
Rola apoptosomu w aktywacji prokaspazy 9
Łukasz Marek 1Streszczenie
Apoptoza, zwana też programowaną śmiercią, jest fizjologicznym procesem powodującym wiele zmian zarówno morfologicznych jak i biochemicznych, zachodzących w komórce i prowadzących do jej śmierci. Wraz z procesami proliferacji, różnicowania i dojrzewania odpowiedzialna jest za kontrolę liczby i rodzaju komórek, usuwając zbędne lub niebezpieczne dla organizmu. Najważniejszymi enzymami biorącymi udział w apoptozie są kaspazy, które hydrolizują białka strukturalne i funkcjonalne, prowadząc ostatecznie do śmierci komórki. Kaspazy syntetyzowane są w komórce jako nieaktywne zymogeny i muszą ulec aktywacji, aby mogły pełnić swoje funkcje. Do aktywacji kaspaz prowadzą dwa szlaki: zewnętrzny związany z receptorami błonowymi i ich ligandami oraz wewnętrzny zależny od mitochondriów. Centralnym elementem w szlaku mitochondrialnym jest specjalny białkowy kompleks, zwany apoptosomem, który umożliwia i ułatwia aktywację prokaspazy 9. Do powstania apoptosomu potrzebne jest białko Apaf-1, cytochrom c i dATP/ATP. Aktywne apoptosomy powstają tylko w odpowiedzi na działanie czynników apoptotycznych. Istnieje wiele czynników regulujących tworzenie apoptosomu w fizjologicznych warunkach. Wykazano, że zaburzenia tworzenia apoptosomu mają istotne znaczenie w patogenezie nowotworów i występowaniu ich oporności na chemioterapię. Dlatego też trwają badania nad znalezieniem związków, które będą mogły indukować lub hamować powstawanie apoptosomu, a to będzie miało duże znaczenie w przypadku leczenia chorych na nowotwory i choroby wynikające z nadmiernie zachodzącego procesu apoptozy. W pracy omówiono budowę i mechanizm powstawania apoptosomu, jego regulacji oraz wykorzystania jako potencjalnego celu terapeutycznego.
Skoncentrowano się na ogólnej charakterystyce apoptosomu, jego roli i mechanizmie powstawania. Poza tym przedstawiono regulację jego aktywności i zwrócono uwagę na rolę zaburzeń aktywności apoptosomu w karcynogenezie i chemiooporności. Szczególną uwagę poświęcono związkom zdolnym wpływać na tworzenie i aktywację apoptosomu, które mogą stanowić w przyszłości ciekawe narzędzia terapeutyczne.
Słowa kluczowe: apoptoza • mitochondrium • kaspazy • apoptosame • Apaf-1
Summary
Apoptosis, also called programmed cell death, is a physiological process that causes a number of morphological and biochemical changes, occurring in the cell and leading to its death. Along with the processes of proliferation, differentiation and maturation, it is responsible for controlling the amount and types of cells, removing those that are unnecessary or dangerous for the organism.
The most important enzymes involved in apoptosis are caspases, which hydrolyze the structural and functional proteins, ultimately leading to cell death. The caspases are synthesized in the cell as inactive zymogens and have to be activated in order to perform their functions. Two pathways lead to the activation of caspases: the extrinsic pathway associated with membrane receptors and their ligands, and the intrinsic pathway dependent on mitochondria. The central element in the mitochondrial pathway is a special protein complex, the apoptosome, which enables and facilitates the activation of procaspase 9. Apaf-1, cytochrome c and dATP/ATP are needed to form the apoptosome. Active apoptosomes arise only in response to apoptotic agents. There are many factors regulating apoptosome formation in physiological conditions. It was shown that disorders in apoptosome formation have great importance in the pathogenesis of cancer and the occurrence of resistance to chemotherapy. Therefore, research continues on finding compounds which are able to induce or inhibit the formation of the apoptosome. The results of this research will have great importance for the treatment of cancers or diseases resulting from an excessively prolonged process of apoptosis. In this paper the general characteristics of the apoptosome, its role and mechanism of formation are presented. Additional information is given on the regulation of its activity and on the role of disturbance of apoptosome activity in cancerogenesis and chemoresistance. Particular attention is paid to those compounds that are able to influence the formation and activation of the apoptosome, and may in future be interesting therapeutic tools.
Key words:apoptosis • mitochondrion • caspases • apoptosome • Apaf-1
Wykaz skrótów:
Apaf-1 – czynnik 1 aktywujący proteazy apoptotyczne (apoptotic protease-activating factor 1); Bcl-2 – rodzina białek proapoptotycznych i antyapoptotycznych (B-cell lymphoma/leukemia-2); B-CLL – przewlekła białaczka limfocytowa B-komórkowa (B-cell chronic lymphocytic leukemia); BIR – funkcjonalna jednostka białek IAP (baculoviral IAP repeat); CARD – domena aktywacji i rekrutacji kaspaz (caspase activation and recruitment domain); DD – domena śmierci (death domain); DED – efektorowa domena śmierci (death effector domain); DISC – degradosom, kompleks sygnałowy, w którym aktywowana jest prokaspaza 8 (death inducing signalling complex); FADD – białko adaptorowe łączące się z receptorem Fas (Fas-associated death domain containing protein); HD1 – domena helikalna 1 (small helical domain); HD2 – druga domena helikalna zwana inaczej domeną superhelikalną (second helical domain); HDAC – deacetylazy histonów (histone deacetylases); Hsps – białka szoku cieplnego (heat shock proteins); IAPS – białka inhibitorowe apoptozy (inhibitors of apoptosis proteins); NBD – domena wiążącą nukleotydy typu α/b (nucleotide binding domain); NOD – domena wiązania nukleotydów i oligomeryzacji (nucleotide binding and oligomerization domain); Omi/HtrA2 – proteaza serynowa, antagonista IAPs, wiążąca inhibitory kaspaz (IAPs) (high temperature requirement A2 serine protease); RING – domena białek IAP o strukturze palca cynkowego, która ma aktywność ligazy E3 ubikwitynowej (really interesting new gene); Smac/DIABLO – mitochondrialny czynnik 2 aktywujący kaspazy, wiążący inhibitory kaspaz (IAPs) (second mitochondria-derived activator of caspases/direct IAP binding protein with low pI); TNF – czynnik martwicy nowotworów (tumor necrosis factor); WHD – domena skrzydlatej helisy (winged helix domain); WRD – domena powtórzeń WD40 (WD40 repeat domain); XIAP – inhibitor apoptozy sprzężony z chromosomem X (X-linked inhibitor of apoptosis protein).
Wstęp
Apoptoza jest ewolucyjnie zachowanym, genetycznie kontrolowanym procesem programowanej śmierci komórki, który stanowi istotny element prawidłowego rozwoju organizmu i homeostazy tkanki [4,17]. Apoptoza powoduje wiele zmian morfologicznych, takich jak zmniejszenie objętości komórki, obkurczanie cytoplazmy komórkowej, kondensację chromatyny, pączkowanie błony komórkowej, a w końcowym efekcie tworzenie ciałek apoptotycznych, które są szybko fagocytowane przez sąsiadujące komórki [32,35]. Rozregulowanie tego procesu może prowadzić do obniżenia odporności, rozwoju nowotworów, chorób autoimmunologicznych i neurodegeneracyjnych [4,21].
Celem apoptotycznego programu jest aktywacja kaspaz, swoistych proteaz, które powodują śmierć komórki poprzez hydrolizę różnych białek strukturalnych i funkcjonalnych [28,63]. Syntetyzowane są w komórce jako nieaktywne zymogeny (prokaspazy) i aktywowane przez autoproteolizę bądź też działanie innych kaspaz lub proteaz. Kaspazy inicjatorowe (kaspaza 2, 8, 9, 10), które są aktywowane jako pierwsze, zapoczątkowują działanie kaskady kaspaz wykonawczych (kaspaza 3, 6, 7) prowadzącej do śmierci komórki. Ich aktywacja wymaga jednak białkowego kompleksu, który zapewnia bliskie ułożenie zymogenów niezbędne do autokatalizy lub allosterycznie aktywuje zymogen bez przetwarzania go [7,49]. W komórkach ssaków do aktywacji kaspaz prowadzą dwa dobrze opisane szlaki: szlak związany z błoną komórkową (szlak zewnętrzny) i szlak zależny od mitochondriów (szlak wewnętrzny) [22,54,63].
Zewnętrzny szlak opiera się głównie na receptorach błonowych oraz ich ligandach. Są to przeważnie białka nadrodziny receptorów TNF (tumor necrosis factor) [5]. Ligand wiążąc się z receptorem odsłania domeny śmierci DD (death domain) od cytoplazmatycznej strony błony, co umożliwia przyłączenie cząsteczki adaptorowej FADD. Ta adaptorowa cząsteczka ma również domenę DED (death effector domain), poprzez którą łączy się z analogiczną domeną prokaspazy 8 lub 10, tworząc kompleks DISC, zwany inaczej degradosomem. W wyniku tego dochodzi do aktywacji przez autoproteolizę prokaspazy 8 lub 10, która następnie aktywuje prokaspazę 3 [7,18] (ryc. 1A).
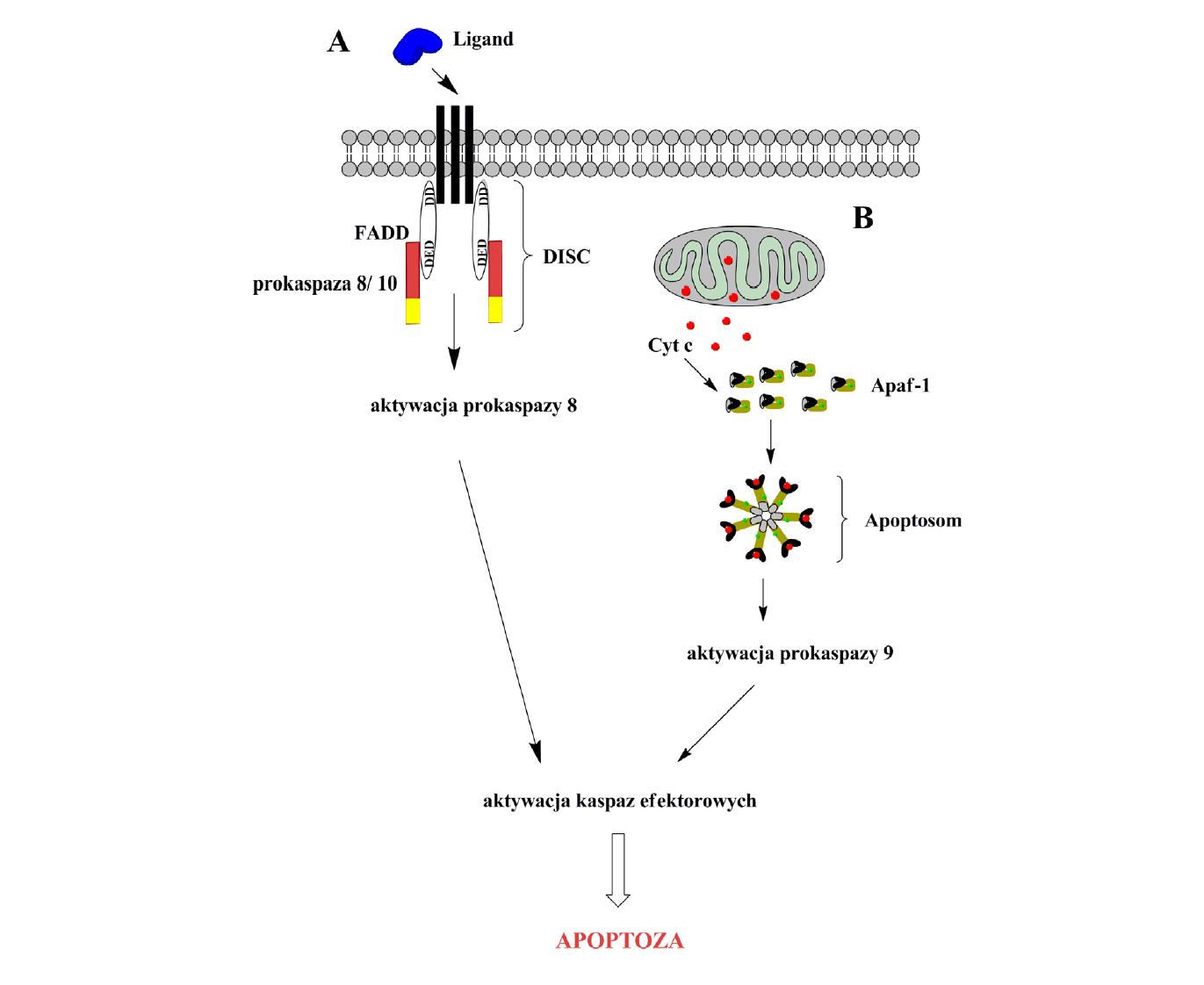
Ryc. 1. Schemat szlaków apoptozy; 1A – szlak zewnętrzny (receptorowy), 1B – szlak wewnętrzny (mitochondrialny)
Szlak mitochondrialny jest inicjowany na skutek wzrostu stężenia jonów wapnia w cytoplazmie, zaburzeń transportu elektrolitów, uszkodzeń DNA oraz wzrostu stężenia reaktywnych form tlenu, tlenku azotu lub toksyn [33]. W wyniku tych wewnątrzkomórkowych sygnałów następuje uwolnienie cytochromu c z przestrzeni międzybłonowej mitochondrium do cytoplazmy. Uwolniony cytochrom c łączy się z białkiem adaptorowym Apaf-1 i w obecności dATP/ATP tworzy cytosolowy kompleks białkowy zwany apoptosomem. Kompleks ten służy następnie jako platforma do aktywacji prokaspazy 9. Zaktywowana kaspaza 9 pozostaje związana z apoptosomem, rekrutując i aktywując kaspazy efektorowe 3 i/lub 7 [7,18,49] (ryc. 1 B).
Apoptosom
Odkrycie apoptosomu ma swoje korzenie w przełomowej pracy Wanga i wsp. [39]. Opracowali oni system aktywacji kaspaz oparty na ekstraktach cytosolowych pochodzących z komórek HeLa, który był stymulowany przez dATP. Stosując klasyczne metody oczyszczania białek oddzielili trzy czynniki aktywujące kaspazy (Apaf-1-3), które okazały się głównymi składowymi mitochondrialnego szlaku apoptozy. Apaf-1 był homologiem białka adaptorowego CED-4 nicienia Caenorhabditis elegans, Apaf-2 został zidentyfikowany jako cytochrom c [4], natomiast Apaf-3 jako kaspaza 9 [12,39]. Aktywacja prokaspazy 9 wymaga Apaf-1, który w obecności cytochromu c i dATP/ATP oligomeryzuje, tworząc platformę sygnalizacyjną w kształcie koła, zwaną apoptosomem [34].
Struktura Apaf-1
Apaf-1 jest 130 kDa białkiem występującym w cytosolu, pełniącym istotną rolę w zależnej od cytochromu c aktywacji kaspaz [11,14]. Apaf-1 składa się z trzech głównych domen: N-terminalnej domeny aktywacji i rekrutacji kaspaz (CARD), centralnej domeny wiązania nukleotydów i oligomeryzacji – NOD, określanej również jako NB-ARC oraz C-końcowej regulatorowej domeny 15 powtórzeń WD40 (WRD). Domena CARD jest odpowiedzialna za interakcję z domeną CARD prokaspazy 9 [7,37,49]. Funkcją domeny NOD, która zawiera motywy Walkera (zachowane motywy aminokwasów odpowiedzialne za wiązanie ATP/dATP i Mg2+), jest zależna od nukleotydów oligomeryzacja. Składa się z regionu ATP-azy, który umiejscawia Apaf-1 w rodzinie AAA+ ATP-az i jest zorganizowany w domenę wiążącą nukleotydy typu α/β (NBD) i domenę helikalną (HD1). Po tym regionie występuje domena skrzydlatej helisy (WHD) i druga domena helikalna (HD2) zwana inaczej domeną superhelikalną [24,40,49]. Region powtórzeń WD40 jest odpowiedzialny za wiązanie cytochromu c i musi negatywnie regulować Apaf- 1, gdyż jego usunięcie powoduje powstanie aktywnej postaci tego białka [2,6]. Zidentyfikowano cztery różne izoformy Apaf-1 w komórkach ssaków, znane jako Apaf-1 (Apaf-1S), Apaf-1XL/Apaf-1L-WD13, Apaf-1LN i Apaf-1LC/Apaf-1L [8,12], z których każda zawiera jedną lub dwie wstawki 11 aminokwasów na N-końcu i dodatkowe powtórzenie WD40 (43 aa) na C-końcu [14,22,40]. Dodatkowe powtórzenie WD- 40 ma zasadnicze znaczenie dla izoform, gdyż tylko te izoformy są w stanie wspierać aktywację kaspaz [8]. Wstawka 11-aminokwasowa między domenami CARD a NOD w niektórych izoformach prawdopodobnie nie bierze udziału w wiązaniu cytochromu c lub aktywacji kaspaz [12]. W przypadku braku sygnału apoptotycznego, Apaf-1 występuje jako nieaktywny monomer związany z dATP/ATP, w którym region domen CARD i NOD przyjmuje zwartą strukturę, a region powtórzeń WD40 dodatkowo go maskuje, co zapobiega oligomeryzacji i wiązaniu prokaspazy 9 [14,37,49] (ryc. 2A). Istnieją niejasności co do rodzaju związanego nukleotydu w nieaktywnym monomerze. Według badań przeprowadzonych przez Wanga i wsp. [34], monomeryczny Apaf-1 oczyszczony z komórek sf21 zawierał dATP. Wyniki innych badań zaprzeczają tym ustaleniom wykazując, że oczyszczony Apaf-1 z komórek różnych gatunków owadów i Escherichia coli był związany z ADP [5,48,80].

Ryc. 2. Mechanizm powstawania apoptosomu; A – zablokowana postać Apaf-1, B – półotwarta postać Apaf-1, C – struktura apoptosomu (opis w tekście)
Znaczenie nukleotydów i cytochromu c w powstawaniu apoptosomu
Apaf-1 występuje w nieaktywnej postaci w komórkach i jest aktywowany przez wiązanie cytochromu c i dATP/ATP. Cytochrom c uwolniony z mitochondrium wiąże się z domeną powtórzeń WD40 białka Apaf-1 odsłaniając zablokowane N-terminalne regiony i wyzwalając hydrolizę związanego z Apaf-1 dATP/ATP do dADP/ADP [18,37] (ryc. 2B). W przypadku gdy związanym nukleotydem jest ADP hydroliza nie jest konieczna do powstania apoptosomu [7,80]. Mimo że blokada zostaje zwolniona, badania z użyciem Apaf-1, w którym region WD40 został usunięty (ΔWD40) wykazały, że domeny CARD, ATP-azy (domena NBD, domena helikalna 1) i domena skrzydlatej helisy tworzą zwartą strukturę z dADP/ADP w jej centrum. W wyniku tego domena CARD jest niedostępna dla prokaspazy 9 i osłabiona jest zdolność Apaf-1 do tworzenia oligomerów [6,49]. Związany z dADP/ADP Apaf-1 pozostaje nieaktywny, dopiero wymiana dADP/ADP na cytosolowy dATP/ATP powoduje zmiany konformacyjne, dzięki którym możliwe jest powstanie apoptosomu [34,77]. Wymiana dADP/ADP na dATP/ATP stanowi mechanizm kontrolny, dADP/ADP blokuje Apaf-1, podczas gdy wiązanie dATP/ATP przełącza Apaf-1 do konformacji aktywnej. Apaf-1 wykorzystuje hydrolizę i wymianę dADP/ADP-dATP/ATP w celu przełączania między poszczególnymi konformacjami [6,49]. W przypadku braku dATP/ATP Apaf-1 nie będzie tworzył apoptosomu, zamiast tego może tworzyć nieaktywne agregaty [34].
Struktura apoptosomu
Liczne badania oparte na kriomikroskopii elektronowej zostały przeprowadzone w celu identyfikacji struktury apoptosomu. Prace Yuana i wsp. [80] dostarczają obrazu tej platformy w rozdzielczości 9,5 A. Wykazały one kompleks w kształcie koła składający się z siedmiu cząsteczek Apaf-1, w którym domeny CARD tworzą pierścień usytuowany powyżej centralnego rdzenia (ryc. 2C). Rdzeń ten składa się z dwóch pierścieni: pierścienia wewnętrznego zbudowanego z domen NBD, otoczonego pierścieniem zewnętrznym, który zawiera domenę helikalną 1 (HD1) i domenę skrzydlatej helisy (WHD). W centralnym rdzeniu domena WHD stanowi pomost między dwiema sąsiednimi parami domen NBD-HD1. Siedem ramion utworzonych przez domenę HD2 odchodzi od rdzenia. Na końcu każdego ramienia znajduje się obszar regulacyjny, czyli region 15 powtórzeń WD-40, który tworzy dwie domeny siedmiopłatowego i ośmiopłatowego beta-śmigła z cytochromem c umieszczonym pomiędzy nimi [1,48,77,80].
Aktywacja prokaspazy 9
Powstawanie apoptosomu pozwala na odsłonięcie domen CARD białek Apaf-1. Odsłonięte domeny CARD Apaf-1 są luźno związane z apoptosomem poprzez łącznik CARD-NBD i mogą oddziaływać z domenami CARD prokaspaz 9, tworząc strukturę podobną do dysku, znajdującą się powyżej centralnego rdzenia [48,77,80]. Mogłoby się wydawać, że do będącego heptametrem apoptosomu powinno się przyłączać siedem cząsteczek prokaspazy 9, jednak ostatnie wyniki badań sugerują, że Apaf-1 i prokaspaza 9 nie występują w stosunku 1:1 w apoptosomie [42,70].
Tak jak inne kaspazy, kaspaza 9 jest dimerem w aktywnej postaci, w odróżnieniu jednak od kaspaz efektorowych zymogen kaspazy 9 jest przede wszystkim monomerem w fizjologicznym stężeniu [7,18]. Kaspazy inicjatorowe są aktywowane na ogół przez autokatalityczne wewnątrzłańcuchowe cięcia, jednak okazało się, że nie są one niezbędne do aktywacji prokaspazy 9, prawdopodobnie ze względu na elastyczność wynikającą z niezwykle długiego łącznika między jej podjednostką dużą i małą [2,50,58]. Zaktywowana kaspaza 9 pozostaje połączona z apoptosomem i wykazuje katalityczną aktywność, która jest o trzy rzędy wielkości wyższa niż uwolnionej kaspazy 9, co może świadczyć o tym, że tak powstały kompleks działa jak holoenzym [7,51]. Aktywna kaspaza 9 może potem aktywować prokaspazę 3 i prokaspazę 7, co prowadzi do proteolizy białek i zmian morfologicznych w komórce, a ostatecznie do jej śmierci [54]. Poza tym badania wykazały, że apoptosom ma znaczenie w zwiększaniu powinowactwa kaspazy 9 do prokaspazy 3 [76]. Immunoprecypitacja katalitycznie aktywnych apoptosomów z komórek Jurkat wykazała, że oprócz prokaspazy 9 do apoptosomu może się przyłączać prokaspaza 3 i białko XIAP (X-linked inhibitor of apoptosis protein) [23]. Okazuje się, że wzajemne oddziaływanie XIAP z kaspazą 9 jest niezbędne do przyłączenia prokaspazy 3 do apoptosomu. Natomiast Smac lub Omi/HtrA2, które są uwalniane z mitochondrium wraz z cytochromem c, mogą wypierać XIAP z apoptosomu, a tym samym zwiększać aktywność apoptosomu [65].
Zaproponowano trzy modele aktywacji prokaspazy 9. Według modelu indukcji bliskością (induced proximity model) kaspazy inicjatorowe ulegają aktywacji, gdy znajdą się w bliskiej odległości między sobą, co zapewnia wiązanie do odpowiednich kompleksów białkowych [7,53]. Jest on podsumowaniem badań, w których zastosowano białko fuzyjne między kaspazami docelowymi i heterologiczną domeną dimeryzacji, taką jak tandem FKBPs. Wiązanie domeny z dimerycznym ligandem FK1012 przybliża kaspazy do siebie, czego wynikiem jest ich aktywacja [7]. Modyfikacją tego modelu jest model napędzanej bliskością dimeryzacji (proximity-driven dimerization model) [9]. Według tego modelu apoptosom rekrutuje monomery prokaspazy 9, umiejscawiając je blisko siebie. Takie rozmieszczenie zwiększa lokalne stężenie monomerów prokaspazy 9 w apoptosomie i sprzyja dimeryzacji, a tym samym ich aktywacji. Jak określono przez strukturę krystaliczną dimeru kaspazy 9, tylko jedna z dwóch podjednostek w dimerze jest aktywna katalitycznie [7,18,54]. Przypuszczalnie nieaktywne monomery prokaspazy 9 koncentrują się w centrum apoptosomu rekrutując dodatkowe monomery prokaspaz do tworzenia aktywnego holoenzymu [1,58]. Kolejny model to model indukcji zmian konformacyjnych (induced conformation model), apoptosom powodując konformacyjne zmiany w prokaspazie 9 i ułatwiając homooligomeryzację ma aktywną rolę w jej aktywacji. Jest to oparte na obserwacji, że skonstruowana dimeryczna kaspaza 9 ma inną aktywność w stosunku do kaspazy 9 zaktywowanej przez apoptosom [7,16]. Prokaspaza 9 może być aktywowana jako monomer z konformacją jej miejsca aktywnego stabilizowaną bezpośrednio przez apoptosom lub jako homodimer, zgodnie z modelem napędzanej bliskością dimeryzacji, z konformacją jej miejsca aktywnego stabilizowaną przez dimer i oddziaływanie dimeru z apoptosomem. Istnieje jeszcze możliwość, że prokaspaza 9 jest aktywowana jako homooligomer wyższego rzędu z konformacją jej miejsca aktywnego stabilizowaną zarówno przez oligomer jak i apoptosom [79]. Przedstawione modele nie muszą wykluczać się wzajemnie. W modelu indukcji zmian konformacyjnych dimeryzacja jest traktowana jako szczególny przypadek, a nie ogólny mechanizm aktywacji kaspaz inicjatorowych [59].
Apoptosom Caenorhabditis elegans
Po raz pierwszy apoptosom zidentyfikowano u nicienia Caenorhabditis elegans. Analiza genetyczna doprowadziła do identyfikacji czterech genów ced-3, ced-4, ced-9 i egl-1, które kontrolują apoptozę 131 komórek somatycznych podczas rozwoju C. elegans [8,28]. Gdy komórki są przeznaczone do śmierci CED-3, jedyna znana apoptotyczna kaspaza u nicienia występująca prawidłowo w postaci zymogenu, jest aktywowana przez białko adaptorowe CED-4. Jest ono strukturalnie podobne do ssaczego Apaf-1 [60], zawiera N-końcową domenę rekrutacji kaspaz (CARD) [27], która pozwala mu wiązać się z CARD CED-3 i region NB-ARC, który umożliwia zależną od nukleotydów oligomeryzację [85], nie ma natomiast powtórzeń WD-40 [2,54]. W przypadku braku bodźca apoptotycznego, oligomeryzacja CED-4 jest zahamowana przez oddziaływanie z antyapoptotycznym białkiem CED-9, które jest związane z błoną mitochondrialną [12,54]. Jedna cząsteczka CED-9 współdziała z asymetrycznym dimerem CED-4, ale tylko specyficznie rozpoznaje jedną z dwóch cząsteczek CED-4 poprzez rozległy obszar wzajemnego oddziaływania. Ta swoista interakcja zapobiega aktywacji CED-3 przez CED-4 [72,74]. CED-9 ma sekwencję homologiczną do antyapoptotycznych członków rodziny Bcl-2, jednak u kręgowców białka te nie regulują bezpośrednio powstawania apoptosomu tylko działają jako regulatory uwalniania cytochromu c z mitochondrium [7,54]. Zwiększenie transkrypcji białka EGL-1 w komórkach skazanych na śmierć i jego wiązanie z CED-9 powoduje zmiany konformacyjne CED-9, które prowadzą do jego dysocjacji z kompleksu (CED-4)2-CED-9. Uwolniony dimer CED-4 dalej oligomeryzuje do postaci tetrameru, co ułatwia autoaktywację CED-3 [12,72,73]. Uważa się, że oligomeryzacja CED-4 prowadzi do nagromadzenia cząsteczek CED-3 blisko siebie, co prowadzi do międzycząsteczkowych autokatalitycznych cięć i aktywacji sąsiednich cząsteczek CED-3 [12,75]. Apoptosom C. elegans zawiera osiem białek CED-4, zorganizowanych jako tetramer asymetrycznych dimerów CED-4. Tych osiem cząsteczek CED-4 tworzy strukturę w kształcie lejka. Oktameryczny apoptosom CED-4 wiąże tylko dwie cząsteczki CED- 3 [47,63].
Apoptosom Drosophila melanogaster
Oprócz nicienia C. elegans, apoptosom i jego komponenty zidentyfikowano u Drosophila melanogaster. Analizując genom znaleziono potencjalny homolog ssaczego Apaf-1, białko Dark/Hac-1/Dapaf-1 [54]. Tak jak Apaf- 1, ma ono N-końcową domenę CARD, region NB-ARC z jego charakterystyczną domeną AAA+ ATP-azy i C-końcową domenę powtórzeń WD40. Wprawdzie Dark D. melanogaster zawiera powtórzenia WD40 nie wymaga jednak cytochromu c i może być aktywowany przez nieznany niemitochondrialny ligand [2,78,79]. Podobnie jak CED-4, Dark pełni rolę cząsteczki adaptorowej promującej oligomeryzację i aktywację kaspazy inicjatorowej Dronc, która z kolei rozszczepia i aktywuje kaspazę efektorową DrICE [6]. Strukturę apoptosomu D. melanogaster poznano dzięki elektronowej kriomikroskopii przy rozdzielczości 18.8 A. Aktywny apoptosom jest oktamerem w kształcie koła, który powstaje w obecności dATP [59]. W zależności od stężenia Dark apoptosom D. melanogaster może występować jako pojedynczy pierścień lub podwójny z dwoma oktamerycznymi pierścieniami leżącymi jeden na drugim [79].
Regulacja aktywności apoptosomu
Komórka ma wiele mechanizmów zapewniających to, że aktywne apoptosomy powstają tylko w odpowiedzi na działanie czynników apoptotycznych. Wiele czynników jest zaangażowanych w tworzenie apoptosomu i jego regulację w fizjologicznych warunkach (ryc. 3). Fizjologiczne stężenia potasu [13], wapnia [5] i nukleotydów hamują powstawanie apoptosomu [16]. Potas powstrzymuje oligomeryzację białka Apaf-1, wapń wiąże się z Apaf-1 nie dopuszczając do wymiany nukleotydów, natomiast ATP wiąże się z resztami lizyny obecnymi na powierzchni cytochromu c, nie dopuszczając do jego wiązania z białkiem Apaf-1 [18,20,54]. Podczas apoptozy hamujące działanie jest zniesione, gdy stężenie cytochromu c w cytosolu zwiększa się, a stężenie tych czynników maleje. Mechanizmy te zapewniają, że apoptosom nie powstaje w przypadku wydzielania małych ilości cytochromu c w zdrowych komórkach [20,22,37].
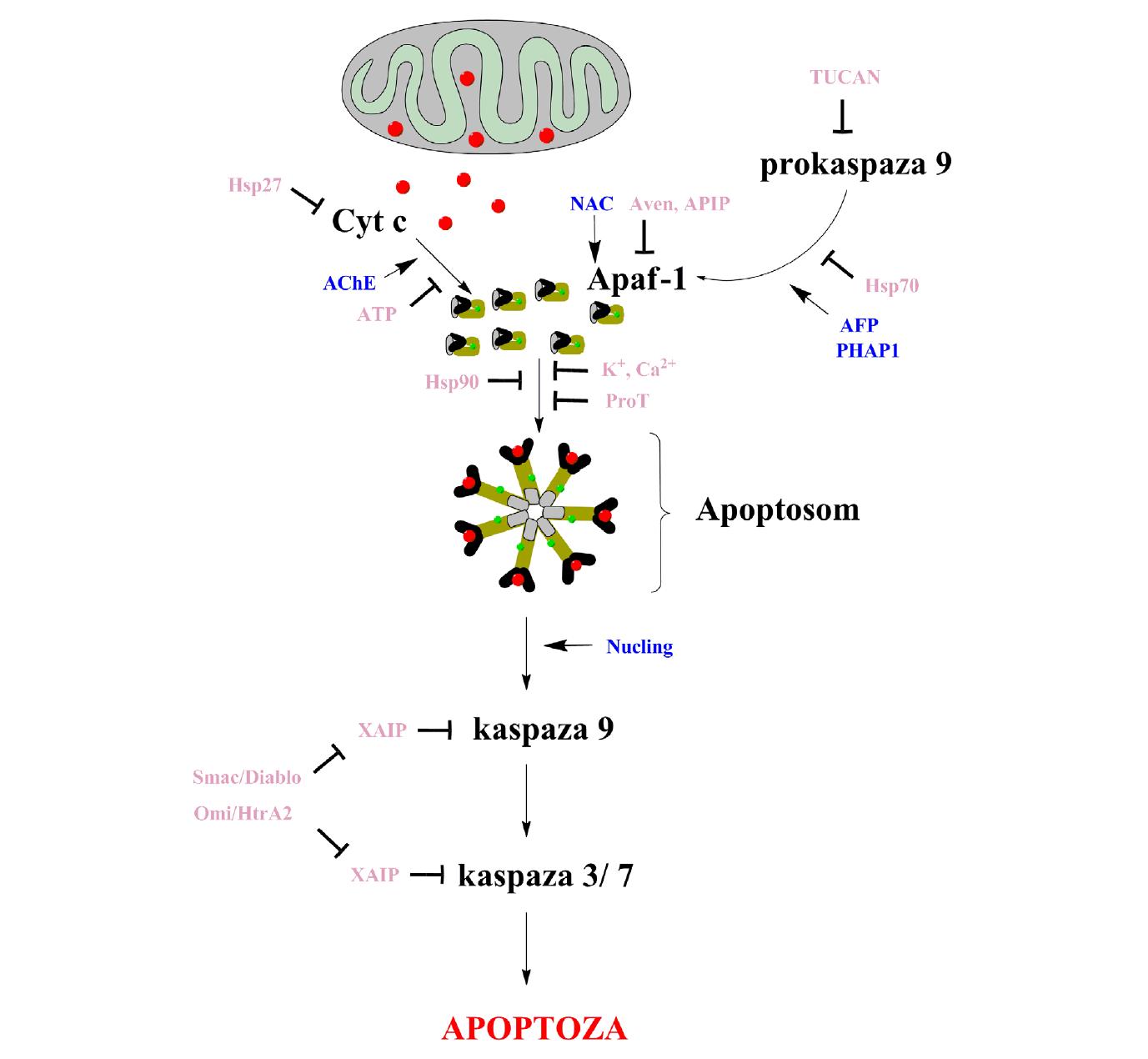
Ryc. 3. Regulacja aktywności apoptosomu (opis w tekście)
Istnieje wiele białek, które wpływają na aktywność apoptosomu poprzez bezpośrednią interakcję z jego składowymi. Białka szoku cieplnego (Hsps) są molekularnymi chaperonami, które są aktywowane w odpowiedzi na stres komórkowy, a ich rolą jest ochrona komórki [31]. Wykazano, że białka te zaangażowane są w hamowanie apoptozy, regulując powstawanie apoptosomu. Hsp70 wiąże się z domeną CARD białka Apaf-1, uniemożliwiając w ten sposób przyłączenie prokaspazy 9 [18,20,22]. Hsp90 oddziałuje z białkiem Apaf-1, hamując jego oligomeryzację, a przez to interakcję między Apaf-1 a prokaspazą 9. Natomiast Hsp27 hamuje apoptozę poprzez wiązanie cytosolowego cytochromu c, zapobiegając jego interakcji z białkiem Apaf-1 [20,22,54].
Białka IAPs (inhibitors of apoptosis proteins) zaangażowane są w regulację aktywności apoptosomu. Każde białko IAP zawiera od jednej do trzech kopii domen BIR (baculoviral IAP repeat), które pozwalają im wiązać się z kaspazami, a przez to hamować ich aktywność. Niektóre IAPs oprócz BIR mają także domenę RING (really interesting new gene). Najbardziej poznanym białkiem IAP jest białko XIAP (X-linked inhibitor of apoptosis protein) [17,18]. Białko XIAP wiąże się z natywnym apoptosomem, oddziałując z małą podjednostką p12 zaktywowanej kaspazy 9 poprzez domenę BIR3, a jednocześnie z kaspazą 3 i 7 poprzez domenę BIR2, która wymaga obecności łącznika ją poprzedzającego. Poprzez domenę RING, która ma aktywność ligazy E3 ubikwitynowej, XIAP prowadzi do ubikwitynacji, a następnie do degradacji kaspazy 3 i 9 [10,56]. W przypadku niewielkiego stresu komórkowego mała ilość Apaf-1 oligomeryzuje i rekrutuje prokaspazę 9, która przetwarzana jest do p35/p12, a jej aktywność hamowana jest przez XIAP. Tak więc, selektywność XIAP do przetworzonej postaci holoenzymu Apaf-1/Kaspaza 9 może stanowić mechanizm bezpiecznego wyłącznika jego przypadkowej aktywacji. Jednak wpływ bodźców chemicznych spowoduje powstanie tylu kompleksów apoptosomu, poprzez które dojdzie do aktywacji wystarczającej ilości prokaspazy 3, zdolnej do przezwyciężenia hamującego efektu XIAP, ostatecznie prowadząc do śmierci komórki [11].
Mitochondrialne białka Smac/Diablo oraz Omi/HtrA2, które są wydzielane z mitochondriów wraz z cytochromem c, znoszą hamujący wpływ białka XIAP na aktywność kaspaz, wiążąc się z nim i odrywając go od apoptosomu [19,66]. Białko Smac zawiera motyw AVPI na N-końcu, który wiąże się do domeny BIR3 białka XIAP, wypierając motyw ATPF kaspazy 9 [69]. Smac jednocześnie wiąże się ciasno do domeny BIR2 białka XIAP, stabilizując wiązanie motywu AVPI do domeny BIR3 poprzez motyw AVPI. Wiązanie Smac/Diablo, Omi/HtrA2 i prawdopodobnie innych, jak dotąd niezidentyfikowanych białek może przeciwdziałać wiązaniu XIAP do kaspazy 9 i w ten sposób regulować aktywność apoptosomu [11].
Zarówno białko Aven jak i APIP wiążą się z białkiem Apaf-1, zapobiegając jego interakcji z prokaspazą 9 [54]. Podobnie TUCAN (tumor-up-regulated CARD-containing antagonist of caspase-9) i jego izoforma TUCAN-54 hamują tę interakcję, wiążąc prokaspazę 9 poprzez ich domenę CARD [46,71]. Oprócz tych inhibitorów aktywności apoptosomu zidentyfikowano białko Nucling, które zwiększa pośredniczoną przez apoptosom aktywację prokaspazy 9 poprzez oddziaływanie z kompleksami Apaf-1/prokaspaza 9, ich stabilizację i przemieszczenie tak powstałych kompleksów do jądra [58]. Białko NAC może oddziaływać z białkiem Apaf-1 poprzez domenę CARD i promować aktywację kaspaz połączonych z apoptosomem [54].
Odkryto wiele czynników, które regulują aktywność apoptosomu bez bezpośredniego fizycznego kontaktu lub modyfikacji jego składowych. PHAP1 (putative HLA-DR-associated protein-1) i protymozynę-alfa (ProT) określa się jako „regulatory apoptosomu”. PHAP1 ułatwia przyłączanie prokaspazy 9 do apoptosomu po oligomeryzacji Apaf-1 [20,37,54]. Ostatnie badania wykazały także jego rolę w zwiększaniu aktywności katalitycznej kaspazy 3. Dodatkowo zaobserwowano zwiększoną ekspresję białka PHAP1 w komórkach raka piersi, przez co są one wrażliwe na indukowaną przez cytochrom c aktywację kaspaz. Mikroiniekcje cytochromu c powodują śmierć komórek raka piersi in vitro na skutek zwiększonego wiązania prokaspazy 9 do białka Apaf-1 i następującej aktywacji kaspaz efektorowych [18,20,55]. Natomiast protymozyna-α (ProT) blokuje stymulującą aktywność PHAP1, hamuje powstawanie apoptosomu i następującą aktywację prokaspazy 9, a wyciszenie ekspresji ProT uwrażliwia komórki HeLa na apoptozę indukowaną przez promieniowanie UV i staurosporynę [37,54]. Acetylocholinoesteraza (AChE) promuje interakcję między cytochromem c a białkiem Apaf-1 [45], natomiast α-fetoproteina (AFP) pobudza aktywację kaspaz poprzez ich rekrutację do apoptosomu [57]. Poza tym donory tlenku azotu mogą wpływać na aktywność apoptosomu hamując oligomeryzację białka Apaf-1 i jego oddziaływanie z prokaspazą 9 [81].
Apoptosom jako cel terapii
Zmiany w regulacji procesu apoptozy mogą się przyczynić do patogenezy nowotworów i odporności komórek nowotworowych na chemioterapię. Przeprowadzono wiele prób zmierzających do korekty upośledzonego lub nadmiernie zachodzącego procesu apoptozy w zwierzęcych i komórkowych modelach chorób, wykorzystując rekombinowane biomolekuły, strategię antysensu, terapię genową lub związki organiczne. Były one głównie wycelowane w receptory śmierci, białka rodziny Bcl-2, kaspazy i inhibitory apoptozy [18,37]. Natomiast stosunkowo niewiele uwagi poświęcono aktywacji apoptosomu [20,54].
Niedawne badania wykazały, że zaburzenia aktywacji apoptosomu mogą mieć istotne znaczenie w różnych rodzajach nowotworów i ich reakcjach na leki cytotoksyczne. Po raz pierwszy modulację funkcji apoptosomu w przypadku nowotworów zaobserwowano w przerzutującym czerniaku złośliwym, który jest odporny na wszelkie leki przeciwnowotworowe [11,60]. Modulacja ta polegała na utracie ekspresji Apaf-1 przez mechanizm, który prawdopodobnie związany jest z transkrypcyjnym wyciszaniem poprzez metylację, ponieważ w liniach komórkowych czerniaka 5-aza-2′-deoksycytydyna (5aza2dC), inhibitor metylacji, przywraca ekspresję Apaf-1. Co ciekawe, guzy czerniaka niewykazujące ekspresji Apaf-1 są odporne na czynniki cytotoksyczne, ale stają się wrażliwe, gdy ekspresja Apaf-1 jest przywrócona przez 5aza2dC [60]. Białko Apaf-1 jest często wyciszone lub inaktywowane w ludzkich nowotworach, w tym glejakach, liniach komórek białaczkowych, i nowotworach jajnika [20,22]. Proces apoptozy w liniach komórek raka szyjki macicy i liniach ludzkich komórek białaczkowych jest upośledzony na skutek niedoboru Apaf-1, a jego wprowadzenie do tych komórek prowadzi do przywrócenia tego procesu [26,30]. W liniach komórkowych raka jajnika, osłabiona aktywność Apaf-1, a co za tym idzie apoptosomu jest związana z odpornością na chemioterapię [36,68]. Dodatkowo utrata ekspresji Apaf-1 jest związana z progresją guza i niekorzystnym rokowaniem w raku jelita grubego, a wyciszenie Apaf-1 w przewlekłej białaczce limfatycznej B-komórkowej (B-CLL) koreluje ze złym rokowaniem w przypadku towarzyszących mutacji p53 [61,82]. Te badania wskazują, że apoptosom może odgrywać główną rolę w cytotoksycznym działaniu leków chemioterapeutycznych a zaburzenia jego funkcji poprzez zmiany w ekspresji lub funkcji Apaf-1 mogą być przyczyną odporności nowotworów na leki [11]. Dlatego też trwają badania nad stworzeniem lub znalezieniem związków, które będą indukować lub uniemożliwiać tworzenie aktywnego kompleksu apoptosomu, co będzie miało duże znaczenie w przypadku leczenia chorych na nowotwory lub choroby wynikające z nadmiernie zachodzącego procesu apoptozy [18].
Zastosowanie związków, które zwiększają ekspresję składowych apoptosomu lub wywołują uwolnienie cytochromu c z mitochondriów może się stać jednym ze sposobów skierowanego na apoptosom leczenia chorych z nowotworami. Na przykład LAQ824, inhibitor rodziny HDAC (deacetylazy histonów deacetylują nukleosomalne histony zmniejszając ekspresję genów), wywołuje śmierć komórek poprzez indukcję ekspresji białka Apaf-1 i prokaspazy 9 [67]. Triacsin c, inhibitor syntazy acylo-CoA, selektywnie wywołuje wydzielanie cytochromu c i pośredniczoną przez apoptosom apoptozę w komórkach [43]. Poza tym próbuje się także opracować małe cząsteczki będące mimetykami cytochromu c, które spowodują zmiany konformacyjne w Apaf-1, niezbędne do powstania apoptosomu i aktywacji prokaspazy 9 [29]. U podstaw tej strategii leży odkrycie, że komórki raka piersi są niezwykle wrażliwe na indukowaną przez cytochrom c apoptozę w porównaniu do prawidłowych komórek nabłonka gruczołu mlekowego, co związane jest z nadekspresją apoptosomalnego regulatora PHAP1. Wprowadzając cytochrom c lub czynnik naśladujący jego działanie można selektywnie zabić komórki raka piersi [55]. α-(trichlorometylo)-4-pyridynoetanol (PETCM) zwiększa in vitro indukowaną przez cytochrom c aktywację prokaspazy 3, stymulując tworzenie apoptosomu przez hamowanie protymozyny (ProT), onkoproteiny, która działa jako negatywny regulator tworzenia apoptosomu [27]. Związki indolu i karbaminiany również indukują aktywację kaspaz, działając jako bezpośrednie aktywatory powstawania apoptosomu, a jeden z nich (tzw. związek 2) działa cytotoksycznie na nowotworowe linie komórkowe, wykazując słabe działanie w stosunku do prawidłowych linii komórkowych. Aktywność PETCM i związku 2 wymaga obecności cytochromu c [37,44].
Aromatyczne pochodne mocznika są skutecznymi inhibitorami aktywacji kaspaz w lizatach komórkowych. Prawdopodobnie ich działanie polega na hamowaniu powstawania apoptosomu [36]. Tauryna (kwas 2-aminoetanosulfonowy) zapobiega indukowanej przez niedokrwienie apoptozie w kardiomiocytach, hamując tworzenia apoptosomu prawdopodobnie przez zmniejszenie ekspresji prokaspazy 9 [62]. Oligomery N-alkiloglicyny (peptoidy), a zwłaszcza peptoid 1a wiążą się odwracalnie do Apaf-1, uniemożliwiając przyłączenie prokaspazy 9, a tym samym aktywację kaskady apoptozy [41].
Podsumowanie
Apoptosom jest centralnym elementem mitochondrialnego szlaku apoptozy. Powstaje wyłącznie w odpowiedzi na działanie czynników apoptotycznych, a jego rola polega na aktywacji inicjatorowej prokaspazy 9, co doprowadza do uruchomienia kaskady kaspaz wykonawczych i ostatecznie śmierci komórki. Do powstania apoptosomu potrzebne jest białko Apaf-1, cytochrom c i dATP/ATP, chociaż nie jest wykluczone, że inne cząsteczki mogą regulować aktywność apoptosomu in vivo. Informacje przedstawione w pracy dowodzą, że zaburzona aktywność apoptosomu jest przyczyną rozwoju niektórych nowotworów i chemiooporności. Stwarza to możliwość zastosowania regulatorów powstawania apoptosomu jako terapeutyków. Istnienie związków, które indukują lub uniemożliwiają tworzenie aktywnego kompleksu apoptosomu sugeruje, że już niedługo apoptosom może się stać realnym celem terapeutycznym, nie tylko w przypadku leczenia chorych na nowotwory, ale również na choroby neurodegeneracyjne.
PIŚMIENNICTWO
[1] Acehan D., Jiang X., Morgan D.G., Heuser J.E., Wang X., Akey C.W.: Three-dimensional structure of the apoptosome: implications for assembly, procaspase-9 binding, and activation. Mol. Cell., 2002; 9: 423-432
[PubMed] [Full Text HTML] [Full Text PDF]
[2] Adams J.M., Cory S.: Apoptosomes: engines for caspase activation. Curr. Opin. Cell Biol., 2002; 14: 715-720
[PubMed]
[3] Adrain C., Brumatti G., Martin S.J.: Apoptosomes: protease activation platforms to die from. Trends Biochem. Sci., 2006; 31: 243-247
[PubMed]
[4] Ameisen J.C.: On the origin, evolution, and nature of programmed cell death: a timeline of four billion years. Cell Death Differ., 2002; 9: 367-393
[PubMed]
[5] Bao Q., Lu W., Rabinowitz J.D., Shi Y.: Calcium blocks formation of apoptosome by preventing nucleotide exchange in Apaf-1. Mol. Cell, 2007; 25: 181-192
[PubMed] [Full Text HTML] [Full Text PDF]
[6] Bao Q., Riedl S.J., Shi Y.: Structure of Apaf-1 in the auto-inhibited form: a critical role for ADP. Cell Cycle 2005; 4: 1001-1003
[PubMed] [Full Text PDF]
[7] Bao Q., Shi Y.: Apoptosome: a platform for the activation of initiator caspases. Cell Death Differ., 2007; 14: 56-65
[PubMed] [Full Text HTML] [Full Text PDF]
[8] Benedict M.A., Hu Y., Inohara N., Núnez G.: Expression and functional analysis of Apaf-1 isoforms. Extra Wd-40 repeat is required for cytochrome c binding and regulated activation of procaspase-9. J. Biol. Chem., 2000; 275: 8461-8468
[PubMed] [Full Text HTML] [Full Text PDF]
[9] Boatright K.M., Salvesen G.S.: Mechanisms of caspase activation. Curr. Opin. Cell Biol., 2003; 15: 725-731
[PubMed]
[10] Bratton S.B., Lewis J., Butterworth M., Duckett C., Cohen G.M.: XIAP inhibition of caspase-3 preserves its association with the Apaf-1 apoptosome and prevents CD95-and Bax-induced apoptosis. Cell Death Differ., 2002; 9: 881-892
[PubMed] [Full Text HTML] [Full Text PDF]
[11] Cain K.: Chemical-induced apoptosis: formation of the Apaf-1 apoptosome. Drug Metab. Rev., 2003; 35: 337-363
[PubMed]
[12] Cain K., Bratton S.B., Cohen G.M.: The Apaf-1 apoptosome: a large caspase-activating complex. Biochimie, 2002; 84: 203-214
[PubMed]
[13] Cain K., Langlais C., Sun X.M., Brown D.G., Cohen G.M.: Physiological concentrations of K+ inhibit cytochrome c – dependent formation of the apoptosome. J. Biol. Chem., 2001; 276: 41985-41990
[PubMed] [Full Text HTML] [Full Text PDF]
[14] Campioni M., Santini D., Tonini G., Murace R., Dragonetti E., Spugnini E.P., Baldi A.: Role of Apaf-1, a key regulator of apoptosis, in melanoma progression and chemoresistance. Exp. Dermatol., 2005; 14: 811-818
[PubMed]
[15] Chandra D., Bratton S.B., Person M.D., Tian Y., Martin A.G., Ayres M., Fearnhead H.O., Gandhi V., Tang D.G.: Intracellular nucleotides act as critical prosurvival factors by binding to cytochrome c and inhibiting apoptosome. Cell, 2006; 125: 1333-1346
[PubMed] [Full Text HTML] [Full Text PDF]
[16] Chao Y., Shiozaki E.N., Srinivasula S.M., Rigotti D.J., Fairman R., Shi Y.: Engineering a dimeric caspase-9: a reevaluation of the induced proximity model for caspase activation. PLoS Biol., 2005; 3: e183
[PubMed] [Full Text HTML] [Full Text PDF]
[17] Czerski L., Nunez G.: Apoptosome formation and caspase activation: is it different in the heart? J. Mol. Cell Cardiol., 2004; 37: 643-652
[PubMed]
[18] D’Amelio M., Tino E., Cecconi F.: The apoptosome: emerging insights and new potential targets for drug design. Pharm. Res., 2008; 25: 740-751
[PubMed] [Full Text HTML] [Full Text PDF]
[19] Du C., Fang M., Li Y., Li L., Wang X.: Smac, a mitochondrial protein that promotes cytochrome c-dependent caspase activation by eliminating IAP inhibition. Cell, 2000; 102: 33-42
[PubMed] [Full Text HTML] [Full Text PDF]
[20] Fadeel B., Ottosson A., Pervaiz S.: Big wheel keeps on turning: apoptosome regulation and its role in chemoresistance. Cell Death Differ., 2008; 15: 443-452
[PubMed] [Full Text HTML] [Full Text PDF]
[21] Grzybowska-Izydorczyk O., Smolewski P.: Rola białek z rodziny inhibitora apoptozy (IAP) w chorobach rozrostowych układu krwiotwórczego. Postępy Hig. Med. Dośw., 2008; 62: 55-63
[PubMed] [Full Text HTML] [Full Text PDF]
[22] Hajra K.M., Liu J.R.: Apoptosome dysfunction in human cancer. Apoptosis, 2004; 9: 691-704
[PubMed]
[23] Hill M.M., Adrain C., Duriez P.J., Creagh E.M., Martin S.J.: Analysis of the composition, assembly kinetics and activity of native Apaf-1 apoptosomes. EMBO J., 2004; 23: 2134-2145
[PubMed] [Full Text HTML] [Full Text PDF]
[24] Hill M.M., Adrain C., Martin S.J.: Portrait of a killer: the mitochondrial apoptosome emerges from the shadows. Mol. Interv., 2003; 3: 19-26
[PubMed]
[25] Horvitz H.R.: Worms, life, and death (Nobel Lecture). Chembiochem, 2003; 4: 697-711
[PubMed]
[26] Jia L., Srinivasula S.M., Liu F.T., Newland A.C., Fernandes-Alnemri T., Alnemri E.S., Kelsey S.M.: Apaf-1 protein deficiency confers resistance to cytochrome c-dependent apoptosis in human leukemic cells. Blood, 2001; 98: 414-421
[PubMed] [Full Text HTML] [Full Text PDF]
[27] Jiang X., Kim H.E., Shu H., Zhao Y., Zhang H., Kofron J., Donnelly.J, Burns D., Ng S.C., Rosenberg S., Wang X.: Distinctive roles of PHAP proteins and prothymosin-alpha in a death regulatory pathway. Science, 2003; 299: 223-226
[PubMed]
[28] Jiang X., Wang X.: Cytochrome C-mediated apoptosis. Annu. Rev. Biochem., 2004; 73: 87-106
[PubMed]
[29] Johnson C.E., Huang Y.Y., Parrish A.B., Smith M.I., Vaughn A.E., Zhang Q., Wright K.M., Van Dyke T., Wechsler-Reya R.J., Kornbluth S., Deshmukh M.: Differential Apaf-1 levels allow cytochrome c to induce apoptosis in brain tumors but not in normal neural tissues. Proc. Natl. Acad. Sci. USA, 2007; 104: 20820-20825
[PubMed] [Full Text HTML] [Full Text PDF]
[30] Kamarajan P., Sun N.K., Sun C.L., Chao C.C.: Apaf-1 overexpression partially overcomes apoptotic resistance in a cisplatin-selected HeLa cell line. FEBS Lett., 2001; 505: 206-212
[PubMed]
[31] Kaźmierczuk A., Kiliańska Z.M.: Plejotropowa aktywność białek szoku cieplnego. Postępy Hig. Med. Dośw., 2009; 63: 502-521
[PubMed] [Full Text HTML] [Full Text PDF]
[32] Kaźmierczuk A., Kiliańska Z.M.: Rola białek szoku cieplnego w apoptozie komórek. Postępy Hig. Med. Dośw., 2010; 64: 273-283
[PubMed] [Full Text HTML] [Full Text PDF]
[33] Kiliańska Z.M., Miśkiewicz A.: Kaspazy kręgowców; ich rola w przebiegu apoptozy. Postępy Biol. Kom., 2003; 30: 129-152
[Abstract]
[34] Kim H.E., Du F., Fang M., Wang X.: Formation of apoptosome is initiated by cytochrome c-induced dATP hydrolysis and subsequent nucleotide exchange on Apaf-1. Proc. Natl. Acad. Sci. USA, 2005; 102: 17545-17550
[PubMed] [Full Text HTML] [Full Text PDF]
[35] Łabędzka K., Grzanka A., Izdebska M.: Mitochondrium a śmierć komórki. Postępy Hig. Med. Dośw., 2006; 60: 439-446
[PubMed] [Full Text HTML] [Full Text PDF]
[36] Lademann U., Cain K., Gyrd-Hansen M., Brown D., Peters D., Jäättelä M.: Diarlyurea compounds inhibit caspase activation by preventing the formation of ~700 kDa apoptosome complex. Mol. Cell Biol., 2003; 23: 7829-7837
[PubMed] [Full Text HTML] [Full Text PDF]
[37] Ledgerwood E.C., Morison I.M.: Targeting the apoptosome for cancer therapy. Clin. Cancer Res., 2009; 15: 420-424
[PubMed] [Full Text HTML] [Full Text PDF]
[38] Liu J.R., Opipari A.W., Tan L., Jiang Y., Zhang Y., Tang H., Nunez G.: Dysfunctional apoptosome activation in ovarian cancer: Implications for chemoresistance. Cancer Res., 2002; 62: 924-931
[PubMed] [Full Text HTML] [Full Text PDF]
[39] Liu X., Kim C.N., Yang J., Jemmerson R., Wang X.: Induction of apoptotic program in cell-free extracts: Requirement for dATP and cytochrome c. Cell, 1996; 86: 147-157
[PubMed] [Full Text HTML] [Full Text PDF]
[40] Mace P.D., Riedl S.J.: Molecular cell death platforms and assemblies. Curr. Opin. Cell Biol. 2010; 22: 828-836
[PubMed] [Full Text HTML] [Full Text PDF]
[41] Malet G., Martin A. G., Orzaez M., Vicent M. J., Masip I., Sanclimens G., Ferrer-Montiel A., Mingarro I., Messeguer A., Fearnhead H. O., Perez-Paya E.: Small molecule inhibitors of Apaf-1-related caspase-3/-9 activation that control mitochondrial-dependent apoptosis. Cell Death Differ., 2006; 13: 1523-1532
[PubMed] [Full Text HTML] [Full Text PDF]
[42] Malladi S., Challa-Malladi M., Fearnhead H.O., Bratton S.B.: The Apaf-1·procaspase-9 apoptosome complex functions as a proteolytic-based molecular timer. EMBO J., 2009; 28: 1916-1925
[PubMed] [Full Text HTML] [Full Text PDF]
[43] Mashima T., Oh-hara T., Sato S., Mochizuki M., Sugimoto Y., Yamazaki K., Hamada J., Tada M., Moriuchi T., Ishikawa Y., Kato Y., Tomoda H., Yamori T., Tsuruo T.: p53-defective tumors with a functional apoptosome-mediated pathway: a new therapeutic target. J. Natl. Cancer Inst., 2005; 97: 765-777
[PubMed] [Full Text HTML] [Full Text PDF]
[44] Nguyen J.T., Wells J.A.: Direct activation of the apoptosis machinery as a mechanism to target cancer cells. Proc. Natl. Acad. Sci. USA, 2002; 100: 7533-7538
[PubMed] [Full Text HTML] [Full Text PDF]
[45] Park S.E., Kim N.D., Yoo Y.H.: Acetylcholinesterase plays a pivotal role in apoptosome formation. Cancer Res., 2004; 64: 2652-2655
[PubMed] [Full Text HTML] [Full Text PDF]
[46] Pathan N., Marusawa H., Krajewska M., Matsuzawa S., Kim H., Okada K., Torii S., Kitada S., Krajewski S., Welsh K., Pio F., Godzik A., Reed J. C.: TUCAN, an antiapoptotic caspase-associated recruitment domain family protein overexpressed in cancer. J. Biol. Chem., 2001; 276: 32220-32229
[PubMed] [Full Text HTML] [Full Text PDF]
[47] Qi S., Pang Y., Hu Q., Liu Q., Li H., Zhou Y., He T., Liang Q., Liu Y., Yuan X., Luo G., Li H., Wang J., Yan N., Shi Y.: Crystal structure of the Caenorhabditis elegans apoptosome reveals an octameric assembly of CED-4. Cell, 2010; 141: 446-457
[PubMed] [Full Text HTML] [Full Text PDF]
[48] Riedl S.J., Li W., Chao Y., Schwarzenbacher R., Shi Y.: Structure of the apoptotic protease activating factor 1 bound to ADP. Nature, 2005; 434: 926-933
[PubMed]
[49] Riedl S.J., Salvesen G.S.: The apoptosome: signalling platform of cell death. Nat. Rev. Mol. Cell Biol., 2007; 8: 405-413
[PubMed]
[50] Riedl S.J., Shi Y.: Molecular mechanisms of caspase regulation during apoptosis. Nat. Rev. Mol. Cell Biol., 2004; 5: 897-907
[PubMed]
[51] Rodriguez J., Lazebnik Y.: Caspase-9 and Apaf-1 form an active holoenzyme. Genes Dev., 1999; 13: 3179-3184
[PubMed] [Full Text HTML] [Full Text PDF]
[52] Sakai T., Liu L., Teng X., Mukai-Sakai R., Shimada H., Kaji R., Mitani T., Matsumoto M., Toida K., Ishimura K., Shishido Y., Mak T.W., Fukui K.: Nucling recruits Apaf-1/pro-caspase-9 complex for the induction of stress-induced apoptosis. J. Biol. Chem., 2004; 279: 41131-41140
[PubMed] [Full Text HTML] [Full Text PDF]
[53] Salvesen G.S., Dixit V.M.: Caspase activation: the induced-proximity model. Proc. Natl. Acad. Sci. USA, 1999; 96: 10964-10967
[PubMed] [Full Text HTML] [Full Text PDF]
[54] Schafer Z.T., Kornbluth S.: The apoptosome: physiological, developmental, and pathological modes of regulation. Dev. Cell, 2006; 10: 549-561
[PubMed] [Full Text HTML] [Full Text PDF]
[55] Schafer Z.T., Parrish A.B., Wright K.M., Margolis S.S., Marks J.R., Deshmukh M., Kornbluth S.: Enhanced sensitivity to cytochrome c-induced apoptosis mediated by PHAPI in breast cancer cells. Cancer Res., 2006; 66: 2210-2218
[PubMed] [Full Text HTML] [Full Text PDF]
[56] Schimmer A.D., Dalili S., Batey R.A., Riedl S.J.: Targeting XIAP for the treatment of malignancy. Cell Death Differ., 2006; 13: 179-188
[PubMed] [Full Text HTML] [Full Text PDF]
[57] Semenkova L., Dudich E., Dudich I., Tokhtamisheva N., Tatulov E., Okruzhnov Y., Garcia-Foncillas J., Palop-Cubillo J.A., Korpela T.: Alpha-fetoprotein positively regulates cytochrome c-mediated caspase activation and apoptosome complex formation. Eur. J. Biochem., 2003; 270: 4388-4399
[PubMed] [Full Text HTML] [Full Text PDF]
[58] Shi Y.: Caspase activation: revisiting the induced proximity model. Cell, 2004; 117: 855-858
[PubMed] [Full Text HTML] [Full Text PDF]
[59] Shi Y.: Mechanical aspects of apoptosome assembly. Curr. Opin. Cell Biol. 2006; 18: 677-684
[PubMed]
[60] Soengas, M.S., Capodieci P., Polsky D., Mora J., Esteller M., Opitz-Araya X., McCombie R., Herman J.G., Gerald W.L., Lazebnik Y.A., Cordón-Cardó C., Lowe S.W.: Inactivation of the apoptosis effector Apaf-1 in malignant melanoma. Nature, 2001; 409: 207-211
[PubMed]
[61] Sturm I., Bosanquet A.G., Radetzki S., Hummel M., Dorken B., Daniel P.T.: Silencing of APAF-1 in B-CLL results in poor prognosis in the case of concomitant p53 mutation. Int. J. Cancer, 2006; 118: 2329-2336
[PubMed]
[62] Takatani T., Takahashi K., Uozumi Y., Shikata E., Yamamoto Y., Ito T., Matsuda T., Schaffer S. W., Fujio Y., Azuma J.: Taurine inhibits apoptosis by preventing formation of the Apaf1/caspase-9 apoptosome. Am. J. Physiol. Cell Physiol., 2004; 287: 949-953
[PubMed] [Full Text HTML] [Full Text PDF]
[63] Taylor R.C., Cullen S.P., Martin S.J.: Apoptosis: controlled demolition at the cellular level. Nat. Rev. Mol. Cell Biol., 2008; 9: 231-241
[PubMed]
[64] Teng X., Hardwick J.M.: The apoptosome at high resolution. Cell, 2010; 141: 402-404
[PubMed] [Full Text HTML] [Full Text PDF]
[65] Twiddy D., Brown D.G., Adrain C., Jukes R., Martin S.J., Cohen G.M., MacFarlane M., Cain K.: Pro-apoptotic proteins released from the mitochondria regulate the protein composition and caspase-processing activity of the native Apaf-1/caspase-9 apoptosome complex. J. Biol. Chem., 2004; 279: 19665-19682
[PubMed] [Full Text HTML] [Full Text PDF]
[66] Verhagen A.M., Ekert P.G., Pakusch M., Silke J., Connolly L.M., Reid G.E.: Identification of DIABLO, a mammalian protein that promotes apoptosis by binding to and antagonizing IAP proteins. Cell, 2000; 102: 43-53
[PubMed] [Full Text PDF]
[67] Wang S., Yan-Neale Y., Cai R., Alimov I., Cohen D.: Activation of mitochondrial pathway is crucial for tumor selective induction of apoptosis by LAQ824. Cell Cycle, 2006; 5: 1662-1668
[PubMed] [Full Text PDF]
[68] Wolf B.B., Schuler M., Li W., Eggers-Sedlet B., Lee W., Tailor P., Fitzgerald P., Mills G.B., Green D.R.: Defective cytochrome c-dependent caspase activation in ovarian cancer cell lines due to diminished or absent apoptotic protease activating factor-1 activity. J. Biol. Chem., 2001; 276: 34244-34251
[PubMed] [Full Text HTML] [Full Text PDF]
[69] Wu G., Chai J., Suber T.L., Wu J.W., Du C., Wang X., Shi Y.: Structural basis of IAP recognition by Smac/DIABLO. Nature, 2000; 408: 1008-1012
[PubMed]
[70] Würstle M.L., Laussmann M.A., Rehm M.: The central role of initiator caspase-9 in apoptosis signal transduction and the regulation of its activation and activity on the apoptosome. Exp. Cell Res., 2012; 318: 1213-1220
[PubMed]
[71] Yamamoto M., Torigoe T., Kamiguchi Y., Nakanishi K., Nabeta C., Asanuma H., Tsuruma T., Sato T., Hata F., Ohmura T., Yamaguchi K., Kurotaki T., Hirata K., Sato N.: A novel isoform of TUCAN is overexpressed in human cancer tissues and suppresses both caspase-8 and caspase-9 mediated apoptosis. Cancer Res., 2005; 65: 8706-8714
[PubMed] [Full Text HTML] [Full Text PDF]
[72] Yan N., Chai J., Lee E.S., Gu L., Liu Q., He J., Wu J.W., Kokel D., Li H., Hao Q., Xue D., Shi Y.: Structure of the CED-4-CED-9 complex provides insights into programmed cell death in Caenorhabditis elegans. Nature, 2005; 437: 831-837
[PubMed]
[73] Yan N., Gu L., Kokel D., Chai J., Li W., Han A., Chen L., Xue D., Shi Y.: Structural, biochemical, and functional analyses of CED-9 recognition by the proapoptotic proteins EGL-1 and CED-4. Mol. Cell, 2004; 15: 999-1006
[PubMed] [Full Text HTML] [Full Text PDF]
[74] Yan N., Xu Y., Shi Y.: 2:1 Stoichiometry of the CED-4-CED-9 complex and the tetrameric CED-4: insights into the regulation of CED-3 activation. Cell Cycle, 2006; 5: 31-34
[PubMed] [Full Text PDF]
[75] Yang X., Chang H.Y., Baltimore D.: Essential role of CED-4 oligomerization in CED-3 activation and apoptosis. Science, 1998; 281: 1355-1357
[PubMed]
[76] Yin Q., Park H., Chung J.Y., Lin S., Lo Y., da Graca L.S., Jiang X., Wu H.: Caspase-9 holoenzyme is a specific and optimal pro-caspase-3 processing machine. Mol. Cell, 2006; 22: 259-268
[PubMed] [Full Text HTML] [Full Text PDF]
[77] Yu X., Acehan D., Ménétret J.F., Booth C.R., Ludtke S.J., Riedl S.J., Shi Y., Wang X., Akey C.W.: A structure of the human apoptosome at 12.8 A resolution provides insights into this cell death platform. Structure, 2005; 13: 1725-1735
[PubMed] [Full Text HTML] [Full Text PDF]
[78] Yu X., Wang L., Acehan D., Wang X., Akey C.W.: Three-dimensional structure of a double apoptosome formed by the Drosophila Apaf-1-related killer. J. Mol. Biol., 2006; 355: 577-589
[PubMed]
[79] Yuan S., Yu X., Topf M., Dorstyn L., Kumar S., Ludtke S.J., Akey C.W.: Structure of the Drosophila apoptosome at 6.9 A resolution. Structure, 2011; 19: 128-140
[PubMed] [Full Text HTML] [Full Text PDF]
[80] Yuan S., Yu X., Topf M., Ludtke S.J., Wang X., Akey C.W.: Structure of an apoptosome procaspase-9 CARD complex. Structure, 2010; 18: 571-583
[PubMed] [Full Text HTML] [Full Text PDF]
[81] Zech B., Kohl R., von Knethen A., Brune B.: Nitric oxide donors inhibit formation of the Apaf-1/caspase-9 apoptosome and activation of caspases. Biochem. J., 2003; 371: 1055-1064
[PubMed] [Full Text HTML] [Full Text PDF]
[82] Zlobec I., Minoo P., Baker K., Haegert D., Khetani K., Tornillo L., Terracciano L., Jass J.R. Lugli A.: Loss of APAF-1 expression is associated with tumour progression and adverse prognosis in colorectal cancer. Eur. J. Cancer, 2007; 43: 1101-1107
[PubMed]
Autor deklaruje brak potencjalnych konfliktów interesów.