
Hemoglobina – źródłem reaktywnych form tlenu
Ewa Zapora 1 , Iwona Jarocka 1Streszczenie
Erytrocyty to komórki wyjątkowo podatne na działanie reaktywnych form tlenu głównie dlatego, iż biorą udział w transporcie tlenu, przez co są bezpośrednio narażone na jego działanie. Ponadto zawarta w erytrocycie hemoglobina zawiera jony żelaza (II), które umożliwiają zachodzenie reakcji Fentona oraz katalizują peroksydację lipidów. Reaktywne formy tlenu w erytrocycie mogą być również generowane w wyniku nieenzymatycznej i enzymatycznej degradacji hemu. Proces nieenzymatycznej degradacji hemu inicjuje m.in. nadtlenek wodoru, natomiast proces enzymatyczny zachodzi pod wpływem oksygenazy hemowej. W obu przypadkach w wyniku degradacji hemu tworzy się biliwerdyna, tlenek węgla (II) oraz uwalniane są jony żelaza Fe2+. Powstałe produkty degradacji hemu mogą inicjować procesy oksydacyjne w erytrocycie, niemniej jednak w niewielkim stężeniu wykazują również właściwości cytoprotekcyjne.
Słowa kluczowe:reaktywne formy tlenu (RFT), erytrocyt, hemoglobina
Summary
Erythrocytes are especially vulnerable to reactive oxygen species because of their direct role in oxygen transport. Moreover, hemoglobin contains iron ions (Fe2+), which catalyze both the Fenton reaction and lipid peroxidation. Reactive oxygen species in erythrocytes are also generated through nonenzymatic and enzymatic processes of heme degradation. The nonenzymatic process of heme degradation is initiated by e.g. hydrogen peroxide, whereas the process of enzymatic degradation is under the influence of heme oxygenase. In both cases biliverdin, carbon monoxide (CO) and iron ions (Fe2+) are generated. These products of heme degradation can initialize the oxidative processes within erythrocytes, but at low concentrations exhibit cytoprotective properties.
Key words:reactive oxygen species (ROS), erythrocyte, hemoglobin
W ostatnich latach wzrasta zainteresowanie udziałem reakcji wolnorodnikowych w fizjologii oraz w różnych stanach patologicznych. W badaniach nad działaniem reaktywnych form tlenu (RFT) na metabolizm komórkowy szczególnie ciekawym modelem ze względu na specyfikę metaboliczną jest erytrocyt. Ponieważ dojrzałe postaci erytrocytów nie mają mitochondriów ani retikulum endoplazmatycznego, które w typowych komórkach są głównym źródłem RFT, w erytrocytach za ten proces odpowiedzialna jest hemoglobina. Niezależnie od innych funkcji fizjologicznych hemoglobina bierze również udział w reakcjach oksydoredukcyjnych. Ma to związek z tym, iż:
• erytrocyt jest komórką transportującą tlen cząsteczkowy, który stanowi potencjalne źródło RFT;
• błony erytrocytarne zawierają duże ilości wielonienasyconych kwasów tłuszczowych, które w środowisku bogatym w tlen i rodniki tlenowe mogą być źródłem rodników lipidowych;
• hemoglobina zawiera jony żelaza (II), które umożliwiają zachodzenie reakcji Fentona generującej rodnik hydroksylowy oraz katalizują peroksydację lipidów;
• hemoglobina jest odpowiedzialna za generację reaktywnych form tlenu w wyniku enzymatycznej i nieenzymatycznej degradacji hemu.
Budowa i funkcje hemoglobiny
Hemoglobina (Hb) jest białkiem należącym do hemoporfiryn. Jest to sferyczny tetramer, w skład którego wchodzi globina złożona z dwóch łańcuchów polipeptydowych α i dwóch łańcuchów β, a także czterech podjednostek hemu. Główną funkcją hemoglobiny jest transport tlenu z płuc do tkanek oraz ditlenku węgla z różnych tkanek do płuc.
Proces wiązania tlenu przez hemoglobinę jest związany ze zmianami w konformacji cząsteczki hemoglobiny i prowadzi do zmiany stopnia utlenienia centralnie położonego jonu żelaza. W związku z tym wyróżnia się kilka postaci hemoglobiny (ryc. 1). W warunkach fizjologicznych postać zredukowana, tzw. deoksyhemoglobina – deoksyHb wiąże w kapilarach płucnych tlen i przechodzi w postać utlenioną – oksyhemoglobinę – oksyHb [HbFe(II)O2] [27]. OksyHb może zostać utleniona do zawierającej jon Fe3+ methemoglobiny – metHb [HbFe(III)], która ma zdolność wiązania cząsteczek wody, ale nie ma zdolności wiązania tlenu. Istnieje także ferrylohemoglobina – ferryloHb, która powstaje w wyniku utleniania deoksyhemoglobiny z udziałem nadtlenku wodoru H2O2 [13].
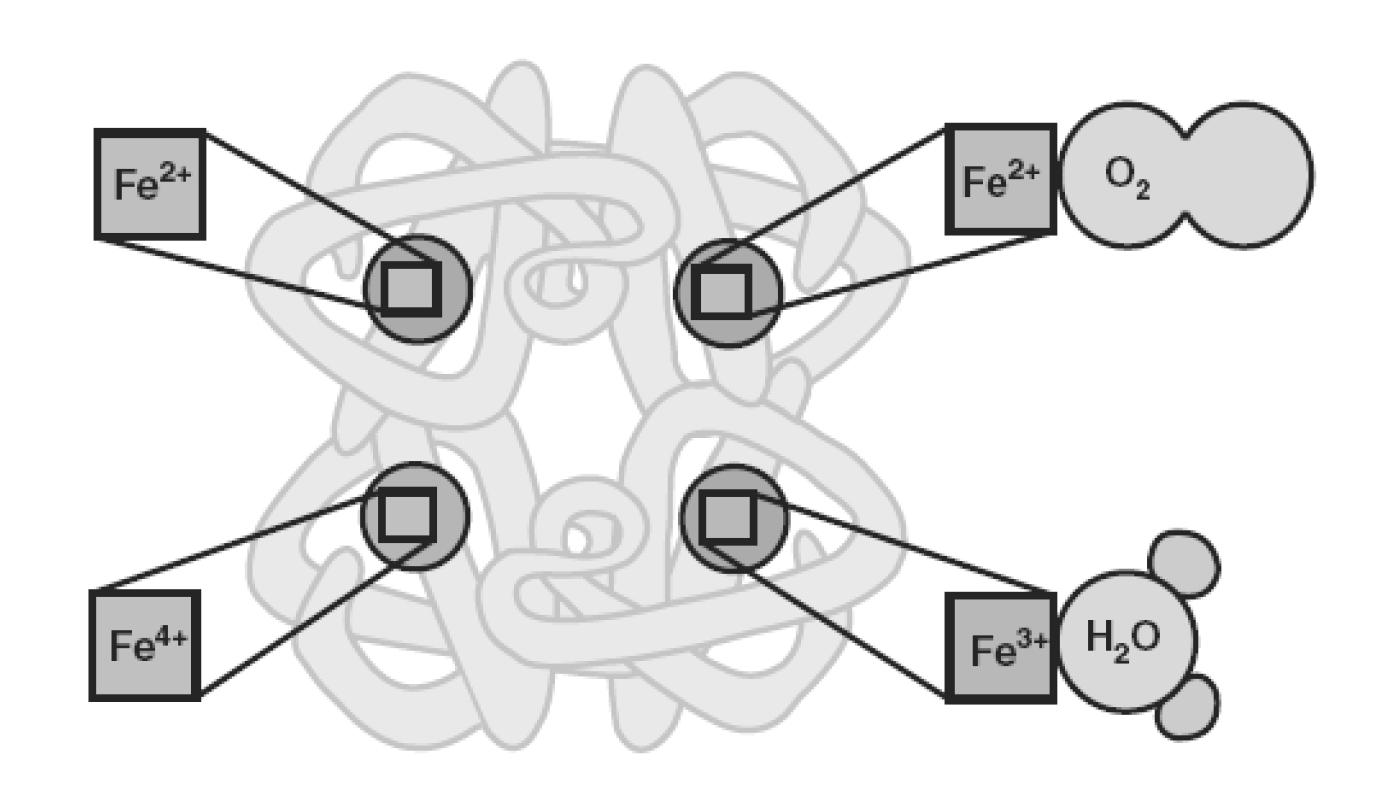
Ryc. 1. Różne postaci hemoglobiny (wg [13] zmodyfikowano)
Konformacja oksyhemoglobiny określana jest jako stan R (relaxed – rozluźniony). Natomiast konformacja deoksyhemoglobiny określana jest jako stan T (taut – naprężony). Przyłączenie cząsteczki tlenu do jednej z czterech cząstek hemu hemoglobiny powoduje zmianę struktury drugo-, trzecio- i czwartorzędowej całego tetrameru. Hemoglobina w stanie R wykazuje większe powinowactwo do tlenu aniżeli w stanie T. W związku z tym każde przyłączenie w płucach cząsteczki tlenu do hemoglobiny ułatwia przyłączanie następnych (tzw. wiązanie kooperacyjne), zaś odczepienie każdej cząsteczki tlenu w tkankach ułatwia uwalnianie kolejnych cząstek tlenu [9].
Oksyhemoglobina jest postacią stabilną, niemniej jednak pod wpływem czynników utleniających jon Fe2+ w cząsteczce hemu może być utleniony do jonu Fe3+, czemu towarzyszy powstanie methemoglobiny, która nie może przenosić tlenu. W warunkach fizjologicznych 0,5-3% hemoglobiny dziennie ulega autooksydacji do methemoglobiny [46].
Hemoglobina pełni istotną rolę nie tylko w transporcie tlenu, ale także w transporcie CO2 z tkanek do płuc. Około 10% CO2 przenoszone jest w postaci rozpuszczonej we krwi, natomiast ~30% w postaci związanej z grupami aminowymi białek, a ~60% w postaci jonów wodorowęglanowych (IV) (HCO3-). Ponadto hemoglobina odgrywa również znaczącą rolę w transporcie tlenku azotu (NO), którego główna rola w układzie krwionośnym polega na rozszerzaniu naczyń krwionośnych. Tlenek azotu łącząc się z grupą tiolową reszty cysteiny tworzy nitrozotiole, które (głównie nitrozoalbumina i nitrozoglutation) mają właściwości podobne do tych, które wykazuje NO, z tym że dodatkowo hamują agregację płytek krwi [10]. Powinowactwo hemoglobiny do tlenku azotu jest duże, jeśli hemoglobina znajduje się w konformacji R, a małe w przypadku konformacji T [3]. Tlenek azotu wiązany przez hemoglobinę pochodzi z zewnątrz erytrocytu, choć istnieją również doniesienia o obecności syntazy tlenku azotu wewnątrz erytrocytu [12].
Hemoglobina oprócz swej podstawowej funkcji transportowej, uczestniczy również w wielu innych procesach, przy czym niektóre z tych aktywności mogą mieć znaczenie fizjologiczne. Chociaż nie jest białkiem enzymatycznym, hemoglobina w niektórych przypadkach wykazuje działanie pseudoenzymatyczne, np. wykazuje aktywność katalazy, przy czym porównanie szybkości rozkładu nadtlenku wodoru przez hemolizaty myszy zdrowych i myszy z akatalazemią (dziedzicznym brakiem katalazy) wskazuje, że hemoglobina wykazuje około 10% aktywności katalazy obecnej w erytrocytach [20]. Hemoglobina ma również aktywność peroksydazową [15]. Może utleniać m.in. dopaminę, o-dianizydynę, gwajakol, pirogallol i 2,2′-azynobis(3-etylobenzotiazolinosulfonian) (ABTS) [22,39]. Hemoglobina jest jednak 10-100 razy mniej aktywna niż właściwe peroksydazy [8]. Wykazano, iż hemoglobina wykazuje również zależną od cytochromu P-450 aktywność monooksygenazową, esterazową (hydrolizuje estry aromatyczne), a także aktywność O– i N-demetylazy [35,37]. Ponadto katalizuje także oksydatywną kondensację aminofenoli [43].
Generacja RFT w wyniku nieenzymatycznej degradacji hemu
Zasadnicze znaczenie w generacji RFT w erytrocytach ma proces degradacji hemu, który może zachodzić zarówno enzymatycznie jak i nieenzymatycznie. Anionorodnik ponadtlenkowy O2–. pierwszy i podstawowy spośród RFT, powstaje w erytrocycie głównie w wyniku autooksydacji hemoglobiny, tj. spontanicznego przejścia oksyhemoglobiny w methemoglobinę (reakcja 1) [28].

Jon Fe2+ w oksyhemoglobinie jest oporny na utlenianie, istnieje jednak możliwość przeniesienia elektronu w kierunku tlenu, w efekcie czego utlenowana hemoglobina pozostaje w równowadze z methemoglobiną (zawierającą jon Fe3+) i anionorodnikiem ponadtlenkowym. Wprawdzie równowaga tej reakcji jest zdecydowanie przesunięta w kierunku oksyhemoglobiny, niemniej jednak około 3% oksyhemoglobiny utlenia się w ciągu doby do methemoglobiny, co oznacza, że jednocześnie uwalniany jest anionorodnik ponadtlenkowy. Jednak obecność w erytrocycie zależnej od NADPH reduktazy methemoglobinowej (Cytb5R) umożliwia ponowną regenerację oksyhemoglobiny (reakcja 2) [23]:

Poza procesami autooksydacji hemoglobiny, białko to narażone jest na oddziaływanie anionorodników ponadtlenkowych wytwarzanych w wyniku aktywacji innych morfotycznych elementów krwi, głównie fagocytów. Na powierzchni komórek fagocytarnych, m.in. neutrofilów, eozynofilów czy makrofagów znajduje się system zależny od oksydazy NADPH, generujący anionorodnik ponadtlenkowy [2]. Ponadto anionorodnik ponadtlenkowy może powstawać również w reakcji utlenienia amin katecholowych i niektórych ksenobiotyków [21,45].
W wyniku dysmutacji anionorodnika ponadtlenkowego dochodzi do powstania H2O2, który bierze udział w degradacji hemu. Mechanizm oddziaływania H2O2 z hemem opiera się na reakcji utleniania żelaza, która jest procesem zależnym od pH środowiska [29]. Stwierdzono, iż hem wchodzi w reakcję z anionem wodoronadtlenkowym, który powstaje w reakcji:

W reakcji anionu wodoronadtlenkowego z hemoglobiną powstaje kompleks Fe3+– nadtlenek, a następnie biliwerdyna i tlenek węgla (II) [11]:

Jeżeli zdolności antyoksydacyjne erytrocytu są obniżone, to może dochodzić do nagromadzenia nadtlenku wodoru, który może reagować zarówno z oksyhemoglobiną, methemoglobiną jak i deoksyhemoglobiną (reakcja 5, 6, 7) (rycina 2C,D,E).
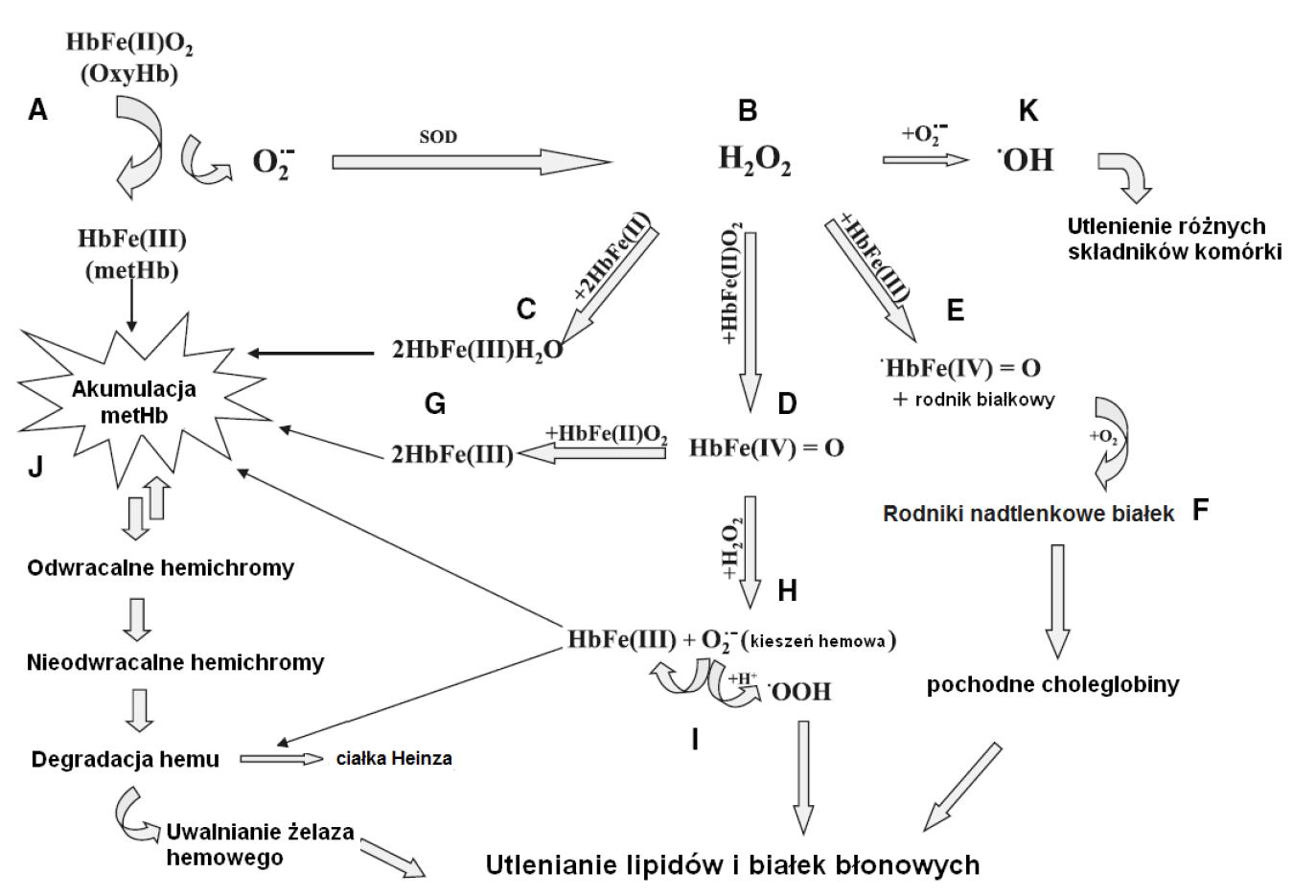
Ryc. 2. Udział hemoglobiny w procesach oksydacyjnych w erytrocycie (wg [13] zmodyfikowano)
Powyższe procesy przyczyniają się do powstania odpowiednio: ferrylohemoglobiny [HbFe(IV)=O], oksoferrylohemoglobiny [•HbFe(IV)=O] i methemoglobiny [HbFe(III)]:



Powstałe w reakcjach (5) i (6) hemoglobiny są czynnikami silnie utleniającymi [28,42,44]. Ferrylohemoglobina może albo ulec reakcji synproporcjonowania z oksyhemoglobiną (reakcja 8) (rycina 2G) albo też może ulec dalszemu utlenieniu z udziałem H2O2 z utworzeniem methemoglobiny i anionorodnika ponadtlenkowego (reakcja 9) (rycina 2H), co w konsekwencji wzmaga proces degradacji hemu [13]:


Dysmutaza ponadtlenkowa nie ma możliwości przeniknięcia do wnętrza „kieszeni” hemowych i katalizowania reakcji dysmutacji, dlatego też w związku z obecnością anionorodników, może dochodzić do utlenienia pierścieni tetrapirolowych [1].
Wytwarzanie anionorodnika ponadtlenkowego jest częściowo ograniczane poprzez konkurencyjną reakcję (10) między oksoferrylohemoglobiną i H2O2, w której zamiast anionorodnika generowany jest tlen [28]:

Badania wykazały ponadto, iż w reakcji H2O2 i methemoglobiny (rycina 2E) powstają również rodniki białkowe, które mogą następnie reagować z tlenem z utworzeniem rodników nadtlenkowych białek (rycina 2F), a także mogą się przyczynić do rozkładu lipofilowych pochodnych choleglobiny, które wbudowane między lipidy błony komórkowej indukują dalsze uszkodzenia oksydacyjne [42]. Z kolei uwodornienie anionorodnika znajdującego się wewnątrz „kieszeni” hemowych prowadzi do wytworzenia rodnika wodoronadtlenkowego (•OOH) (rycina 2I), będącego inicjatorem procesu peroksydacji lipidów w błonie erytrocytarnej [28].
Akumulacja methemoglobiny (rycina 2J) prowadzi do powstania hemichromów będących nieaktywną postacią hemoglobiny. W początkowym etapie ich tworzenia dochodzi do utlenienia grup tiolowych hemoglobiny, czego konsekwencją są zmiany konformacyjne łańcuchów globiny [38,44]. Wykazano, iż przemianom hemoglobiny towarzyszy degradacja hemu i utrata żelaza hemowego, co w konsekwencji prowadzi do precypitacji zdenaturowanych cząsteczek hemoglobiny i powstawania złogów, tzw. ciałek Heinza. Wykazano, iż powstałe produkty degradacji hemu inicjują procesy oksydacyjne w erytrocycie [5,33]. Obecność zarówno O2–. jak i H2O2 przyczynia się do wytworzenia rodników hydroksylowych (rycina 2K) w reakcji Habera-Weissa, katalizowanej przez uwolnione podczas degradacji hemu jony żelaza [26]. Ponadto uwolniony hem hamuje aktywność enzymów cytosolowych oraz nieswoiście reaguje z białkami błonowymi erytrocytu powodując utlenienie ich grup sulfhydrylowych [27]. W efekcie tworzą się niepożądane wiązania krzyżowe, co prowadzi do utraty funkcji białek i ich agregacji.
Generacja RFT w wyniku enzymatycznej degradacji hemu
Niezależnie od procesów nieenzymatycznych, hem ulega również degradacji enzymatycznej. Najważniejszą rolę w tym procesie odgrywa oksygenaza hemowa (HO) [EC 1.14.99.3]. Występuje ona w postaci 3 izoform: HO-1, HO-2 oraz HO-3. Z dwóch funkcjonalnych izoform (HO- 1 i HO-2) HO-1 jest łatwo indukowalna i ulega ekspresji w wyniku działania różnych bodźców stymulujących, m.in. cytokin czy jonów metali przejściowych [24,34,36]. Ponadto w ekspresji izoformy HO-1 biorą udział czynniki transkrypcyjne (AP-1, NF-κB) oraz współdziałające z nimi kinazy (ERK, JUN, p38MAPK) [16]. Indukcja HO-1 może prowadzić do nawet stukrotnego wzrostu aktywności enzymu. Uważa się, że działanie HO-1 jest najprawdopodobniej związane z mechanizmami obronnymi przed stresem oksydacyjnym.
Początkowo uważano, że głównym efektem działania oksygenazy hemowej HO-1 jest proces degradacji wolnego hemu, w którym HO-1 z udziałem tlenu cząsteczkowego i NADPH rozkłada hem do tlenku węgla (II) [CO], jonów żelaza Fe2+ oraz rozpuszczalnej biliwerdyny (ryc. 3), która jest następnie przekształcana przez reduktazę biliwerdyny w nierozpuszczalną bilirubinę [6,34].

Ryc. 3. Enzymatyczna degradacja hemu (wg [41] zmodyfikowano)
Produkty rozkładu hemu w wysokich stężeniach są toksyczne. Zarówno jony Fe2+ jak i wolny hem uczestnicząc w reakcji Fentona (reakcja 11) katalizują powstawanie RFT [6,47]. Wykazano jednak, iż niewielkie ich ilości mają również właściwości cytoprotekcyjne. Zwiększona ekspresja HO-1 oraz uwalnianie jonów Fe2+ pociąga za sobą jednoczesny wzrost ekspresji ferrytyny. Ferrytyna oprócz wiązania jonów Fe2+ wykazuje działanie cytoprotekcyjne oraz antyapoptotyczne [4,19].
Powstała z rozkładu hemu biliwerdyna jest natychmiast przekształcana w bilirubinę. Związek ten w postaci wolnej (niezwiązany z glukuronianem) jest toksyczny. Po przekroczeniu bariery krew-mózg, wywołuje żółtaczkę jąder podkorowych mózgu, co skutkuje uszkodzeniem komórek nerwowych [47]. Wykazano, że w małych stężeniach bilirubina i biliwerdyna działają przeciwzapalnie i przeciwutleniająco, gdyż wychwytują reaktywne formy tlenu i azotu [34]. Ponadto biliwerdyna hamuje interakcje neutrofilów z komórkami śródbłonka oraz aktywację limfocytów [30].
Najintensywniej badanym produktem rozkładu hemu jest tlenek węgla (II) (CO). W warunkach fizjologicznych w organizmie człowieka ponad 85% tlenku węgla (II) pochodzi z rozpadu hemu [6] Związek ten wiąże się z hemoglobiną z wytworzeniem karboksyhemoglobiny i w tej postaci transportowany jest do płuc, gdzie jest wydzielany w wydychanym powietrzu. Tlenek węgla, podobnie jak tlenek azotu, pełni istotną rolę w sygnalizacji międzykomórkowej. Aktywując cyklazę guanylową, powoduje wzrost cGMP, dzięki czemu wykazuje działanie antykoagulacyjne [48]. Poprzez kaskady kinaz MAP hamuje wydzielanie cytokin prozapalnych: IL-1β, IL-6, TNF-α, oraz MIP1α [18,32]. Tlenek węgla zmniejsza także ekspresję cząsteczek adhezyjnych, jednocześnie nasilając ekspresję przeciwzapalnej IL-10 [48]. Ponadto, dzięki aktywacji białka p38 hamuje apoptozę [49]. W niektórych typach komórek, m.in. w limfocytach, tlenek węgla działa antyproliferacyjnie [34,37], jednak w przypadku komórek śródbłonka może zwiększać ich proliferację [19]. Tlenek węgla (II) wpływa także na przekazywanie sygnałów z niektórych receptorów TLR (Toll like receptors), istotnych w inicjacji odpowiedzi immunologicznej przeciwko patogenom [31]. Rozważa się możliwość wykorzystania związków uwalniających CO (tzw. carbon monoxide releasing molecule – CORM) w terapii chorób układu krążenia [7,25].
Badania in vitro wykazały dwukierunkowe działania oksygenazy hemowej. Stwierdzono, że 2,5-krotny wzrost aktywności oksygenazy hemowej HO-1 chroni komórki przed toksycznym działaniem tlenu, natomiast ponad 15-krotny wzrost aktywności zwiększa jej toksyczność [40]. Produkty rozpadu hemu, a zwłaszcza biliwerdyna/ bilirubina oraz tlenek węgla wykazują właściwości cytoprotekcyjne i ograniczają skutki działania wielu czynników szkodliwych. Ponadto stwierdzono, iż degradacji hemu z udziałem oksygenazy hemowej w warunkach in vitro towarzyszy generacja nadtlenku wodoru H2O2, który bierze udział zarówno w bezpośrednim procesie niszczenia hemu, jak i inaktywuje samą oksygenazę hemową HO-1 [26].
Produkty degradacji hemu nasilając generację reaktywnych form tlenu, powodują oksydacyjne modyfikacje biologicznie ważnych składników erytrocytu, co ma istotny udział w patogenezie chorób układu krążenia.
PIŚMIENNICTWO
[1] Alayash A.I.: Oxygen therapeutics: can we tame haemoglobin? Nat. Rev. Drug Discov., 2004; 3: 152-159
[PubMed]
[2] Bylund J., Brown K.L., Movitz C., Dahlgren C., Karlsson A.: Intracellular generation of superoxide by the phagocyte NADPH oxidase: how, where, and what for? Free Radic. Biol. Med., 2010; 49: 1834-1845
[PubMed]
[3] Carlsen C.U., Moller J.K., Skibsted L.H.: Heme-iron in lipid oxidation. Coord. Chem. Rev., 2005; 249: 485-498
[4] Chiu H., Brittingham J.A., Laskin D.L.: Differential induction of heme oxygenase-1 in macrophages and hepatocytes during acetaminophen-induced hepatotoxicity in the rat: effects of hemin and biliverdin. Toxicol. Appl. Pharmacol., 2002; 181: 106-115
[PubMed]
[5] Dani C., Masini E., Bertini G., di Felice A.M., Pezzati M., Ciofini S., Rossetti M., Gallori G., Ciuti R., Buonocore G., Paffetti P., Mannaioni P.F., Rubaltelli F.F.: Role of heme oxygenase and bilirubin in oxidative stress in preterm infants. Pediatr. Res., 2004; 56: 873-877
[PubMed] [Full Text HTML] [Full Text PDF]
[6] Florczyk U.M., Józkowicz A., Dulak J.: Biliverdin reductase: new features of an old enzyme and its potential therapeutic significance. Pharmacol. Rep., 2008; 60: 38-48
[PubMed] [Full Text PDF]
[7] Foresti R., Hammad J., Clark J.E., Johnson T.R., Mann B.E., Friebe A., Green C.J., Motterlini R.: Vasoactive properties of CORM-3, a novel water-soluble carbon monoxide-releasing molecule. Br. J. Pharmacol., 2004; 142: 453-460
[PubMed] [Full Text HTML] [Full Text PDF]
[8] Giulivi C., Davies K.J.: Mechanism of the formation and proteolytic release of H2O2-induced dityrosine and tyrosine oxidation products in hemoglobin and red blood cells. J. Biol. Chem., 2001; 276: 24129-24136
[PubMed] [Full Text HTML] [Full Text PDF]
[9] Harmening D.M.: Clinical Hematology and Fundamentals of Hemostasis, wyd. 4, F.A.Davis, Philadelphia, 2002
[10] Herold S., Rock G.: Reactions of deoxy-, oxy-, and methemoglobin with nitrogen monoxide. Mechanistic studies of the S-nitrosothiol formation under different mixing conditions. J. Biol. Chem., 2003; 278: 6623-6634
[PubMed] [Full Text HTML] [Full Text PDF]
[11] Hersleth H.P., Hsiao Y.W., Ryde U., Görbitz C.H., Andersson K.K.: The crystal structure of peroxymyoglobin generated through cryoradiolytic reduction of myoglobin compound III during data collection. Biochem. J., 2008; 412: 257-264
[PubMed] [Full Text HTML] [Full Text PDF]
[12] Kang E.S., Ford K., Grokulsky G., Wang Y.B., Chiang T.M., Acchiardo S.R.: Normal circulating adult human red blood cells contain inactive NOS proteins. J. Lab. Clin. Med., 2000; 135: 444-451
[PubMed]
[13] Kanias T., Acker J.P.: Biopreservation of red blood cells – the struggle with hemoglobin oxidation. FEBS J., 2010; 277: 343-356
[PubMed] [Full Text HTML] [Full Text PDF]
[14] Kapitulnik J.: Bilirubin: an endogenous product of heme degradation with both cytotoxic and cytoprotective properties. Mol. Pharmacol., 2004, 66: 773-779
[PubMed] [Full Text HTML] [Full Text PDF]
[15] Kapralov A., Vlasova I.I., Feng W., Maeda A., Walson K., Tyurin V.A., Huang Z., Aneja R.K., Carcillo J., Bayir H., Kagan V.E.: Peroxidase activity of hemoglobin-haptoglobin complexes: covalent aggregation and oxidative stress in plasma and macrophages. J. Biol. Chem., 2009; 284: 30395-30407
[PubMed] [Full Text HTML] [Full Text PDF]
[16] Kietzmann T., Samoylenko A., Immenschuh S.: Transcriptional regulation of heme oxygenase-1 gene expression by MAP kinases of the JNK and p38 pathways in primary cultures of rat hepatocytes. J. Biol. Chem., 2003; 278: 17927-17936
[PubMed] [Full Text HTML] [Full Text PDF]
[17] Kim E.Y., Kim J.S., Kim M.Y., Koh W.S., Guengerich F.P., Yun C.H.: Non-specific inhibition of human cytochrome P450-catalyzed reactions by hemin. Toxicol. Lett., 2004; 153: 239-246
[PubMed]
[18] Kim H.P., Choi A.M.: A new road to induce heme oxygenase-1 expression by carbon monoxide. Circ. Res., 2007; 101: 862-864
[PubMed] [Full Text HTML] [Full Text PDF]
[19] Łoboda A., Jaźwa A., Grochot-Przeczek A., Rutkowski A.J., Cisowski J., Agarwal A., Józkowicz A., Dulak J.: Heme oxygenase-1 and the vascular bed: from molecular mechanisms to therapeutic opportunities. Antioxid. Redox Signal., 2008; 10: 1767-1812
[PubMed]
[20] Masuoka N., Kodama H., Abe T., Wang D.H., Nakano T.: Characterization of hydrogen peroxide removal reaction by hemoglobin in the presence of reduced pyridine nucleotides. Biochim. Biophys. Acta, 2003; 1637: 46-54
[PubMed] [Full Text HTML] [Full Text PDF]
[21] McCord J.M.: The evolution of free radicals and oxidative stress. Am. J. Med., 2000; 108: 652-659
[PubMed]
[22] Miyazaki K., Arai S., Iwamoto T., Takasaki M., Tomoda A.: Metabolism of pyrogallol to purpurogallin by human erythrocytic hemoglobin. Tohoku J. Exp. Med., 2004; 203: 319-330
[PubMed] [Full Text PDF]
[23] Moore T.J., Walsh C.S., Cohen M.R.: Reported adverse event cases of methemoglobinemia associated with benzocaine products. Arch. Intern. Med., 2004; 164: 1192-1196
[PubMed] [Full Text HTML] [Full Text PDF]
[24] Morse D., Choi A.M.: Heme oxygenase-1: the „emerging molecule” has arrived. Am. J. Respir. Cell Mol. Biol., 2002; 27: 8-16
[PubMed] [Full Text HTML] [Full Text PDF]
[25] Motterlini R., Sawle P., Hammad J., Bains S., Alberto R., Foresti R., Green C.J.: CORM-A1: a new pharmacologically active carbon monoxide-releasing molecule. FASEB J., 2005; 19: 284-286
[PubMed] [Full Text PDF]
[26] Nagababu E., Chrest F.J., Rifkind J.M.: Hydrogen-peroxide-induced heme degradation in red blood cells: the protective roles of catalase and glutathione peroxidase. Biochim. Biophys. Acta, 2003; 1620: 211-217
[PubMed]
[27] Nagababu E., Mohanty J.G., Bhamidipaty S., Ostera G.R., Rifkind J.M.: Role of the membrane in the formation of heme degradation products in red blood cells. Life Sci., 2010; 86: 133-138
[PubMed]
[28] Nagababu E., Rifkind J.M.: Heme degradation during autoxidation of oxyhemoglobin. Biochem. Biophys. Res. Commun., 2000; 273: 839-845
[PubMed]
[29] Nagababu E., Rifkind J.M.: Reaction of hydrogen peroxide with ferrylhemoglobin: superoxide production and heme degradation. Biochemistry, 2000; 39: 12503-12511
[PubMed]
[30] Naito Y., Takagi T., Yoshikawa T.: Heme oxygenase-1: a new therapeutic target for inflammatory bowel disease. Aliment. Pharmacol. Ther., 2004; 20 (Suppl. 1): 177-184
[PubMed] [Full Text HTML] [Full Text PDF]
[31] Nakahira K., Kim H.P., Geng X.H., Nakao A., Wang X., Murase N., Drain P.F., Wang X., Sasidhar M., Nabel E.G., Takahashi T., Lukacs N.W., Ryter S.W., Morita K., Choi A.M.: Carbon monoxide differentially inhibits TLR signaling pathways by regulating ROS-induced trafficking of TLRs to lipid rafts. J. Exp. Med., 2006; 203: 2377-2389
[PubMed] [Full Text HTML] [Full Text PDF]
[32] Ning W., Choi A.M., Li C.: Carbon monoxide inhibits IL-17-induced IL-6 production through the MAPK pathway in human pulmonary epithelial cells. Am. J. Physiol. Lung Cell. Mol. Physiol., 2005; 289: L268-L273
[PubMed] [Full Text HTML] [Full Text PDF]
[33] Pradines B., Ramiandrasoa F., Hammadi A., Henry M., Briolant S., Orlandi-Pradines E., Bogreau H., Mosnier J., Baret E., Kunesch G., Le Bras J., Rogier C.: Generation of free radicals and enhancement of hemin-induced membrane damage by a catechol iron chelator in Plasmodium falciparum. J. Biol. Sci., 2005; 5: 463-471
[Abstract] [Full Text PDF]
[34] Ryter S.W., Alam J., Choi A.M.: Heme oxygenase-1/carbon monoxide: from basic science to therapeutic applications. Physiol. Rev., 2006; 86: 583-650
[PubMed] [Full Text HTML] [Full Text PDF]
[35] Sen S., Bose T., Roy A., Chakraborti A.S.: Effect of non-enzymatic glycation on esterase activities of hemoglobin and myoglobin. Mol. Cell. Biochem., 2007; 301: 251-257
[PubMed]
[36] Silva G., Cunha A., Grégoire I.P., Seldon M.P., Soares M.P.: The antiapoptotic effect of heme oxygenase-1 in endothelial cells involves the degradation of p38α MAPK isoform. J. Immunol., 2006; 177: 1894-1903
[PubMed] [Full Text HTML] [Full Text PDF]
[37] Song R., Mahidhara R.S., Zhou Z., Hoffman R.A., Seol D.W., Flavell R.A., Billiar T.R., Otterbein L.E., Choi A.M.: Carbon monoxide inhibits T lymphocyte proliferation via caspase-dependent pathway. J. Immunol., 2004; 172: 1220-1226
[PubMed] [Full Text HTML] [Full Text PDF]
[38] Sugawara Y., Kadono E., Suzuki A., Yukuta Y., Shibasaki Y., Nishimura N., Kameyama Y., Hirota M., Ishida C., Higuchi N., Haramoto K., Sakai Y., Soda H.: Hemichrome formation observed in human haemoglobin A under various buffer conditions. Acta Physiol. Scand., 2003; 179: 49-59
[PubMed]
[39] Sun W., Jiang H., Jiao K.: Electrochemical determination of hydrogen peroxide using ο-dianisidine as substrate and hemoglobin as catalyst. J. Chem. Sci., 2005; 117: 317-322
[Full Text PDF]
[40] Suttner D.M., Dennery P.A.: Reversal of HO-1 related cytoprotection with increased expression is due to reactive iron. FASEB J., 1999; 13: 1800-1809
[PubMed] [Full Text HTML] [Full Text PDF]
[41] Szade A., Dulak J., Józkowicz A.: The role of heme oxygenase-1 in the inflammatory bowel diseases. Przegl. Gastroenterol., 2009; 4: 283-287
[Abstract] [Full Text HTML] [Full Text PDF]
[42] Sztiller M., Puchała M., Kowalczyk A., Bartosz G.: The influence of ferrylhemoglobin and methemoglobin on the human erythrocyte membrane. Redox Rep., 2006; 11: 263-271
[PubMed]
[43] Tao W., Liu Y., Pan D., Nie L., Yao S.: Study on the enhancement of catalytic activity for hemoglobin by quinhydrone in poly(o-aminophenol) film. Bioelectrochemistry, 2004; 65: 51-58
[PubMed]
[44] Telen M.J., Kaufman R.E.: The mature erythrocyte. W: Wintrobe’s clinical hematology, wyd. 11, red.: J.P. Greer, J. Forester i in., Williams and Wilkins, Lippincot, 2004, 230
[45] Terland O., Almas B., Flatmark T., Andersson K.K., Sorlie M.: One-electron oxidation of catecholamines generates free radicals with an in vitro toxicity correlating with their lifetime. Free Radic. Biol. Med., 2006; 41: 1266-1271
[PubMed]
[46] Umbreit J.: Methemoglobin – it’s not just blue: a concise review. Am. J. Hematol., 2007; 82: 134-144
[PubMed] [Full Text PDF]
[47] Wagner K.R., Sharp F.R., Ardizzone T.D., Lu A., Clark J.F.: Heme and iron metabolism: role in cerebral hemorrhage. J. Cereb. Blood Flow Metab., 2003; 23: 629-652
[PubMed] [Full Text HTML] [Full Text PDF]
[48] Wang R., Wang Z., Wu L., Hanna S.T., Peterson-Wakeman R.: Reduced vasorelaxant effect of carbon monoxide in diabetes and the underlying mechanisms. Diabetes, 2001; 50: 166-174
[PubMed] [Full Text HTML] [Full Text PDF]
[49] Zhang X., Shan P., Otterbein L.E., Alam J., Flavell R.A., Davis R.J., Choi A.M., Lee P.J.: Carbon monoxide inhibition of apoptosis during ischemia-reperfusion lung injury is dependent on the p38 mitogen-activated protein kinase pathway and involves caspase 3. J. Biol. Chem., 2003; 278: 1248-1258
[PubMed] [Full Text HTML] [Full Text PDF]
Autorki deklarują brak potencjalnych konfliktów interesów.