunol., 2010; 108: 21-44
[PubMed] [Full Text HTML] [Full Text PDF]
[30] Krausgruber T., Blazek K., Smallie T., Alzabin S., Lockstone H., Sahgal N., Hussell T., Feldmann M., Udalova I.A.: IRF5 promotes inflammatory macrophage polarization and TH1-TH17 responses. Nat. Immunol., 2011; 12: 231-238
[PubMed] [Full Text HTML] [Full Text PDF]
[31] Kwiatek M., Kwaśniewska A.: Rola komórek T regulatorowych w ciąży. Post. Biol. Kom., 2011; 38: 395-406
[Full Text PDF]
[32] Lanier L.L.: Shades of grey – the blurring view of innate and adaptive immunity. Nat. Rev. Immunol., 2013; 13: 73-74
[PubMed]
[33] Larski Z.: Niektóre nowe dane dotyczące wirusologii, prionów i immunologii. Medycyna Wet., 2011; 67: 787-791
[Abstract] [Full Text PDF]
[34] Leavy O.: The TH17 kiss of death for neurons. Nat. Rev. Immunol., 2010; 10: 750
[PubMed]
[35] Liao W., Lin J.X., Leonard W.J.: IL-2 family cytokines: new insights into the complex roles of IL-2 as a broad regulator of T helper cell differentiation. Curr. Opin. Immunol., 2011; 23: 598-604
[PubMed] [Full Text HTML] [Full Text PDF]
[36] Liu B., Tahk S., Yee K.M., Fan G., Shuai K.: The ligase PIAS1 restricts natural regulatory T cell differentiation by epigenetic repression. Science, 2010; 330: 521-525
[PubMed] [Full Text HTML] [Full Text PDF]
[37] Majewska M., Szczepanik M.: Reakcja nadwrażliwości kontaktowej, jej mechanizm i regulacja. Postępy Hig. Med. Dośw., 2009; 63: 47-57
[PubMed] [Full Text HTML] [Full Text PDF]
[38] Maślanka T.: Komórki regulatorowe z populacji limfocytów CD4+. Medycyna Wet., 2010; 66: 827-832
[Abstract] [Full Text PDF]
[39] Maślanka T.: Aktualne poglądy na temat komórek regulatorowych bydła, świni i psa. Medycyna Wet., 2011; 67: 172-176
[Abstract]
[40] McMurchy A.N., Bushell A., Levings M.K., Wood K.J.: Moving to tolerance: clinical application of T regulatory cells. Semin. Immunol., 2011; 23: 304-313
[PubMed] [Full Text HTML] [Full Text PDF]
[41] Mękal A., Trzeciak-Ryczek A., Tokarz-Deptuła B., Deptuła W.: Natural TH2 (nTH2) cells, interleukin 36 and interleukin 37 – new elements of innate immunity. Centr. Eur. J. Immunol., 2011; 36: 113-116
[Abstract] [Full Text HTML] [Full Text PDF]
[42] Mękal A., Trzeciak-Ryczek A., Tokarz-Deptuła B., Działo J., Deptuła W.: Nowe elementy odporności wrodzonej. Post. Biol. Kom., 2011; 38: 349-357
[Full Text PDF]
[43] Moro K., Yamada T., Tanabe M., Takeuchi T., Ikawa T., Kawamoto H., Furusawa J., Ohtani M., Fujii H., Koyasu S.: Innate production of TH2 cytokines by adipose tissue-associated c-Kit+ Sca-1+ lymphoid cells. Nature, 2010; 463: 540-544
[PubMed]
[44] Neill D.R., McKenzie A.N.: TH9 cell generation. TH9: the latest addition to the expanding repertoire of IL-25 targets. Immunol. Cell Biol., 2010; 88: 502-504
[PubMed] [Full Text HTML] [Full Text PDF]
[45] Neill D.R., Wong S.H., Bellosi A., Flynn R.J., Daly M., Langford T.K., Bucks C., Kane C.M., Fallon P.G., Pannell R., Jolin H.E., McKenzie A.N.: Nuocytes represent a new innate effector leukocyte that mediates type-2 immunity. Nature, 2010; 464: 1367-1370
[PubMed]
[46] Niedźwiedzka-Rystwej P., Deptuła W.: Limfocyty Treg, Th17, TFH – fakty znane i nieznane. Alergia Astma Immunologia, 2010; 15: 81-85
[Abstract]
[47] Noelle R.J., Nowak E.C.: Cellular sources and immune functions of interleukin-9. Nat. Rev. Immunol., 2010; 10: 683-687
[PubMed]
[48] Ohkura N., Sakaguchi S.: Maturation of effector regulatory T cells. Nat. Immunol., 2011; 12: 283-284
[PubMed] [Full Text HTML] [Full Text PDF]
[49] Pandiyan P., Conti H.R., Zheng L., Peterson A.C., Mathern D.R., Hernández-Santos N., Edgerton M., Gaffen S.L., Leonardo M.J.: CD4+CD25+Foxp3+ regulatory T cells promote Th17 cells in vitro and enhance host resistance in mouse Candida albicans Th17 cell infection model. Immunity, 2011; 34: 422-434
[PubMed] [Full Text HTML] [Full Text PDF]
[50] Park S.G., Mathur R., Long M., Hosh N., Hao N., Hayden M.S., Ghosh S.: T regulatory cells maintain intestinal homeostasis by suppressing γδ T cells. Immunity, 2010; 33: 791-803
[PubMed] [Full Text HTML] [Full Text PDF]
[51] Peaudecerf L., Rocha B.: Role of the gut as a primary lymphoid organ. Immunol. Lett., 2011; 140: 1-6
[PubMed]
[52] Pekkarinen P.T., Vaali K., Junnikkala S., Rossi L.H., Tuovinen H., Meri S., Vaarala O., Arstila T.P.: A functional complement system is required for normal T helper cell differentiation. Immunobiology, 2011; 216: 737-743
[PubMed] [Full Text HTML] [Full Text PDF]
[53] Pillai M.R., Collison L.W., Wang X., Finkelstein D., Rehg J.E., Boyd K., Szymczak-Workman A.L., Doggett T., Griffith T.S., Ferguson T.A., Vignali D.A.: The plasticity of regulatory T cell function. J. Immunol., 2011; 187: 4987-4997
[PubMed] [Full Text HTML] [Full Text PDF]
[54] Pot C., Apetoh L., Awasthi A., Kuchroo V.K.: Induction of regulatory Tr1 cells and inhibition of TH17 cells by IL27. Semin. Immunol., 2011; 23: 438-445
[PubMed] [Full Text HTML] [Full Text PDF]
[55] Pot C., Apetoh L., Kuchroo V.K.: Type 1 regulatory T cells (Tr1) in autoimmunity. Semin. Immunol., 2011; 23: 202-208
[PubMed] [Full Text HTML] [Full Text PDF]
[56] Rogala E., Nowicka A., Wertel I., Polak G., Tarkowski R., Kotarski J., Kotarski J.: Limfocyty Th17 nowym sojusznikiem w walce z rakiem jajnika? Post. Biol. Kom., 2011; 38: 423-433
[Abstract] [Full Text PDF]
[57] Roncarolo M.G., Gregori S., Battaglia M., Bacchetta R., Fleischhauer K., Levings M.K.: Interleukin-10-secreting type 1 regulatory T cells in rodents and humans. Immunol. Rev., 2006; 212: 28-50
[PubMed] [Full Text HTML] [Full Text PDF]
[58] Saenz S.A., Noti M., Artis D.: Innate immune cell populations function as initiators and effectors in Th2 cytokine responses. Trends Immunol., 2010; 31: 407-413
[PubMed] [Full Text HTML] [Full Text PDF]
[59] Saenz S.A., Siracusa M.C., Perrigoue J.G., Spencer S.P., Urban J.F.Jr., Tocker J.E., Budelsky A.L., Kleinschek M.A., Kastelein R.A., Kambayashi T., Bhandoola A., Artis D.: IL25 elicits a multipotent progenitor cell population that promotes TH2 cytokine responses. Nature, 2010; 464: 1362-1366
[PubMed]
[60] Seidelin J.B., Rogler G., Nielsen O.H.: A role for interleukin-33 in TH2-polarized intestinal inflammation? Mucosal Immunol., 2011; 4: 496-502
[PubMed] [Full Text HTML] [Full Text PDF]
[61] Shevach E.M.: From vanilla to 28 flavors: multiple varieties of T regulatory cells. Immunity, 2006; 25: 195-201
[PubMed] [Full Text HTML] [Full Text PDF]
[62] Spits H., Artis D., Colonna M., Diefenbach A., Di Santo J.P., Eberl G., Koyasu S., Locksley R.M., McKenzie A.N., Mebius R.E., Powrie F., Vivier E.: Innate lymphoid cells – a proposal for uniform nomenclature. Nat. Rev. Immunol., 2013; 13: 145-149
[PubMed] [Full Text HTML] [Full Text PDF]
[63] Strober W.: The expanding TH2 universe. Nature, 2010; 463: 434-435
[PubMed]
[64] Tan C., Ramaswamy M., Shi G., Vistica B.P., Siegel R.M., Gery I.: Inflammation-inducing Th1 and Th17 cells differ in their expression patterns of apoptosis-related molecules. Cell. Immunol., 2011; 271: 210-213
[PubMed] [Full Text HTML] [Full Text PDF]
[65] Trifari S., Kaplan C.D., Tran E.H., Crellin N.K., Spits H.: Identification of a human helper T cell population that has abundant production of interleukin 22 and is distinct from TH-17, TH1 and TH2 cells. Nat. Immunol., 2009; 10: 864-871
[PubMed] [Full Text HTML] [Full Text PDF]
[66] Trifari S., Spits H.: IL-22-producing CD4+ T cells: middle-men between the immune system and its environment. Eur. J. Immunol., 2010; 40: 2369-2371
[PubMed] [Full Text HTML] [Full Text PDF]
[67] Van Stry M., Bix M.: Explaining discordant coordination. Nat. Immunol., 2011; 12: 16-17
[PubMed] [Full Text HTML] [Full Text PDF]
[68] Vinay D.S., Kim C.H., Choi B.K., Kwon B.S.: Origins and functional basis of regulatory CD11c+CD8+ T cells. Eur. J. Immunol., 2009; 39: 1552-1563
[PubMed] [Full Text HTML] [Full Text PDF]
[69] Walker J.A., Barlow J.L., McKenzie A.N.: Innate lymphoid cells – how did we miss them? Nat. Rev. Immunol., 2013; 13: 75-87
[PubMed]
[70] Wan Y.Y.: Multi-tasking of helper T cells. Immunology, 2010; 130: 166-171
[PubMed] [Full Text HTML] [Full Text PDF]
[71] Yu D., Vinuesa C.G.: The elusive identity of T follicular helper cells. Trends Immunol., 2010; 31: 377-383
[PubMed] [Full Text HTML] [Full Text PDF]
[72] Zhao D.M., Thornton A.M., DiPaolo R.J., Shevach E.M.: Activated CD4+CD25+ T cells selectively kill B lymphocytes. Blood, 2006; 107: 3925-3932
[PubMed] [Full Text HTML] [Full Text PDF]
Autorzy deklarują brak potencjalnych konfliktów interesów.