
Karnityna: funkcja, metabolizm i znaczenie w niewydolności wątroby u pacjentów w przewlekłym zatruciu alkoholem
Alina Kępka 1 , Sławomir Szajda 2 , Napoleon Waszkiewicz 3 , Paweł Płudowski 1 , Sylwia Chojnowska 4 , Michał Rudy 5 , Agata Szulc 3 , Jerzy Ładny 2 , Krzysztof Zwierz 6Streszczenie
Alkoholizm jest jednym z najczęstszych uzależnień człowieka prowadzącym do wyniszczenia organizmu i śmierci. Chroniczne nadużywanie alkoholu zmniejsza utlenianie kwasów tłuszczowych przez zaburzenie metabolizmu karnityny i cyklu kwasu cytrynowego. Zablokowanie przez alkohol i jego metabolity cyklu kwasu cytrynowego jest częściowo kompensowane przez zwiększenie wytwarzania ciał ketonowych, powodującego ketonemię. Przewlekłe nadużycie alkoholu uszkadza wątrobę, powoduje jej zapalenie, marskość, ogniskową martwicę oraz stłuszczenie.
L-karnityna (kwas L-3-hydroksy-4-N, N, N-trimetyloaminomasłowy) odgrywa główną rolę w transporcie aktywowanych długołańcuchowych kwasów tłuszczowych do miejsc β-oksydacji w mitochondriach. Karnityna uczestniczy również w stabilizacji błon komórkowych przez usunięcie długołańcuchowych acylo-CoA i nadmiaru acylowych grup z organizmu. L-karnityna może być użytecznym i bezpiecznym środkiem terapeutycznym w patologii wątroby wywołanej przez długotrwałe nadużywanie etanolu.
Słowa kluczowe:karnityna • alkohol • stłuszczenie wątroby • marskość wątroby • przewlekłe zatrucie alkoholem
Summary
Alcoholism is one of the most frequent dependences among people, leading to damage of the liver and death of the person. Chronic alcohol consumption decreases fatty acid oxidation by interfering with carnitine metabolism and citric acid cycle activity. Block in activity of the citric acid cycle caused by alcohol and its metabolites is partially compensated by increased ketone body production, which results in ketosis. Chronic administration of alcohol induces liver injury, inflammation, cirrhosis, focal necrosis and steatosis.
L-carnitine (L-3-hydroxy-4-N, N, N-trimethylaminebutyric acid) is an essential factor in fatty acid metabolism, which plays a major role in transport of activated long-chain fatty acids to sites of β-oxidation in mitochondria. Carnitine also stabilizes cell membranes by removing long-chain acyl-CoA and excess of the acyl group from the body. L-carnitine can be a useful and safe drug in the liver pathology induced by chronic ethanol exposure.
Key words:carnitine • alcohol • hepatic steatosis • hepatic cirrhosis • chronic alcohol intoxication
Wprowadzenie
Nadużywanie alkoholu etylowego przez coraz młodszych ludzi stało się poważnym problemem społecznym i zdrowotnym. Od dawna wiadomo, iż alkohol i jego metabolity działają toksycznie na komórkę wątrobową. Etanol indukuje m.in. hiperlipidemię i prowadzi do stłuszczenia wątroby [46,60], powstawania w wątrobie nacieków zapalnych z ogniskami martwicy oraz do włóknienia, a w końcu do marskości. Chroniczne nadużywanie alkoholu może prowadzić do uszkodzenia: ośrodkowego układu nerwowego (OUN), nerek, żołądka i jelit. Alkohol zaburza również prawidłową funkcję metaboliczną karnityny. Karnityna (kwas L-3-hydroksy-4-N, N,N-trimetyloaminomasłowy) występuje w postaci dwóch stereoizomerów: D (prawoskrętna) i L (lewoskrętna) (ryc. 1).

Ryc. 1. Budowa L-karnityny i D-karnityny (wg [41] zmodyfikowano)
Postać D jest metabolicznie nieaktywna, natomiast L jest postacią fizjologiczną. L-karnityna występuje w postaci wolnej lub zestryfikowanej (związanej z grupami acylowymi różnej długości). Szczególną i główną rolą karnityny jest transport wolnych kwasów tluszczowych do macierzy mitochondrium w celu utlenienia ich w procesie β-oksydacji. Karnityna spełnia również inne funkcje:
• Przenosi produkty β-oksydacji z peroksysomów do mitochondriów, gdzie skrócony kwas tłuszczowy ulega dalszej β-oksydacji [35].
• Pełni rolę antyoksydanta zapobiegając akumulacji końcowych produktów peroksydacji lipidów [11,65].
• Działa chelatująco przyczyniając się do zmniejszenia stężenia żelaza w cytosolu komórek [5].
• Wpływa na stabilizację błon komórkowych (głównie erytrocytów) [31,33].
• Jest lekiem, który wykorzystuje się do leczenia: wrodzonych niedoborów karnityny, chorób układu krążenia, mięśni szkieletowych, nerek, wątroby, a także neurodegeneracyjnych chorób OUN [4,5,7,8,12,14,22,27,54,55,56,58]. Nałogowe spożywanie alkoholu prowadzi do funkcjonalnego niedoboru karnityny, co powoduje zahamowanie działania karnityny, mimo jej dostępności w diecie [2,3,50].
Synteza i metabolizm karnityny
Zapotrzebowanie dojrzałego organizmu na karnitynę jest pokrywane w 25% przez endogenną syntezę, a 75% dostarczane w pożywieniu [54]. W ciągu doby dorosły człowiek syntetyzuje 100-200 µmoli karnityny, a z dietą dostarczane jest 300-400 µmoli [31], czyli około 38 mg karnityny [43]. Najbogatszym źródłem karnityny dla osób dorosłych jest mięso czerwone, a dla niemowląt i dzieci mleko [54]. Karnityna przyjmowana w pożywieniu ulega całkowitej absorpcji w przewodzie pokarmowym w wyniku aktywnego transportu zależnego od sodu. W organizmie prekursorami i kofaktorami karnityny są przede wszystkim lizyna (dostarcza szkieletu węglowego L-karnityny) i metionina (dostarcza grup metylowych), a także kwas askorbinowy, niacyna, witamina B6 i jony Fe+2 [27]. Biosynteza karnityny w organizmie (ryc. 2) zapoczątkowana jest metylacją lizyny zawartej w takich białkach, jak: miozyna, aktyna czy histony.

Ryc. 2. Biosynteza karnityny (wg [45] zmodyfikowano)
Do grupy ε-aminowej lizyny przyłączane są 3 grupy metylowe z S-adenozylometioniny (SAM). Powstaje ε-N-trimetylolizyna, która rozpada się na glicynę i aldehyd γ-N-trimetyloaminomasłowy. Grupa aldehydowa aldehydu γ-N-trimetyloaminomasłowego jest utleniana do grupy karboksylowej, a węgiel β jest hydroksylowany. Powstaje kwas β-hydroksy-γ-N-trimetyloaminomasłowy, czyli karnityna [20]. Karnityna jest syntetyzowana endogennie w wątrobie, nerkach i mózgu, które mają pełny zestaw enzymów do syntezy L-karnityny [5]. Do pozostałych narządów karnityna dostarczana jest z krwią. Do wnętrza komórek L-karnityna trafia za pomocą transportu aktywnego.
Karnityna jest związkiem hydrofilnym o m.cz. 162 kDa [27]. Około 75% całkowitej zawartości karnityny u ludzi występuje w postaci wolnej, 15% w postaci acetylokarnityny, a pozostałe 10% w innych postaciach. W niewielkim procencie karnityna jest degradowana do metylocholiny, jednak w czasie głodu i przy diecie bogatotłuszczowej ilość wydalanej metylocholiny wzrasta. Nerki odgrywają główną rolę w homeostazie karnityny i jej estrów. Z moczem w ciągu doby wydalane jest 300-360 µmoli karnityny, z czego w postaci estrów wydala się prawie 50% karnityny [12]. Reabsorpcja kanalikowa wolnej karnityny wynosi 96-99% i jest większa niż jej estrów. Główną rolę w reabsorpcji karnityny odgrywa nerkowy transporter OCTN2 (sodium-dependent carnitine organic kation transporter) [12,24]. Próg nerkowy dla wolnej karnityny mieści się w zakresie jej fizjologicznego stężenia w osoczu i wynosi 45-65 µmoli/l. Zaburzony metabolizm L-karnityny występuje w wielu chorobach, m.in.: w niewydolności nerek [54], kwasicach organicznych, w cukrzycy [19], kardiomiopatii [55,64], a także w nadużywaniu alkoholu [2,3,16,21,34,47,48,50].
Rola karnityny w β-oksydacji kwasów tłuszczowych
L-karnityna odgrywa istotną rolę w metabolizmie długołańcuchowych kwasów tłuszczowych. Przenosi aktywowane (acylo-CoA) cząsteczki kwasów tłuszczowych z cytosolu do macierzy mitochondrialnej, gdzie ulegają utlenianiu w procesie oksydacji. W wyniku oksydacji kwasów tłuszczowych w mitochondriach powstaje energia w postaci adenozynotrifosforanu (ATP). Głównym źródłem kwasów tłuszczowych w komórce jest lipoliza triacylogliceroli i estrów cholesterolu oraz biosynteza kwasów tłuszczowych de novo. Błony mitochondrialne są nieprzepuszczalne dla długołańcuchowych acylo-CoA i niepolarnych cząsteczek CoA. Długołańcuchowe niepolarne kwasy tłuszczowe mogą przenikać przez wewnętrzną błonę mitochondrialną jedynie po połączeniu z polarną cząsteczką L-karnityny. Kwasy tłuszczowe ulegają aktywacji do acylo-CoA na zewnętrznej błonie mitochondrialnej, natomiast utlenianie kwasów tłuszczowych zachodzi w macierzy mitochondrium. Utlenianie kwasów tłuszczowych w mitochondrium jest procesem dwuetapowym. Pierwszy etap, czyli β-oksydacja polega na wielokrotnie powtarzanych reakcjach odwodornienia łańcucha węglowodorowego kwasu tłuszczowego przy węglu β i na rozpadzie utlenionego substratu na fragmenty dwuwęglowe, także przy kolejnych węglach β. Każdy cykl β-oksydacji powoduje skrócenie łańcucha kwasu tłuszczowego o kolejny fragment dwuwęglowy. Produktem β-oksydacji są cząsteczki acetylo~S-CoA. Podczas jednego cyklu b-oksydacji powstaje 5 cząsteczek ATP. Drugi etap utleniania kwasów tłuszczowych polega na utlenianiu reszt acetylowych w cyklu kwasów trikarboksylowych do CO2 i H2O, w czasie którego z jednej cząsteczki acetylo~S-CoA powstaje 12 cząsteczek ATP.
Kwasy tłuszczowe o krótkim łańcuchu zawierającym do 10 atomów węgla przenikają bezpośrednio przez wewnętrzną błonę do macierzy mitochondrium. Natomiast kwasy tłuszczowe o długich łańcuchach (>10 i więcej atomów węgla w cząsteczce) nie mogą wniknąć do mitochondrium bezpośrednio [33]. Kwasy tłuszczowe o długich łańcuchach, np. kwas palmitynowy muszą być aktywowane w cytosolu przez przyłączenie cytosolowego CoA-SH z udziałem tiokinazy (syntetazy acylo~S-CoA). Wewnętrzna błona mitochondrialna jest nieprzepuszczalna, ani dla CoA ani dla dużych niepolarnych cząsteczek, takich jak acylo~S-CoA, dlatego grupy acylowe muszą być przetransportowane do mitochondrium przez specjalny system transportu zwanym czółenkiem karnitynowym lub mostkiem karnitynowym (ryc. 3).

Ryc. 3. Mechanizm działania czółenka karnitynowego w mitochondriach wątroby: (1) Syntetaza acylo-CoA; (2) CPT I (palmitoilotransferaza karnitynowa I); (3) CACT (translokaza karnityna-acylokarnityna-przenośnik antyportowy); (4) CPT II (palmitoilotransferaza karnitynowa II); (5) CAT (acetylotransferaza karnitynowa) (wg [20] zmodyfikowano)
Przenośnikiem w tym systemie jest karnityna. Proces transportu długołańcuchowych kwasów z cytoplazmy do macierzy mitochondrium składa się z kilku etapów. Pierwszym etapem jest synteza acylo-CoA z kwasu tłuszczowego połączonego z CoA z udziałem syntetazy acylo-CoA (EC 6.2.1.3; tiokinazy), która katalizuje powstanie aktywnych kwasów tłuszczowych (Acylo-CoA). Syntetazy acylo-CoA znajdują się w siateczce śródplazmatycznej, w obrębie mitochondriów i na zewnętrznej błonie mitochondrialnej. Występuje kilka syntetaz (tiokinaz). Każda z nich jest swoista wobec kwasów tłuszczowych o określonej długości łańcucha [41]. Drugim etapem transportu długołańcuchowych kwasów tłuszczowych z cytoplazmy do macierzy mitochondrium jest synteza acylowych pochodnych karnityny. W cytosolu grupa acylowa zostaje przeniesiona z atomu siarki CoA na grupę hydroksylową karnityny z udziałem acylotransferaz karnitynowych np. acylotransferazy karnitynowej I (palmitoilotransferazy karnitynowej I, CPT I) (EC 2.3.1.21), znajdujących się po zewnętrznej stronie wewnętrznej błony mitochondrialnej (CPT I katalizuje powstanie acylowych pochodnych karnitynowych). Acylotransferazy karnitynowe różnią się między sobą umiejscowieniem narządowym i komórkowym [44] oraz swoistością w stosunku do reszt acylowych o różnej długości łańcucha węglowego. Wyróżnia się trzy grupy acylotransferaz karnitynowych [17,26,28,32,35,39,51]:
• acetylotransferazy karnitynowe (CrAT/CAT) swoiste w stosunku do krótkich łańcuchów węglowych (C2-C6),
• oktanylotransferazy (COT) swoiste w stosunku do pośredniej długości łańcucha węglowego (C6-C10),
• palmitoilotransferazy (CPT) swoiste w stosunku do długich łańcuchów węglowych (C14-C24).
Trzecim etapem transportu długołańcuchowych kwasów tłuszczowych z cytoplazmy do macierzy mitochondrium jest przeniesienie acylokarnityny przez wewnętrzną błonę mitochondrialną. Wymiana wolnej karnityny z jej acylową pochodną zachodzi z udziałem translokazy karnitynoacylokarnitynowej (carnitine-acylcarnitine-translocase – CACT), która działa jako błonowy wymienny przenośnik antyportowy karnityny [12].
W ostatnim etapie transportu kwasów tłuszczowych z cytoplazmy do macierzy mitochondrialnej, acylokarnityna reaguje z CoA znajdującym się w macierzy mitochondrium tworząc w macierzy mitochondrium acylo-CoA i wolną karnitynę. Ostatni etap transportu kwasów tłuszczowych z cytoplazmy do macierzy mitochondrium zachodzi z udziałem acylotransferazy karnitynowej II (palmitoilotransferazy karnitynowej II) (CPT II) (EC 2.3.2.21) umiejscowionej na wewnętrznej powierzchni wewnętrznej błony mitochondrialnej (ryc. 3) [39]. Wolna karnityna powraca do cytoplazmy z udziałem translokazy karnityna-acylokarnityna, poprzez wymianę z nową cząsteczką acylokarnityny. Natomiast powstały w macierzy mitochondrialnej acylo-CoA ulega β-oksydacji w wyniku czego zostaje wytworzony acetylo-CoA: wchodzący do cyklu kwasów trikarboksylowych [8], zużywany do wytwarzania ciał ketonowych lub acetylokarnityny. Badania z ostatnich lat wskazały na możliwość udziału również innych białek w przechodzeniu acylo-CoA do wnętrza mitochondrium. W błonie mitochondrialnej zidentyfikowano białkowe przenośniki kwasów tłuszczowych: FAT/CD36, FABPpm [13,25] oraz FATP-1 [52]. W macierzy mitochondriów znajduje się również acetylotransferaza karnitynowa (carnitine acetyltransferase – CAT), która przekształca krótko- i średniołańcuchowe kwasy tłuszczowe do pochodnych L-acylokarnityny z wykorzystaniem wewnątrzmitochondrialnej L-karnityny oraz katalizuje przenoszenie grup acetylowych z acetylo-CoA powstającego w β-oksydacji na L-karnitynę (ryc. 3) [26]. W reakcji katalizowanej przez acetylotransferazę karnitynową powstaje wolny CoA i acetylokarnityna, która jest przenoszona przez translokazę (CACT) do cytosolu. Reakcje katalizowane przez acetylotransferazę karnitynową i translokazę zapobiegają kumulacji w mitochondriach grup acylowych i acetylowych, które hamują proces β-oksydacji oraz regenerują wolny CoA.
Wpływ alkoholu etylowego na funkcje metaboliczne wątroby
Następstwem spożywania nadmiernej ilości alkoholu jest uszkodzenie narządów wewnętrznych, zwłaszcza wątroby. Uszkodzenie wątroby przez alkohol zależy od dobowego spożycia, indywidualnej wrażliwości, czasu nadużywania alkoholu, a także stanu odżywienia pacjenta. W wątrobie osób nadużywających alkoholu dochodzi kolejno do stłuszczenia, zapalenia, zwłóknienia, a w końcu do marskości. Prawdopodobieństwo alkoholowego zapalenia wątroby zwiększa się proporcjonalnie do ilości spożywanego etanolu i długości okresu nadużywania alkoholu. Metabolizm alkoholu etylowego zachodzi głównie w cytosolu i mitochondriach wątroby. Produktem utleniania alkoholu w cytosolu wątroby, katalizowanego przez dehydrogenazę alkoholową jest aldehyd octowy, który utlenia się do octanu. Utlenienie cząsteczki etanolu do octanu prowadzi do nadmiernego zużycia utlenionej postaci dinukleotydu nikotynamido-adeninowego (NAD+) i powstania nadmiaru zredukowanej postaci dinukleotydu nikotynamido-adeninowego (NADH), co prowadzi do zahamowania utleniania różnych substratów zależnych od NAD+, w tym β-oksydacji kwasów tłuszczowych. Octan ulega dalszej przemianie w mitochondrium. Przy nadużyciu alkoholu możliwości wątroby do utleniania octanu są ograniczone z powodu braku NAD+ i nadmiaru NADH. Nadmiar NADH po spożyciu alkoholu hamuje cykl kwasów trikarboksylowych i tworzenie ATP i GTP, bo nie ma NAD+ – akceptora kationów wodoru i elektronów powstałych z utleniania kwasów w cyklu kwasów trikarboksylowych [1]. Ze wzgledu na to, że NADH hamuje cykl Krebsa (a po etanolu jest go nadmiar), więc nie może być spalony octan, który przedostaje się do krążenia ogólnego. Przy braku NAD+ dochodzi również do skierowania acetylo~S-CoA do biosyntezy kwasów tłuszczowych lub ciał ketonowych, co prowadzi do ketonemii. Aldehyd octowy powstający podczas utleniania etanolu jest bardzo toksyczny dla mitochondrium wątroby [36]. Sam alkohol hamując katabolizm cholesterolu do kwasów żółciowych prowadzi do hipercholesterolemii. Hiperlipemia występująca w chronicznym alkoholizmie może być wynikiem hipertrofii retikulum endoplazmatycznego i aparatu Golgiego. Zwiększona biosynteza i zmniejszona β-oksydacja kwasów tłuszczowych prowadzi do ich kumulacji i w konsekwencji do zwiększonej biosyntezy triacylogliceroli, innych lipidów, cholesterolu i fosfolipidów, które prowadzą z czasem do stłuszczenia wątroby [30,46]. Zwiększony stosunek NADH/NAD+ powoduje również zwiększenie stosunku mleczan/pirogronian, czego wynikiem jest hiperlaktacydemia, która z kolei prowadzi do zmniejszonego wydalania kwasu moczowego przez nerki.
Główna droga przemiany alkoholu przebiega z udziałem dehydrogenazy alkoholowej, ale pewna część etanolu metabolizowana jest przez układ mikrosomalny zależny od cytochromu P-450 2E1 (CYP2E1), w którym uczestniczą także NADPH i O2 [36]. Podczas przewlekłego nadużywania alkoholu, metabolity utleniania etanolu przez układ mikrosomalny zależny od cytochromu (CYP2E1), mogą powodować uszkodzenie błon komórkowych i śmierć hepatocyta. Wzrosty stężenia NADH, a także kwasu mlekowego stymulują syntezę kolagenu w miofibroblastach. CYP2E1 indukowany jest nie tylko przez alkohol, ale też przez niektóre kwasy tłuszczowe i ketony, co prowadzi do generacji wolnych rodników [36], stresu oksydacyjnego z peroksydacją lipidów i uszkodzeniem błon komórkowych oraz z obniżeniem aktywności enzymów. Stres oksydacyjny i następcze uszkodzenie komórek wątrobowych prowadzące do zapalenia są spowodowane uwalnianiem wolnych rodników, cytokin zapalnych, czynnika martwicy nowotworów (TNF-α) oraz enzymów lizosomalnych w komórkach Kupffera [9,36,42,59,61,62]. Pod wpływem przewlekłego zatrucia alkoholem dochodzi nie tylko do ciężkich uszkodzeń wątroby, ale także uszkodzenia nerek, zwyrodnienia mięśnia sercowego i uszkodzenia gruczołów wydzielania wewnętrznego [15,29].
Nadużywanie alkoholu hamuje cykl Krebsa, a nagromadzone krótkołańcuchowe kwasy organiczne np. mlekowy, pirogronowy, propionowy, metylomalonowy, są wydzielane w większej ilości do moczu. Udowodniono, że zwiększona synteza m.in. kwasu propionowego i metylomalonowego związana jest ze zmniejszonym stężeniem karnityny w osoczu i zwiększonym wydalaniem acylokarnityn do moczu [10]. Wydaje się, że karnityna może odgrywać ochronną rolę, jako antagonista w procesie akumulacji wyżej wymienionych krótkołańcuchowych kwasów organicznych w wyniku nadmiernej intoksykacji alkoholem [10].
Karnityna w alkoholowej chorobie wątroby
Brak L-karnityny hamuje spalanie kwasów tłuszczowych. Z kolei nagromadzenie w wątrobie acylo-CoA może prowadzić do wrodzonej (sprzężonej z płcią) choroby cyklu mocznikowego – hiperamonemii [7,22,49]. Czasami objawy kliniczne, które towarzyszą niedoborowi L-karnityny w organizmie są podobne do zespołu Reye’a (encefalopatii wątrobowej). Niedobory wtórne L-karnityny w organizmie są związane z obniżoną ilością karnityny w diecie, zaburzeniami we wchłanianiu i transporcie, ze zwiększonym wydalaniem z moczem, chorobami wątroby i nerek, a także w wyniku zwiększonego zapotrzebowania metabolicznego na L-karnitynę (np. wskutek wzmożonego wysiłku fizycznego) [63]. Karnityna odgrywa niezwykle ważną rolę w wykorzystywaniu i metabolizowaniu kwasów tłuszczowych w wątrobie. Wykazano, że niedobór karnityny w wątrobie może się przyczyniać do stłuszczenia wątroby spowodowanego nałogowym spożywaniem alkoholu. Wykazano ponadto, że suplementacja karnityną w znacznym stopniu hamuje niewydolność wątroby (zapalenie, stłuszczenie) wywołane spożywaniem nadmiernej ilości alkoholu [6,9,57].
Alkoholowe stłuszczenie wątroby
Stłuszczenie wątroby jest pierwszą fazą alkoholowej choroby wątroby i jest stanem odwracalnym (zwykle w wyniku zaprzestania spożywania alkoholu). W rozdętych hepatocytach dotkniętych stłuszczeniem obserwuje się nagromadzenie triacylogliceroli. Stwierdza się upośledzone wydzielanie VLDL, natomiast aktywności transaminaz mogą być w normie. Wyniki badań, dotyczących stężenia karnityny w surowicy w poalkoholowym stłuszczeniu wątroby przeprowadzone przez różnych autorów, nie są jednoznaczne. De Sousa i wsp. [16] w surowicy, moczu i w wątrobie u alkoholików ze stłuszczeniem wątroby nie stwierdzili wyraźnej różnicy w stężeniu karnityny wolnej, całkowitej i acylokarnityny, w porównaniu do kontroli. Brak znamiennej różnicy w stężeniu karnityn, w porównaniu do kontroli, skłania do konkluzji, iż deficyt karnityny nie ma wpływu na stłuszczenie wątroby. De Sousa i wsp. u pacjentów nadużywających alkoholu stwierdzili natomiast bardzo wysokie stężenie karnityny wolnej i całkowitej w mięśniach.
Alonzo de la Pena i wsp. [2] u pacjentów z alkoholowym stłuszczeniem wątroby stwierdzili wzrost stężenia acylokarnityny głównie w osoczu, w porównaniu do grupy kontrolnej i uważają, że stężenie karnityny w osoczu zależy od stopnia alkoholowego uszkodzenia wątroby. Zdaniem Alonzo de la Pena i wsp. [2] hiperkarnitynemia w stłuszczeniu i marskości wątroby może być związana z przewlekłą niewydolnością i zmniejszonym klirensem nerkowym.
Badania eksperymentalne Sachana i wsp. [48] na szczurach karmionych dietą z etanolem wykazały niskie stężenia karnityny w osoczu, natomiast w wątrobie nie wykazano istotnego wzrostu stężenia karnityny wolnej, całkowitej i karnityny połączonej z długołańcuchowymi kwasami tłuszczowymi. Sachan i wsp. [48] udowodnili, iż podanie karnityny w diecie zapobiega stłuszczeniu wątroby.
Alkoholowa marskość wątroby
Alkoholowa marskość wątroby jest końcowym stadium alkoholowej choroby wątroby. Podobnie jak w alkoholowym stłuszczeniu wątroby badania dotyczące stężenia karnityny w alkoholowej marskości wątroby nie są jednoznaczne. Fuller i Hoppel [21] w surowicy pacjentów z alkoholową marskością wątroby stwierdzili istotnie podwyższone stężenie karnityny wolnej, całkowitej i połączonej z krótko- i długołańcuchowymi kwasami tłuszczowymi. Także Alonzo de la Pena i wsp. [2] u chorych z alkoholową marskością wątroby stwierdzili w osoczu ekstremalnie wysokie stężenie karnityny wolnej i acylokarnityny, w porównaniu do grupy kontrolnej. Według tych autorów [2] w chorobach wątroby, a zwłaszcza w marskości, dochodzi również do niewydolności nerek i do spadku klirensu karnityny, co w konsekwencji prowadzi do wzrostu stężenia karnityny w osoczu. Natomiast Rudman i wsp. [47] stwierdzili u pacjentów z alkoholową marskością wątroby hipokarnitynemię w osoczu i tkankach, tłumacząc to brakiem prekursorów w pożywieniu dla endogennej syntezy karnityny, a także zablokowaniem jej syntezy przez uszkodzenie komórek wątroby. Natomiast Sakvarelidze [50] stwierdził, że stężenie karnityny zależy od czasu intoksykacji alkoholem. Według niego im dłuższa ekspozycja organizmu na alkohol tym coraz mniejsze stężenia L-karnityny w osoczu.
Ciekawych spostrzeżeń w swojej pracy dostarczyli Amodio i wsp. [3]. Stwierdzili oni w surowicy pacjentów z marskością wątroby (niezależnie od etiologii marskości wątroby) wysokie stężenia: acetylokarnityny, krótkołańcuchowej acylokarnityny, karnityny zestryfikowanej i całkowitej karnityny. Jednakże najwyższe stężenie acetylokarnityny Amodio i wsp. [3] stwierdzili u nałogowych alkoholików. Według nich alkohol jest czynnikiem zaostrzającym (pogorszającym) funkcję wątroby i przyspieszającym marskość wątroby.
Krähenbűhl i Reichen [34] stwierdzili u pacjentów z alkoholową marskością wątroby wzrost stężenia karnityny całkowitej oraz karnityny połączonej z krótko- i długołańcuchowymi kwasami tłuszczowymi. Według nich zwiększone stężenie karnityny w alkoholowej marskości wątroby związane jest ze zwiększoną biosyntezą karnityny z powodu zwiększonego metabolizmu białek w mięśniach szkieletowych.
Alkoholowa przewlekła niewydolność wątroby
W swoich badaniach Alonso de la Pena i wsp. [2] wykazali u alkoholików bez marskości wątroby, ale z przewlekłą niewydolnością wątroby bez nadciśnienia wrotnego, niskie stężenie karnityny, co może być według nich spowodowane niedożywieniem i zmniejszoną endogenną syntezą karnityny.
Natomiast u pacjentów z przewlekłą niewydolnością wątroby bez marskości wątroby Krähenbűhl i Reichen [34] nie stwierdzili zmian w stężeniu karnityny w surowicy, ale chorzy z marskością wątroby o innej etiologii mieli o 29% wyższe stężenie długołańcuchowej acylkarnityny.
Karnityna w chorobach wątroby o etiologii niezwiązanej ze spożyciem alkoholu
Zaburzenia metabolizmu karnityny stwierdza się również w chorobach wątroby o etiologii innej niż alkoholowa. Selimoglu i wsp. [53] u niedożywionych dzieci i u dzieci z atrezją dróg żółciowych oraz marskością kryptogenną, stwierdzili duże stężenie karnityny zarówno w surowicy jak i w wątrobie. Natomiast u dzieci z chorobą Wilsona stężenia karnityny w surowicy i w wątrobie są niewielkie.
Terapeutyczne działanie suplementacji karnityną w niewydolności wątroby wywołanej toksycznym działaniem alkoholu
Doniesiono, że podawanie L-karnityny lub acetylokarnityny zmniejsza metaboliczne zaburzenia wywołane chronicznym nadużywaniem alkoholu [11]. Doświadczalne podanie karnityny szczurom ze stłuszczeniem wątroby zapobiegało destrukcji mitochondriów i zmniejszało wypływ aminotransferazy alaninowej (ALT) i dehydrogenazy mleczanowej (LDH) na zewnątrz komórki [57]. Bertelli i wsp. [6] wykazali, że podawanie L-karnityny i koenzymu Q10 zapobiega naciekaniu lipidami wątroby u szczurów narażonych na zatrucie etanolem.
Badania Bykova i wsp. [9] wykazały pozytywny wpływ suplementacji karnityną na funkcję wątroby u badanych szczurów. Stwierdzili oni zmniejszone o 44% stłuszczenie wątroby oraz zmniejszone zapalenie wątroby o 41%. Przed podaniem karnityny stwierdzili [9] w wątrobie nagromadzenie triacylogliceroli (TAG) 4 razy większe w stosunku do kontroli, natomiast po podaniu karnityny TAG wzrósł tylko 1,7 razy w stosunku do kontroli. Bykov i wsp. [9] wykazali hepatoprotekcyjny wpływ karnityny na komórki Kupffera oraz zmniejszenie przez komórki Kupffera syntezy czynnika martwicy guza (tumor necrosis factor – TNF-α). Również badania Rhewa i Sachana [46] potwierdzają pozytywny wpływ suplementacji karnityną na funkcję wątroby.
Sachan i wsp. [48] w badaniach doświadczalnych stwierdzili, że karnityna zapobiega kumulacji lipidów w wątrobie i przeciwdziała stłuszczeniu wątroby u szczurów. Tempesta i wsp. [56] wykazali korzystny wpływ karnityny na funkcje neuropsychologiczne u alkoholików w trakcie abstynencji. Stwierdzili, że acetylo-L-karnityna może być pożytecznym i bezpiecznym czynnikiem terapeutycznym dobrze wpływającym na funkcje kognitywne, które są zaburzone u osób przewlekle zatrutych alkoholem. U pacjentów z encefalopatią wątrobową w marskości wątroby (z powodu alkoholizmu, hepatitis typu B i C oraz marskości kryptogennej). Malaguarnera i wsp. [38] wykazali pozytywne wyniki leczenia L-karnityną, w postaci zmniejszenia stężenia amoniaku w wątrobie (L-karnityna zwiększa wydajność cyklu mocznikowego, w którym toksyczny amoniak ulega wzmożonej przemianie do mocznika, ponieważ L-karnityna działa podobnie jak ornityna i arginina – aminokwasy związane z cyklem mocznikowym).
Podsumowanie
Z przeglądu piśmiennictwa wynika, że wyniki badań dotyczących stężenia karnityny w surowicy (osoczu) i tkankach w alkoholowej marskości wątroby, stłuszczeniu wątroby i w chorobach wątroby o innej etiologii, przeprowadzonych przez wielu autorów, nie są jednoznaczne (tab. 1).
Tabela 1. Zmiana stężenia karnityny w surowicy (osoczu) oraz w tkankach u pacjentów w przewlekłym zatruciu alkoholem oraz u pacjentów niezwiązanych ze spożywaniem alkoholu
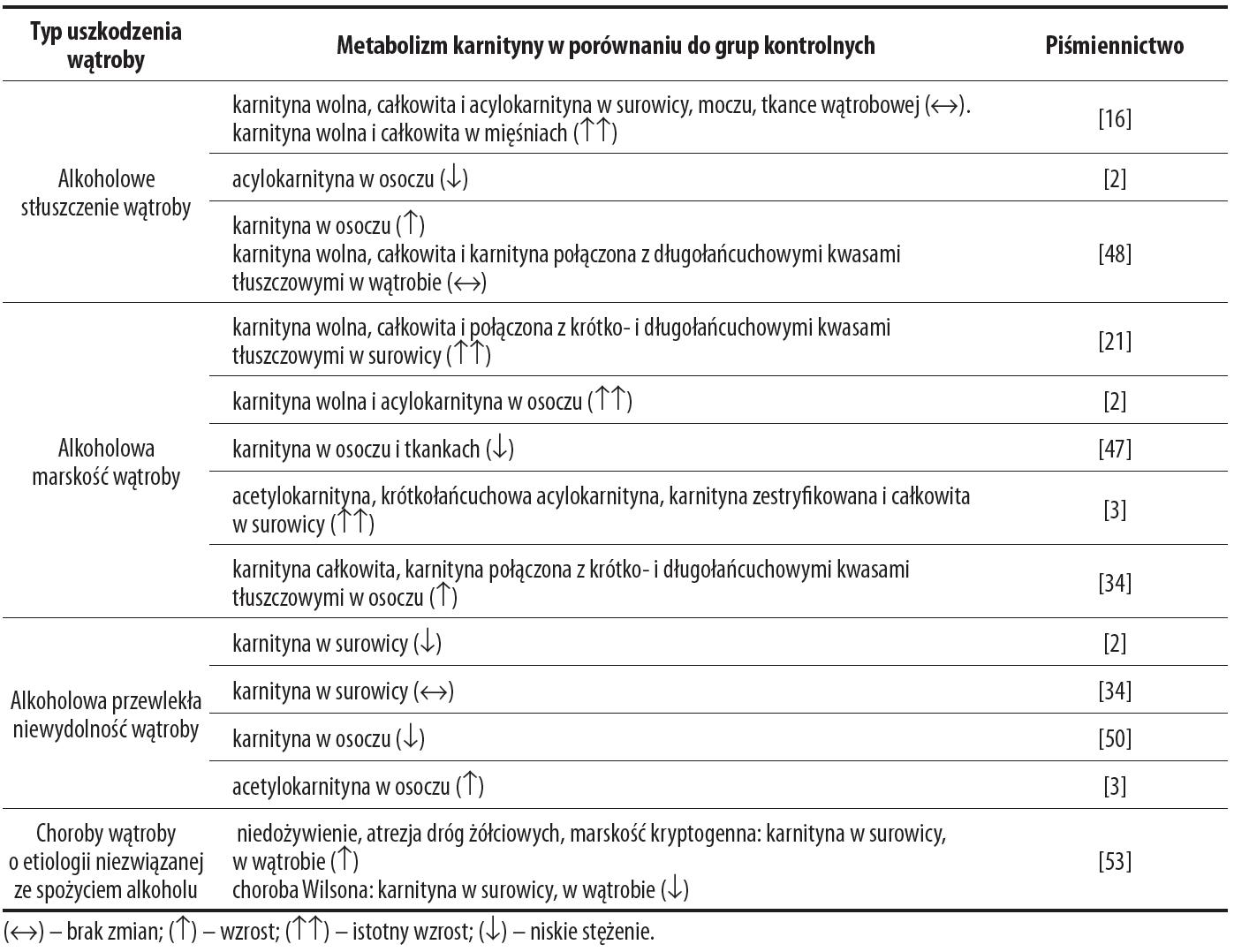
Niejednoznaczność wyników oznaczania karnityny wolnej i związanej z kwasami tłuszczowymi w surowicy (osoczu) i tkankach zależy od wielu czynników m.in. czasu ekspozycji organizmu na alkohol, etiologii chorób wątroby, stanu odżywiania oraz współistnienia innych chorób, a zwłaszcza niewydolności nerek. W alkoholowym stłuszczeniu wątroby Alonso de la Pena i wsp. [2] stwierdzili głównie w osoczu wzrost stężenia tylko acylokarnityny, natomiast De Sousa i wsp. [16] w surowicy, moczu i w wątrobie u alkoholików ze stłuszczeniem wątroby nie stwierdzili wyraźnej różnicy w stężeniu karnityny wolnej, całkowitej i acylkarnityny, w porównaniu do kontroli. Również inni autorzy [16] u alkoholików nie stwierdzili różnic w stężeniu karnityny wolnej i związanej z kwasami tłuszczowymi w surowicy, moczu i wątrobie w stosunku do kontroli, ale stwierdzili bardzo wysokie stężenie karnityny w mięśniach [16]. Podobne wyniki uzyskano w alkoholowej marskości wątroby. Wzrost stężenia karnityny opisali jedni autorzy [2,3,21,34], natomiast niskie stężenia karnityny uzyskali drudzy autorzy [47,50]. Również podobne wyniki uzyskano u alkoholików w przewlekłej niewydolności wątroby. Alonso de la Pena i wsp. [2] stwierdzili niskie stężenia karnityny w osoczu, natomiast Krähenbűhl i Reichen [34] nie zaobserwowali zmian w stężeniu karnityny w surowicy uzależnionych od alkoholu.
Rozbieżności w wynikach badań mogą pochodzić także z braku jednolitych standardów przeprowadzonych badań. Wyjściowe (pierwszorazowe) oznaczenia stężenia karnityny w surowicy (osoczu) przeprowadzane były u pacjentów będących w różnym okresie abstynencji (od doby do 2-3 tygodni), poza tym nie uwzględniano współistniejących innych chorób, głównie niewydolności nerek, które odgrywają znaczącą rolę w metabolizmie karnityny.
L-karnityna, acetylo-L-karnityna i propionylo-L-karnityna są stosowane w chorobach układu krążenia [4] i neurodegeneracyjnych chorobach ośrodkowego układ nerwowego[14]. L-karnitynę jako lek wspomagający stosuje się również w cukrzycy [40], u pacjentów dializowanych [23], w kuracjach odchudzających, ale także w okresie wytężonej pracy fizycznej i umysłowej [58]. Suplementacja L-karnityną osób narażonych na chroniczne nadużywanie alkoholu zmniejsza metaboliczne zaburzenia spowodowane toksycznym działaniem alkoholu. Z przeglądu literatury wynika, że podawanie karnityny w diecie zapobiega kumulacji lipidów w wątrobie i przeciwdziała stłuszczeniu wątroby [9,18,48].
Po przeanalizowaniu rozbieżności dotyczących stężenia karnityny w surowicy (osoczu) i tkankach osób narażonych na przewlekłe nadużywanie alkoholu, a także u innych chorych z niewydolnością wątroby o innej etiologii niż alkoholowa, wydaje się że suplementacja karnityną przeciwdziała toksycznym efektom alkoholu na metabolizm komórek wątroby [18,37].
PIŚMIENNICTWO
[1] Adamo S., Siliprandi N., Dl Lisa F., Carrara M., Azzurro M., Sartori G., Vita G., Ghidini O.: Effect of L-carnitine on ethanol and acetate plasma levels after oral administration of ethanol in humans. Alcohol. Clin. Exp. Res., 1988; 12: 653-654
[PubMed]
[2] Alonso de la Pena C., Rozas I., Alvares-Prechous A., Pardinas M.C., Paz J.M., Rodriguez-Segade S.: Free carnitine and acylcarnitine levels in sera of alcoholics. Biochem. Med. Metab. Biol., 1990; 44: 77-83
[PubMed]
[3] Amodio P., Angeli P., Merkel C., Menon F., Gatta A.: Plasma carnitine levels in liver cirrhosis: relationship to nutritional status and liver damage. J. Clin. Chem. Clin. Biochem., 1990; 28: 619-626
[PubMed]
[4] Arsenian M.A.: Carnitine and its derivatives in cardiovascular disease. Prog. Cardiovasc. Dis., 1997; 40: 265-286
[PubMed]
[5] Aydogdu N., Atmaca G., Yalcin O., Taskiran R., Tastekin E., Kaymak K.: Protective effects of L-carnitine on myoglobinuric acute renal failure in rats. Clin. Exp. Pharmacol. Physiol., 2006; 33: 119-124
[PubMed]
[6] Bertelli A., Cerrati A., Giovannini L., Mian M., Spaggiari P., Bertelli A.A.: Protective action of L-carnitine and coenzyme Q10 against hepatic trigliceride infiltration induced by hyperbaric oxygen and ethanol. Drugs Exp. Clin. Res., 1993; 19: 65-68
[PubMed]
[7] Bonomini M., Sirolli V., Dottori S., Amoroso L., Di Liberato L., Arduini A.: L-carnitine inhibits a subset of platelet activation responses in chronic uraemia. Nephrol. Dial. Transplant., 2007; 22: 2623-2629
[PubMed] [Full Text HTML] [Full Text PDF]
[8] Brass E.P., Hiatt W.R.: The role of carnitine and carnitine supplementation during exercise in man and individuals with special needs. J. Am. Coll. Nutr., 1998; 17: 207-215
[PubMed] [Full Text HTML] [Full Text PDF]
[9] Bykov I., Järveläinen H., Lindros K.: L-carnitine alleviates alcohol-induced liver damage in rats: role of tumour necrosis factor-alpha. Alcohol Alcohol., 2003; 38: 400-406
[PubMed] [Full Text HTML] [Full Text PDF]
[10] Calabrese V., Calvani M., Butterfield D.A.: Increased formation of short-chain organic acids after chronic ethanol administration and its interaction with the carnitine pool in rat. Arch. Biochem. Biophys., 2004; 43: 271-278
[PubMed]
[11] Calabrese V., Scapagnini G., Latteri S., Colombrita C., Ravagna A., Catalano C., Pennisi G., Calvani M., Butterfield D.A.: Long-term ethanol administration enhances age-dependent modulation of redox state in different brain regions in the rat: protection by acetyl carnitine. Int. J. Tissue React., 2002; 24: 97-104
[PubMed]
[12] Calvani M., Benatti P., Mancinelli A., D’Iddio S., Giordano V., Koverech A., Amato A., Brass E.P.: Carnitine replacement in end-stage renal disease and hemodialysis. Ann. NY Acad. Sci., 2004; 1033: 52-66
[PubMed]
[13] Campbell S.E., Tandon N.N., Woldegiorgis G., Luiken J.J., Glatz J.F., Bonen A.: A novel function for fatty acid translocase (FAT)/CD36: involvement in long chain fatty acid transfer into the mitochondria. J. Biol. Chem., 2004; 279: 36235-36241
[PubMed] [Full Text HTML] [Full Text PDF]
[14] Carta A., Calvani M., Bravi D., Bhuachalla S.N.: Acetyl-L-carnitine and Alzheimer’s disease: pharmacological considerations beyond the cholinergic sphere. Ann.N.Y. Acad. Sci., 1993; 695: 324-326
[PubMed]
[15] Cylwik B., Daniluk M., Chrostek L., Szmitkowski M.: Wpływ zasobów żelaza na wskaźniki nadużywania alkoholu i alkoholowego uszkodzenia wątroby. Pol. Merkur. Lek., 2010; 28: 450-453
[PubMed]
[16] De Sousa C., Leung N.W., Chalmers R.A., Peters T.J.: Free and total carnitine and acylcarnitine content of plasma, urine, liver and muscle of alcoholics. Clin. Sci., 1988; 75: 437-440
[PubMed]
[17] Duranti G., Boenzi S., Rizzo C., Rav? L., Di Ciommo V., Carrozzo R., Meschini M.C., Johnson D.W., Dionisi-Vici C.: Urine acylcarnitine analysis by ESI-MS/MS: a new tool for the diagnosis of peroxisomal biogenesis disorders. Clin. Chim. Acta, 2008; 398: 86-89
[PubMed]
[18] Eaton S., Zaitoun A.M., Record C.O., Bartlett K.: Beta-oxidation in human alcoholic and non-alcoholic hepatic steatosis. Clin. Sci., 1996; 90: 307-313
[PubMed]
[19] Fan J.P., Kim D., Kawachi H., Ha T.S., Han G.D.: Ameliorating effects of L-carnitine on diabetic podocyte injury. J. Med. Food., 2010; 13: 1324-1330
[PubMed]
[20] Flanagan J.L., Simmons P.A., Vehige J., Willcox M.D., Garrett Q.: Role of carnitine in disease. Nutr. Metab., 2010; 7: 30-44
[PubMed] [Full Text HTML] [Full Text PDF]
[21] Fuller R.K., Hoppel C.L.: Elvated plasma carnitine in hepatic cirrhosis. Hepatology, 1983; 3: 554-558
[PubMed]
[22] Guarnieri G., Biolo G., Vinci P., Massolino B., Barazzoni R.: Advances in carnitine in chronic uremia. J. Ren. Nutr., 2007; 17: 23-29
[PubMed]
[23] Hakeshzadeh F., Tabibi H., Ahmadinejad M., Malakoutian T., Hedayati M.: Effects of L-Carnitine supplement on plasma coagulation and anticoagulation factors in hemodialysis patients. Ren. Fail., 2010; 32: 1109-1114
[PubMed]
[24] Hedayati S.S.: Dialysis-related carnitine disorder. Semin. Dial., 2006; 19: 323-328
[PubMed]
[25] Holloway G.P., Lally J., Nickerson J.G., Alkhateeb H., Snook L.A., Heigenhauser G.J., Calles-Escandon J., Glatz J.F., Luiken J.J., Spriet L.L., Bonen A.: Fatty acid binding protein facilitates sarcolemmal fatty acid transport but not mitochondrial oxidation in rat and human skeletal muscle. J. Physiol., 2007; 582: 393-405
[PubMed] [Full Text HTML] [Full Text PDF]
[26] Hsiao Y.S., Jogl G., Tong L.: Crystal structures of murine carnitine acetyltransferase in ternary complexes with its substrates. J. Biol. Chem., 2006; 281: 28480-28487
[PubMed] [Full Text HTML] [Full Text PDF]
[27] Hurot J.M., Cucherat M., Haugh M., Fouque D.: Effects of L-carnitine supplementation in maintenance hemodialysis patients: a systematic review. J. Am. Soc. Nephrol., 2002; 13: 708-714
[PubMed] [Full Text HTML] [Full Text PDF]
[28] Jaudzems K., Kuka J., Gutsaits A., Zinovjevs K., Kalvinsh I., Liepinsh E., Liepinsh E., Dambrova M.: Inhibition of carnitine acetyltransferase by mildronate, a regulator of energy metabolism. J. Enzyme Inhib. Med. Chem., 2009; 24: 1269-1275
[PubMed]
[29] Jelski W., Kutyłowska E., Szmitkowski M.: Rola alkoholu etylowego w patogenezie zapaleń trzustki. Pol. Merkur. Lek., 2011; 30: 66-68
[PubMed]
[30] Jeong W.I., Osei-Hyiaman D., Park O., Liu J., Bátkai S., Mukhopadhyay P., Horiguchi N., Harvey-White J., Marsicano G., Lutz B., Gao B., Kunos G.: Paracrine activation of hepatic CB1 receptors by stellate cell-derived endocannabinoids mediates alcoholic fatty liver. Cell. Metab., 2008; 7: 227-235
[PubMed] [Full Text HTML] [Full Text PDF]
[31] Kadiroglu A.K., Yilmaz M.E., Sit D., Kara I.H., Isikoglu B.: The evaluation of postdialysis L-carnitine administration and its effect on weekly requiring doses of rHuEPO in hemodialysis patients. Ren. Fail., 2005; 27: 367-372
[PubMed]
[32] Kerner J., Hoppel C.: Fatty acid import into mitochondria. Biochim. Biophys. Acta, 2000; 1486: 1-17
[PubMed]
[33] Kopple J.D., Ding H., Letoha A., Ivanyi B., Qing D.P., Dux L., Wang H.Y., Sonkodi S.: L-carnitine ameliorates gentamicin-induced renal injury in rats. Nephrol. Dial. Transplant., 2002; 17: 2122-2131
[PubMed] [Full Text HTML] [Full Text PDF]
[34] Krähenbűhl S., Reichen J.: Carnitine metabolizm in patients with chronic liver disease. Hepatology, 1997; 25: 148-153
[PubMed]
[35] Kunau W.H., Dommes V., Schulz H.: β-oxidation of fatty acids in mitochondria, peroxisomes, and bacteria: a century of continued progress. Prog. Lipid. Res., 1995; 34: 267-342
[PubMed]
[36] Lieber C.S.: Alcoholic fatty liver: its pathogenesis and mechanism of progression to inflammation and fibrosis. Alcohol, 2004; 34: 9-19
[PubMed]
[37] Łapiński T.W., Grzeszczuk A.: Wpływ karnityny na stężenie amoniaku i metabolizm lipidów u chorych na marskość wątroby. Pol. Merkur. Lek., 2003; 15: 38-41
[PubMed]
[38] Malaguarnera M., Pistone G., Elvira R., Leotta C., Scarpello L., Liborio R.: Effect of L-carnitine in patients with hepatic encephalopathy. World J. Gastrolenterol., 2005; 11: 7197-7202
[PubMed] [Full Text HTML] [Full Text PDF]
[39] McGarry J.D., Brown N.F.: The mitochondrial carnitine palmitylotransferase system. From concept to molecular analysis. Eur. J. Biochem., 1997; 244: 1-14
[PubMed] [Full Text HTML] [Full Text PDF]
[40] Mingorance C., Rodríguez-Rodríguez R., Justo M.L., Alvarez de Sotomayor M., Herrera M.D.: Critical update for the clinical use of L-carnitine analogs in cardiometabolic disorders. Vasc. Health Risk Manag., 2011; 7: 169-176
[PubMed] [Full Text HTML] [Full Text PDF]
[41] Murray R., Granner D.K., Mayes P.A., Rodwell V.W.: Harper’s Biochemistry twenty-second edition, Copyright for the Polish Edition by PZWL, Warszawa, 1995; 24: 260-273
[42] Nagata K., Suzuki H., Sakaguchi S.: Common pathogenic mechanism in development progression of liver injury caused by non-alcoholic or alcoholic steatohepatitis. J. Toxicol. Sci., 2007; 32: 453-468
[PubMed] [Full Text PDF]
[43] Paik W.K., Kim S.: Protein methylation, chemical, enzymological and biological significance. Adv. Enzymol. Relat. Areas Mol. Biol., 1975; 42: 227-286
[PubMed]
[44] Ramsay R.R., Gandour R.D., van der Leij F.R.: Molecular enzymology of carnitine transfer and transport. Biochim. Biophys. Acta, 2001; 1546: 21-43
[PubMed]
[45] Rebouche C. J.: Carnitine function and requirements during the life cycle. FASEB J., 1992; 6: 3379-3386
[PubMed] [Full Text PDF]
[46] Rhew T.H., Sachan D.S.: Dose-dependent lipotropic effect of carnitine in chronic alcoholic rats. J. Nutr., 1986; 116: 2263-2269
[PubMed] [Full Text PDF]
[47] Rudman D., Sewell C., Ansley J.D.: Deficiency of carnitine in cachectic cirrhotic patients. J. Clin. Invest., 1977; 60: 716-723
[PubMed] [Full Text HTML] [Full Text PDF]
[48] Sachan D.S., Rhew T.H, Ruark R.A.: Ameliorating effect of carnitine and its precursors on alcohol-induced fatty liver. Am. J. Clin. Nut., 1984; 39: 738-744
[PubMed] [Full Text PDF]
[49] Saheki T.: Carnitine as a vitamin-like biofactor. Nihon Rinsho, 1999; 57: 2270-2275
[PubMed]
[50] Sakvarelidze E.P.: Change of concentration of L-carnitine in blood and other tissues in rats on a background of the alcohol intake and influence of mildronate on its level. Georgian Med. News, 2006; 137: 94-96
[PubMed]
[51] Schmalix W., Bandlow W.: The ethanol-inducible YAT1 gene from yeast encodes a presumptive mitochondrial outer carnitine acetyltransferase. J. Biol. Chem., 1993; 268: 27428-27439
[PubMed] [Full Text PDF]
[52] Sebastián D., Guitart M., García-Martínez C., Mauvezin C., Orellana-Gavalda J.M., Serra D., Gómez-Foix A.M., Hegardt F.G., Asins G.: Novel role of FATP1 in mitochondrial fatty acid oxidation in skeletal muscle cells. J. Lipid Res., 2009; 50: 1789-1799
[PubMed] [Full Text HTML] [Full Text PDF]
[53] Selimoglu M.A., Aydogdu S., Yagci R.V., Huseyinov A.: Plasma and liver carnitine status of children with chronic liver disease and cirrhosis. Pediatr. Int., 2001; 43: 391-395
[PubMed]
[54] Steiber A.L., Weatherspoon L.J., Spry L., Davis A.T.: Serum carnitine concentrations correlated to clinical outcome in chronic hemodialisis patients. Clin. Nutr., 2004; 23: 27-34
[PubMed]
[55] Strauss M., Anselmi G., Hermoso T., Tejero F.: Carnitine promotes heat shock protein synthesis in adriamycin-induced cardiomyopathy in a neonatal rat experimental model. J. Mol. Cell. Cardiol., 1998; 30: 2319-2325
[PubMed]
[56] Tempesta E., Troncon R., Janiri L., Colusso L., Riscica P., Saraceni G., Gesmundo E., Calvani M., Benedetti N., Pola P.: Role of acetyl-L-carnitine in the treatment of cognitive deficit in chronic alcoholism. Int. J. Clin. Pharmacol. Res., 1990; 10: 101-107
[PubMed]
[57] Tolba R.H., Pűtz U., Decker D., Dombrowski F., Lauschke H.: L-carnitine ameliorates abnormal vulnerability of steatotic rat livers to cold ischemic preservation. Transpl., 2003; 76: 1681-1686
[PubMed]
[58] Wanic-Kossowska M., Kaźmierski M., Pawliczak E., Kobelski M.: Skojarzone leczenie niedokrwistości L-karnityną i erytropoetyną u chorych na przewlekłą niewydolność nerek poddanych hemodializoterapii. Pol. Arch. Med. Wew., 2007; 117: 14-19
[PubMed] [Full Text PDF]
[59] Waszkiewicz N., Szajda S.D., Jankowska A., Kepka A., Dobryniewski J., Szulc A., Zwierz K.: The effect of the binge drinking session on the activity of salivary, serum and urinary beta-hexosaminidase: preliminary data. Alcohol. Alcohol., 2008; 43: 446-450
[PubMed] [Full Text HTML] [Full Text PDF]
[60] Waszkiewicz N., Szajda S.D., Kępka A., Zalewska A., Konarzewska B., Szulc A., Chojnowska S., Zwierz-Gugała D.Z., Zwierz K.: Hepatotoksyczność „binge drinking”. Med. Sci. Rev. Hepatol., 2009; 9: 106-112
[61] Waszkiewicz N., Szajda S.D., Konarzewska B., Szulc A., Kepka A., Zwierz K.: Underappreciated role of binge drinking in the risk of lung cancer. Eur. J. Public. Health, 2010; 20: 6
[PubMed] [Full Text HTML] [Full Text PDF]
[62] Waszkiewicz N., Szajda S.D., Zalewska A., Konarzewska B., Szulc A., Kepka A., Zwierz K.: Binge drinking-induced liver injury. Hepatology, 2009; 50: 1676
[PubMed]
[63] Winter S., Opala G.: Wtórny niedobór karnityny – przyczyny, objawy kliniczne i leczenie. Post. Ped., 1990; 11-12: 99-103
[64] Zeidán Q., Strauss M., Porras N., Anselmi G.: Differential long-term subcellular responses in heart and liver to adriamycin stress. Exogenous L-carnitine cardiac and hepatic protection. J. Submicrosc. Cytol. Pathol., 2002; 34: 315-321
[PubMed]
[65] Zou W., Noh S.K., Owen K.Q., Koo S.I.: Dietary L-carnitine enhances the lymphatic absorption of fat and α-tocopherol in ovariectomized rats. J. Nutr., 2005; 135: 753-756
[PubMed] [Full Text HTML] [Full Text PDF]
Autorzy deklarują brak potencjalnych konfliktów interesów.