
Udział laktoferryny w gospodarce żelazem w organizmie. Część II. Działanie przeciwmikrobiologiczne i przeciwzapalne poprzez sekwestrację żelaza
Jolanta Artym 1Streszczenie
Laktoferryna (LF) jest glikoproteiną powszechnie występującą w organizmie ssaków. Jest wytwarzana przez komórki nabłonkowe i stąd obecna w wydzielinach na powierzchni błon śluzowych. Znajduje się też w ziarnistościach granulocytów obojętnochłonnych, skąd podczas urazu, infekcji i zapalenia jest uwalniana do krwi. LF należy do rodziny transferryn – białek wiążących jony żelaza z dużym powinowactwem. Zdolność do wiązania żelaza to jedna z najwcześniej odkrytych właściwości białka. LF wyizolowano z mleka w latach 60. ub. wieku i nazwano „czerwonym białkiem wiążącym żelazo”. Ze zdolnością do chelatowania żelaza wiążą się inne funkcje, jakie białko pełni w ustroju. Jak przedstawiono w I części artykułu [14] LF uczestniczy w pobieraniu żelaza z pożywienia i jego magazynowaniu, a w pewnym stopniu również w transporcie do komórek (choć tu główną rolę należy przypisać transferrynie).
W II części artykułu omówiono wpływ LF na zwalczanie mikroorganizmów w wyniku chelatowania żelaza, przez co pierwiastek ten staje się niedostępny dla komórek patogenów, ograniczając ich wzrost. Żelazo poprzez udział w wielu procesach metabolicznych staje się niezbędnym składnikiem odżywczym dla niemal wszystkich drobnoustrojów. Dla patogenów żelazo nie jest łatwo dostępne, gdyż organizm żywiciela pilnie strzeże swoich zapasów. Ponieważ żelazo jest niezbędne do prawidłowego funkcjonowania zarówno drobnoustroju, jak i organizmu gospodarza, nabywanie żelaza przez patogen podczas infekcji jest uznawane za ważny czynnik wirulencji. Organizm wyższy wykształcił skomplikowany system ochrony zapasów żelaza, a jednym z jego istotnych elementów jest laktoferryna. Białko w postaci ubogiej w żelazo skutecznie zwalcza wiele bakterii oraz grzybów, działając mikrobiostatycznie lub bójczo. Stopień wysycenia żelazem wpływa również na aktywność przeciwwirusową LF. Niektóre patogeny (m.in. Helicobacter pylori, Neisseria sp., Haemophilus influenzae) wykształciły system sideroforów lub receptorów komórkowych, dzięki któremu mogą nabywać żelazo z LF i transferryny. Tak zwana laktoferrynowa teoria hipoferremii w zapaleniu zakłada ponadto ochronny udział białka w stanach zapalnych, sepsie i urazach. LF poprzez wychwytywanie i gromadzenie żelaza osoczowego w wątrobie i śledzionie czasowo obniża jego dostępność dla drobnoustrojów i procesów tworzenia toksycznych reaktywnych form tlenu, co przyczynia się do wygaszania reakcji zapalnych.
Słowa kluczowe:laktoferryna • transferryna • ferrytyna • chelatowanie żelaza • zwalczanie drobnoustrojów • właściwości przeciwzapalne
Summary
Lactoferrin (LF) is a glycoprotein widely distributed in mammalian organisms. It is synthesized by epithelial cells; hence it is present in secretions of mucous membranes. It is also contained in secondary granules of neutrophils and released to the circulation during trauma, infection or inflammation. LF belongs to the transferrin family – proteins binding iron ions with a high affinity. Upon isolation in 1961 LF was initially called a red, iron-binding protein. LF’s ability to bind iron is associated with other functions which the protein fulfils in the body. As described in the first part of the article [14], LF participates in acquisition of iron from food and its storage in the body, and to a certain degree also in iron transport to cells. In this part of the article the effect of LF in combating microorganisms by chelating iron is described. The iron-chelating property of LF renders iron inaccessible to the pathogens, thus restricting their growth. Iron, due to its participation in many metabolic processes, is an essential element for almost all microorganisms. Iron is not easily accessible for pathogens within the host. Since iron is crucial for normal function of both pathogens and the host, an ability to acquire iron during infection is regarded as an important virulence factor. Higher vertebrates have evolved a complicated protection system of iron storage and LF is an important element of this system. Low iron-saturated LF effectively combats bacteria and fungi, acting in a bacteriostatic and fungistatic way. The degree of iron saturation also influences antiviral activity of LF. Some pathogens (e.g. Helicobacter pylori, Neisseria sp, Haemophilus influenzae) have evolved a system of siderophores or cellular receptors which can acquire iron from LF and transferrin. The so-called lactoferrin theory of hypoferremia in inflammation assumes, in addition, a protective role of the protein in inflammation, sepsis and trauma. LF, by chelation and storage of plasma iron in the liver and spleen, temporarily restricts its accessibility for microorganisms and processes of formation of toxic, reactive oxygen species, which contributes to the amelioration of inflammatory states.
Key words:lactoferrin • transferrin • ferritin • iron chelation • antimicrobial activity • antiinflammatory activity
Wykaz skrótów:
apo-LF – laktoferryna wolna od żelaza; BLF – laktoferryna bydlęca; DFO – desferoksamina; HLF – laktoferryna ludzka; holo-LF – laktoferryna wysycona żelazem; LF – laktoferryna; Mn-LF – laktoferryna wysycona manganem; RFT – reaktywne formy tlenu; rHLF – rekombinacyjna laktoferryna ludzka; TF – transferryna; Zn-LF – laktoferryna wysycona cynkiem.
Wstęp
Laktoferryna (LF) odkryta po raz pierwszy w mleku ponad 70 lat temu to plejotropowa glikoproteina o zdolności wiązania żelaza, należąca do rodziny transferryn. Powszechnie występuje w wydzielinach egzokrynnych ssaków (głównie mleku, łzach, wydzielinie przewodu pokarmowego, dróg oddechowych, dróg moczowo-płciowych), do których jest uwalniana przez komórki nabłonkowe. LF jest ponadto składnikiem wtórnych ziarnistości granulocytów obojętnochłonnych, które stanowią główne źródło LF we krwi oraz miejscach objętych zapaleniem. LF jest istotnym składnikiem systemu odporności gospodarza. Zwalcza drobnoustroje na powierzchni śluzówek, przez ograniczanie namnażania oraz bezpośrednie zabijanie. Od dawna wiadomo, że te właściwości białka są związane w dużej części ze zdolnością do sekwestracji koniecznego do wzrostu mikroorganizmów żelaza w płynach ustrojowych. Późniejsze badania wykazały dodatkowe niezwiązane z sekwestracją żelaza mechanizmy działania LF na drobnoustroje (m.in. destabilizację osłon komórkowych, wpływ na metabolizm, działanie poprzez wzmocnienie funkcji układu odpornościowego). Podobnie większość właściwości przeciwzapalnych i immunoregulatorowych LF nie wiąże się bezpośrednio ze zdolnością do chelatowania żelaza, ale obejmuje regulację wytwarzania i aktywności cytokin, chemokin i innych cząsteczek biologicznie aktywnych, a na tej drodze regulację aktywności komórek immunologicznych. Duże znaczenie należy przypisać zdolności wiązania przez dodatnio naładowaną cząsteczkę LF wielu różnych cząsteczek naładowanych ujemnie, m.in. bakteryjnego LPS, heparyny, lizozymu, DNA.
W pierwszej części opracowania [14] omówiono udział LF w pobieraniu z pożywienia i dostarczaniu żelaza do komórek, w których może być magazynowane. Na rąbku szczoteczkowym jelita znaleziono receptory LF, co sugeruje jej rolę w pierwszym z procesów. Receptory LF wykryto też na powierzchni innych komórek, w tym makrofagów, i można przypuszczać, że umożliwiają one transfer żelaza z LF do ferrytyny, białka magazynującego żelazo w komórkach.
Usunięcie wolnego żelaza z jelita, powierzchni śluzówek czy płynów ustrojowych ogranicza dostępność tego pierwiastka dla mikroorganizmów, pozwalając na skuteczną kontrolę flory bakteryjnej. Jednym z głównych elementów tej kontroli u ssaków jest laktoferryna. W artykule omówiono właściwości przeciwbakteryjne, przeciwgrzybicze i przeciwwirusowe oraz przeciwzapalne białka związane z sekwestracją żelaza.
Żelazo jako składnik niezbędny dla drobnoustrojów i organizmów wyższych
Żelazo jest niezbędnym składnikiem odżywczym dla niemalże wszystkich istot żywych: organizmów wyższych i mikroorganizmów. Znaczenie żelaza dla tych pierwszych omówiono w I części artykułu [14]. W komórkach drobnoustrojów żelazo, choć występuje w śladowych ilościach, ma zasadnicze znaczenie dla wielu procesów metabolicznych, w tym obejmujących oddychanie komórkowe, transport tlenu, syntezę DNA, regulację genów, wiązanie azotu, aktywację i uwalnianie wodoru cząsteczkowego, fotosyntezę, metanogenezę. Jest składnikiem wielu enzymów: cytochromów, białek żelazo-siarkowych (np. ferredoksyny), katalazy, peroksydazy oraz aktywatorem hydrogenazy, oksygenazy katecholowej, nitrogenazy, dehydrogenazy tlenku węgla i innych [10].
W świecie drobnoustrojów istnieje stałe współzawodnictwo o żelazo, zarówno między samymi mikroorganizmami, jak i między mikroorganizmami a ich gospodarzami, podczas bytowania komensalnego i infekcji. Ponieważ żelazo jest podstawowym składnikiem odżywczym zarówno dla drobnoustroju, jak i organizmu gospodarza, nabywanie żelaza przez drobnoustrój podczas infekcji jest uznawane za ważny czynnik wirulencji. Istnieje wiele danych epidemiologicznych oraz wyników badań eksperymentalnych potwierdzających związek między statusem żelaza w ustroju a jego podatnością na zakażenia. Na przykład kolonizacja organizmu gospodarza i proliferacja komórek Candida albicans są możliwe jedynie wtedy, gdy w środowisku wzrostu grzybów znajdują się wystarczające ilości żelaza [2,82]. Nadmierne nagromadzenie żelaza w ustroju („iron overload”) stanowiło czynnik ryzyka rozwoju inwazyjnej aspergilozy u pacjentów z nowotworami hematologicznymi [43] oraz po przeszczepach wątroby [3] i szpiku kostnego [5]. Podobne korelacje między zawartością żelaza w ustroju a patogennością drobnoustrojów obserwowano dla: Mycobacterium tuberculosis, Salmonella sp. i Yersinia sp. [27,31,66,75,86] oraz dla patogennych grzybów Cryptococcus neoformans [15]. Chorzy poddani transfuzji krwi przechowywanej przez ponad 3 tygodnie (a zatem zawierającej zlizowane erytrocyty i nadmiar wolnego żelaza) wykazywali zwiększoną podatność na ciężkie zakażenia Yersinia enterocolitica [49]. Oczywiste jest zatem, że zawartość żelaza w ustroju gospodarza wpływa na jego podatność na infekcje, a metody pozbawiające środowisko wolnego żelaza to istotne naturalne mechanizmy obronne organizmów wyższych.
Z punktu widzenia obronności ustroju wydawać by się mogło, że najlepszym rozwiązaniem byłoby mocne związanie żelaza i tym samym usunięcie go ze środowiska wzrostu drobnoustrojów. Nie jest to jednak takie proste. Należy bowiem pamiętać, że żelazo w ustroju jest nie tylko potencjalnym składnikiem odżywczym dla komórek patogenów, ale też pełni wiele ważnych funkcji, w tym funkcję kofaktora tworzenia reaktywnych form tlenu (RFT), które uczestniczą w zabijaniu drobnoustrojów. I tak np. makrofagi wymagają obecności wystarczających ilości żelaza wewnątrzkomórkowego do indukcji skutecznych mechanizmów zwalczania drobnoustrojów, obejmujących zależny od oksydazy NADPH wybuch tlenowy i wytwarzanie tlenku azotu (NO•) katalizowane przez indukowalną syntazę NO• (iNOS) [53]. Tego samego żelaza potrzebują pasożytujące wewnątrzkomórkowo bakterie, takie jak M. tuberculosis i Salmonella typhimurium [27]. W tej sytuacji organizm ogranicza ilość żelaza w aktywowanych makrofagach, obniżając ekspresję receptora transferryny (TF) na ich powierzchni [24,72], co jednocześnie może jednak ograniczać zabijanie sfagocytowanych komórek. Jak więc widać, równowaga między ilością żelaza dostępnego dla drobnoustrojów i potrzebnego do sprawnego funkcjonowania organizmu gospodarza jest bardzo delikatna i płynna. Potwierdzeniem są dwie przeciwstawne obserwacje. Pierwsza pochodzi z badań epidemiologicznych, które wykazały, że kobiety ciężarne z anemią z niedoboru żelaza są bardziej narażone na infekcję Helicobacter pylori niż kobiety zdrowe [63]. Podobnie z testów na myszach wynika, że podanie chelatora żelaza – desferoksaminy (DFO) redukowało stężenie żelaza w surowicy i znacznie nasilało przebieg doświadczalnej salmonelozy, czemu towarzyszył spadek wytwarzania H2O2 przez aktywowane makrofagi [27,28]. Zaobserwowano jednak, że również bardziej podatni na zakażenia bakteryjne, w tym bakteriami wewnątrzkomórkowymi są pacjenci z nadmiarem żelaza w ustroju [31,66,75,86].
Jak ustrój chroni zapasy żelaza?
Żelazo jest przedmiotem „walki” między drobnoustrojami a organizmami wyższymi. By pozbawić środowisko wolnego żelaza i uniemożliwić jego pobór przez komórki patogenów organizm wytworzył liczne związki (głównie białka), które tak ściśle wiążą żelazo, by stało się nieosiągalne dla wrogich drobnoustrojów, ograniczając w ten sposób ich wzrost. Są to tzw. związki chelatujące (chelatory; związki mocno chwytające jony metali, jakby szczypcami, „chele” z grec. szczypce).
Do związków tego typu zaliczamy: kanalbuminę (w białku jaja kurzego) oraz u ssaków: transferrynę (w osoczu krwi), laktoferrynę (w wydzielinach nabłonkowych, neutrofilach i osoczu krwi) oraz ferrytynę (w przestrzeni wewnątrzkomórkowej). W wydzielinach i osoczu obecne są zarówno TF, jak i LF, jednak ta pierwsza przeważa we krwi (2-4 mg/ml), a druga jest powszechna w wydzielinach. Właściwości LF jako białka wiążącego żelazo omówiono w I części artykułu [14]. Tu warto przypomnieć, że LF wiąże z dużym powinowactwem (Kd rzędu 10-20 M) dwa jony żelazowe (Fe3+) i z mniejszym jony żelazawe (Fe2+). Podobnie TF wiąże dwa jony Fe3+, ale z około 360 razy mniejszym powinowactwem. Pierwsze z białek uwalnia jony żelaza w kwaśnym środowisku (pH poniżej 3,5), a drugie już przy słabym zakwaszeniu (pH~6) [1]. TF jest głównym białkiem transportowym żelaza, mimo to jedynie około 3 mg tego pierwiastka (0,1% całkowitej zawartości w organizmie) są związane z TF [37]. U zdrowych ludzi 20-30% cząsteczek TF i 5-8% cząsteczek LF jest wysyconych żelazem, reszta zatem pozostaje gotowa do wyłapywania i wiązania jonów żelaza, które znajdą się w osoczu. Dzięki temu ilość wolnego żelaza w osoczu krwi jest bardzo mała i wynosi około 10-18 M [22], jest zatem 108-1010 razy mniejsza niż wymagana do wzrostu drobnoustrojów (10-7-10-5 M) [10]. Gdy zdolność TF do wiązania żelaza zostanie przekroczona (np. u chorych na hemochromatozę – chorobę dziedziczną przebiegającą z nadmiernym wchłanianiem żelaza), żelazo jest wiązane (ale znacznie słabiej) z albuminą, cytrynianem i innymi związkami niskocząsteczkowymi [16].
Białkiem odpowiedzialnym za sekwestrowanie wewnątrzkomórkowego żelaza jest ferrytyna, która utrzymuje homeostazę tego pierwiastka w komórce, chroniąc ją przed toksycznymi reakcjami katalizowanymi przez żelazo [83]. W warunkach fizjologicznych ferrytyna jest zwykle wysycona żelazem zaledwie w 20%. Z tego wewnątrzkomórkowego magazynu żelazo jest dostarczane do różnego rodzaju procesów metabolicznych przebiegających z jego udziałem. Ferrytyna jest wyjątkowo stabilną cząsteczką, jej czwartorzędowa struktura dysocjuje dopiero w bardzo kwaśnym środowisku (przy pH <2), a odnawia się, gdy pH wraca do wartości 7,4 [51]. Żelazo z ferrytyny uwalniane jest w wyniku redukcji, w środowisku o kwaśnym odczynie. W komórce istnieje również przejściowa pula słabo związanego żelaza aktywnego w procesach redoks, która wiąże ze sobą etapy: nabywania żelaza z TF, przechowywania w postaci ferrytyny oraz wykorzystania w procesach metabolicznych. To tzw. pula labilnego żelaza (labile iron pool – LIP) i jej stężenie w cytoplazmie komórek ssaków nie przekracza 1 µM, co stanowi jedynie małą frakcję (<5%) całkowitego żelaza komórkowego (50-100 µM) [46]. W przestrzeni cytoplazmatycznej komórki żelazo przeważnie występuje w postaci rozpuszczalnych jonów Fe+2, a w przestrzeni zewnątrzkomórkowej – w postaci nierozpuszczalnych, czyli trudniej dostępnych, jonów Fe+3. Ustrój poza związkami chelatującymi żelazo ma również inne sposoby ograniczania dostępu patogenów do żelaza, uruchamiane podczas infekcji. I tak podniesiona temperatura ciała obniża stężenie żelaza we krwi, co sprzyja wygaszaniu infekcji [23]. Niektóre cytokiny (IFN-γ, IL-1, IL-6, TNF-α) zmniejszają ekspresję receptora TF na powierzchni makrofagów, co uniemożliwia pobór żelaza do komórki i ogranicza możliwość wzrostu patogenów wewnątrzkomórkowych [24,72].
Najbardziej powszechną postacią żelaza w ustroju jest żelazo hemowe, które jest potencjalnym źródłem żelaza jonowego we krwi i innych płynach pozakomórkowych. Dlatego nie dziwi, że organizm ściśle chroni swoje zapasy żelaza hemowego, które może być nie tylko wykorzystane przez drobnoustroje, ale też może prawdopodobnie katalizować tworzenie RFT [16]. Organizm wykształcił białka: haptoglobinę i hemopeksynę, silnie wiążące odpowiednio: wolną hemoglobinę oraz hem [10]. Słabsze zdolności wiążące ma również albumina osocza krwi.
Jak drobnoustroje walczą o żelazo?
Zważywszy na bardzo niewielkie ilości dostępnego żelaza w organizmie gospodarza, nie dziwi, że drobnoustroje „opracowały” specjalne strategie służące pozyskiwaniu tego cennego składnika odżywczego. Wiele patogenów wydziela siderofory – substancje wychwytujące żelazo i dostarczające je do komórki. Są to prawie zawsze związki niskocząsteczkowe (<1 kDa), rozpuszczalne w wodzie i wiążące żelazo (Fe+3) swoiście z dużym powinowactwem (Kd~10-30 M) [10]. Na przykład niektóre bakterie jelitowe wytwarzają enterobaktynę (enterochelinę), która w postaci wolnej jest uwalniana do podłoża, gdzie wiąże żelazo, a komórka pobiera i rozkłada powstały kompleks, uwalniając żelazo. Wiele grzybów wytwarza tzw. ferrichromy, wiążące żelazo nierozpuszczalne (Fe+3) z podłoża, które wewnątrz komórki jest redukowane do postaci rozpuszczalnej (Fe+2), do której siderofor ma niewielkie powinowactwo. Ze względu, iż powinowactwo niektórych sideroforów do żelaza jest nawet wyższe niż powinowactwo białek ustrojowych, siderofory mogą z nich „odbierać” żelazo. Dotyczy to nawet tak silnych chelatorów żelaza jak: ferrytyna, TF czy LF. Inny system pobierania żelaza, który wykształciły niektóre bakterie, to specjalne białka powierzchniowe służące do wiązania chelatorów gospodarza i bezpośredniego nabywania z nich żelaza [10]. W ten sposób pierwiastek pozyskują m.in.: H. pylori, Haemophilus influenzae, Neisseria sp. i Moraxella sp. Niektóre drobnoustroje wykształciły jednocześnie kilka sposobów na zdobycie żelaza w organizmie żywiciela. Ciekawym przykładem może być komensalny i oportunistyczny drożdżak – C. albicans. Według dotychczasowej wiedzy, grzyb ten nie wytwarza wprawdzie własnych sideroforów, ale potrafi wykorzystywać żelazo skompleksowane z sideroforami innych mikroorganizmów. Wykorzystuje ponadto hemoglobinę, ferrytynę i transferrynę jako źródła żelaza, a po inwazji i destrukcji tkanek także żelazo z labilnej puli cytoplazmatycznej, białek żelazo-siarkowych i innych białek niehemowych [4]. Białka hemowe (np. hemoglobina, mioglobina, cytochromy) mogą stanowić źródło żelaza dla patogenów, ale ogólnie są dla nich trudno dostępne ze względu na swoje umiejscowienie (są chronione wewnątrz komórek). Stają się jednak dostępne po uszkodzeniu tkanek i uwolnieniu zawartości komórek, co sprzyja nasileniu infekcji.
Zwalczanie drobnoustrojów poprzez sekwestrację żelaza
Laktoferryna jest aktywna wobec wielu bakterii Gram-ujemnych i Gram-dodatnich, grzybów, wirusów oraz pasożytów [45]. Zawarta w wydzielinach śluzówek stanowi ważny czynnik usuwający drobnoustroje już „u wrót” zakażenia – na powierzchni błon śluzowych, zapobiegając tym samym ich inwazji do organizmu. W miejscach infekcji/urazu stężenie LF może dodatkowo wzrosnąć po degranulacji naciekających granulocytów. Na przykład stężenia LF w ślinie osób zdrowych wynoszą średnio 5 µg/ml, w chorobach przyzębia wzrastają do 12 µg/ml, a po naświetlaniach w radioterapii aż do 80 µg/ml [40]. Granulocyty obojętnochłonne są również źródłem LF w osoczu krwi. Dzienne wytwarzanie LF w organizmie zdrowego człowieka można oszacować na około 5 g (średnia zawartość LF w 106 neutrofilów to 5 µg, dziennie wytwarzanych jest 1012 neutrofilów), z czego jedynie 10% (0,5 g) jest uwalniane z komórek podczas prawidłowego ich obrotu. Podczas zakażenia bakteryjnego obrót neutrofilów wzrasta nawet 50-krotnie, czemu towarzyszy 3-krotnie większa destrukcja tych komórek. W tym czasie wytwarzanie LF wzrasta do 30 g/dobę, z czego 1/3 (10 g) jest uwalniana do krwiobiegu [70]. Stężenie LF w osoczu krwi dość stałe w zdrowiu (średnio 0,3 µg/ml) [17], wzrasta podczas infekcji, zapalenia oraz rozległych urazów. U chorych z płatowym zapaleniem płuc to 1-2 µg/ml, a u pacjentów z ciężkimi oparzeniami 10-40 µg/ml [17,93].
Hamowanie wzrostu drobnoustrojów przez chelatowanie wolnego żelaza to jedna z najwcześniej opisanych właściwości LF. Już w 1966 roku (a więc niedługo po odkryciu białka) Masson i wsp. zauważyli, że LF wyizolowana z ludzkiego mleka w testach in vitro hamowała wzrost Staphylococcus albus, nie działając na S. aureus [60]. Hamujący wpływ LF był stopniowo znoszony przez dodatek wzrastających ilości jonów żelaza do zastosowanego roztworu LF, co potwierdzało proponowany mechanizm działania białka. W kolejnych latach przeprowadzono wiele testów, które potwierdziły hamujące działanie LF w wyniku sekwestracji jonów żelaza (tabela 1). W ten sposób białko działa na chorobotwórcze bakterie i grzyby. Przeważnie jest to wpływ mikrobiostatyczny, ale czasem również bójczy. Jak należy się domyślać większą aktywność będzie miało białko w postaci wolnej od żelaza (apo-LF) niż wysyconej żelazem (holo-LF). Co ciekawe, stopień wysycenia żelazem (i innymi metalami) cząsteczki LF ma też wpływ na jej aktywność przeciwwirusową.
Tabela 1. Wykaz wrażliwych na działanie LF bakterii, grzybów i wirusów
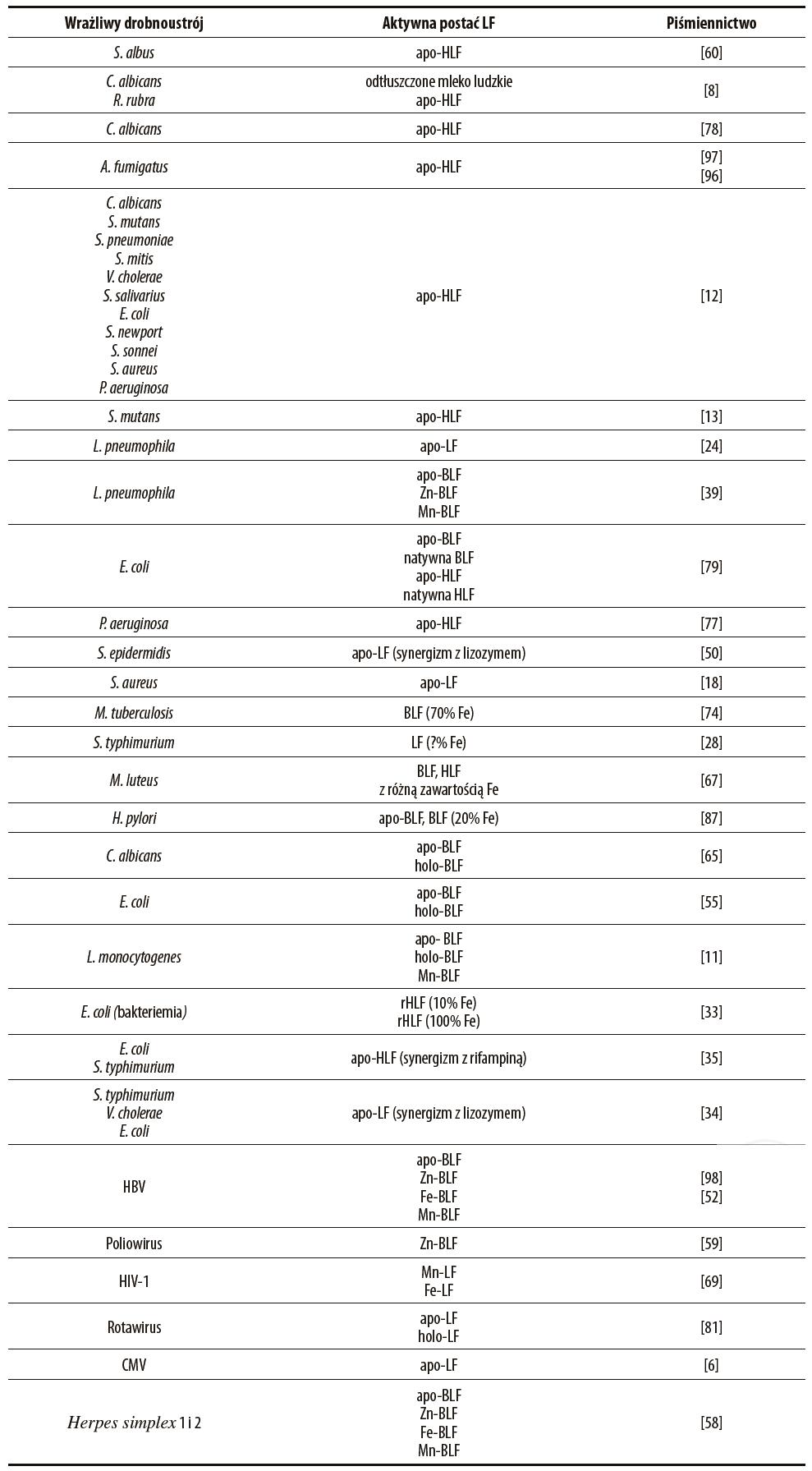
Laktoferryna z ludzkiego mleka hamowała wzrost grzybów C. albicans oraz Rhodotorula rubra, przy czym aktywność ta była związana jedynie z zawartością apo-LF, a dodatek żelaza ją znosił [8]. Działanie białka było zależne od stężenia i czasu. Stwierdzono również hamujący wpływ odtłuszczonego mleka ludzkiego oraz wyizolowanej LF, ale nie innych białek mleka (kazeiny i α-laktoalbuminy). Na podstawie testów żywotności komórek grzybów i mikroskopii elektronowej, autorzy badań podkreślają fungostatyczne (ale nie bójcze) działanie białka. Kolejne badania potwierdziły działanie LF na komórki C. albicans [78]. Wyizolowana z siary ludzka częściowo wysycona i apo-LF (ale nie holo-LF) dodana do hodowli zabijała komórki grzybów w sposób zależny od dawki, czasu i warunków hodowli (temperatura, pH, obecność jonów fosforanowych i węglanowych). Z kolei autorzy tych badań podkreślają wyraźne grzybobójcze działanie białka. Grzybobójcze działanie ludzkiej LF zanotowano również wobec klinicznych izolatów C. albicans i C. krusei [64]. Było ono zależne od dawki i obserwowane tylko dla LF wolnej od żelaza. Badanie z użyciem mikroskopu elektronowego wykazało zmiany na powierzchni komórek, takie jak tworzenie pęcherzyków i wyciek białek, co wskazuje na bezpośrednie działanie toksyczne na komórki grzybów.
Ludzka LF dodana do hodowli hamowała wzrost konidiów Aspergillus fumigatus [97]. Podobną aktywność miały leukocyty wielojądrzaste wyizolowane z krwi osób zdrowych i cierpiących na przewlekłą chorobę ziarniniakową oraz czynnik uwolniony z tych komórek po ich aktywacji. Czynnik ten zidentyfikowano jako LF. Co istotne, wysycenie białka żelazem znosiło jego działanie przeciwgrzybicze, zatem działanie to opierało się na sekwestracji żelaza. Ten sam zespół badał następnie aktywność wobec tych samych grzybów LF oraz innych chelatorów żelaza (DFO, deferipronu, ciclopiroxu) w monoterapii lub połączonych z lekami przeciwgrzybiczymi [96]. LF, deferipron, ciclopirox hamowały, podczas gdy DFO stymulowała wzrost grzybów. Synergistyczne działanie przeciwgrzybicze wykazały: ketokonazol i ciclopirox, flukonazol i deferipron oraz amfoterycyna B i LF. Zatem chelatory żelaza w monoterapii lub w terapii kombinowanej z antybiotykami przeciwgrzybiczymi mogą stanowić ważny element profilaktyki i leczenia aspergilozy, groźnej u pacjentów z osłabioną odpornością.
Arnold i wsp. w testach in vitro wykazali aktywność ludzkiej LF pochodzącej z siary wobec wiele gatunków bakterii oraz C. albicans [12]. LF wolna od żelaza (ale nie holo-LF) silnie hamowała (nawet o 99%) wzrost szczepów laboratoryjnych: Streptococcus mutans, S. pneumoniae oraz Vibrio cholerae. Zanotowano ponadto hamowanie wzrostu szczepów izolowanych od pacjentów: S. salivarius, S. mitis i C. albicans (izolaty z płytek nazębnych) oraz E. coli, P. aeruginosa, Salmonella newport, Shigella sonnei i S. aureus (izolaty z różnych tkanek). Stwierdzono prawidłowość, że szczepy wirulentne są mniej wrażliwe na działanie LF, podobnie szczepy laboratoryjne po pasażu na myszach odznaczały się mniejszą wrażliwością. Dalsze badania zespołu Arnolda wykazały, że ludzka apo-LF zabijała komórki S. mutans wiążąc się do ich powierzchni i nieodwracalnie hamując ich metabolizm [13]. Internalizacja apo-LF do monocytów zakażonych Legionella pneumophila prowadziła do zahamowania wzrostu bakterii poprzez chelatowanie żelaza, przeciwnie – internalizowana holo-LF hamowała przeciwbakteryjną aktywność IFN-γ oraz była źródłem żelaza do wzrostu patogenu [24]. Apo-BLF oraz BLF wysycona Zn i Mn hamowały, podczas gdy LF wysycona żelazem promowała wewnątrzkomórkowe namnażanie L. pneumophila w nieswoistych fagocytach (komórki linii HeLa) [39]. Autorzy cytowanych badań nie stwierdzili wprawdzie internalizacji komórkowej Fe-BLF, ale można przypuszczać, że białko ulegało jednak endocytozie do komórek, gdzie uwalniało Fe, które po włączeniu do labilnej puli żelaza komórkowego mogło być wykorzystane przez bakterie.
LF wyizolowana z mleka ludzkiego i krowiego w postaci apo- i natywnej (wysyconej żelazem w kilku%) w obecności immunoglobulin IgA lub IgG hamowała namnażanie Escherichia coli w hodowli. Bakteriostaza była zniesiona przez wysycenie białka żelazem [79]. Aktywność hamującą tworzenie biofilmu Pseudomonas aeruginosa miała wyłącznie postać apo-HLF, wysycenie żelazem ją znosiło [77]. Poprzez sekwestrację żelaza LF stymulowała ruch drgający bakterii, dzięki czemu bakterie przemieszczały się po podłożu i nie tworzyły kolonii i biofilmu. Biofilm jest zwartą i trwałą strukturą tworzoną przez niektóre bakterie na powierzchni nabłonków w przewlekłych infekcjach.
Biofilmy odznaczają się większą opornością na mechanizmy obronne gospodarza i leczenie antybiotykami. Do tworzenia biofilmów wymagane są wyższe stężenia żelaza, bo tu dostęp poszczególnych komórek do tego składnika jest ograniczony [90]. W cytowanych badaniach chelatowanie żelaza przez LF zmniejszało również oporność bakterii na czynniki bójcze: H2O2 i antybiotyk tobramycynę [77]. Apo-LF działała synergistycznie z lizozymem w hamowaniu wzrostu Staphylococcus epidermidis. Wysycenie białka żelazem znosiło jego aktywność [50]. Synergizm działania LF i lizozymu może mieć znaczenie ochronne u osób noszących szkła kontaktowe. S. epidermidis łatwo kolonizuje powierzchnie różnego rodzaju protez i implantów, ale rzadko powierzchnię soczewek kontaktowych, choć są wytwarzane z podobnych materiałów. Można sądzić, że właśnie lizozym i słabo wysycona żelazem LF odpowiadają przynajmniej w części za obserwowany efekt. Apo-LF hamowała wzrost S. aureus w testach in vitro, podczas gdy białko wysycone żelazem nie miało takiej aktywności [18]. Co ciekawe, w testach in vivo podana myszom apo-LF była tak samo skuteczna jak białko wysycone w różnym stopniu żelazem, co wskazuje, że w tym przypadku aktywność białka nie zależała wyłącznie od sekwestracji żelaza [18].
Interesujące wyniki uzyskano w testach na mysim modelu gruźlicy. Zastosowano myszy z defektem w metabolizmie żelaza (nokaut genu β2-mikroglobuliny), którym podawano dodatkowe ilości żelaza w wodzie pitnej, osiągając stan „przeładowania” organizmu żelazem, co istotnie nasilało przebieg zakażenia M. tuberculosis [74]. Podana donosowo BLF wyraźnie redukowała rozsiew bakterii w narządach wewnętrznych w procesie sekwestracji żelaza (choć użyto białka wysyconego żelazem aż w 70%). LF podnosiła ponadto surowicze stężenia NO• (wytwarzanie tego mediatora jest zaburzone podczas „przeładowania” żelazem). W testach in vitro wykazano natomiast, że LF obniża ekspresję receptora TF na makrofagach, co ogranicza pobieranie żelaza przez te komórki.
Ciekawe obserwacje pochodzą z cytowanych już badań Collins i wsp. nad namnażaniem innych bakterii wewnątrzkomórkowych – S. typhimurium [28]. Badacze ci używali DFO (siderofor wytwarzany przez Streptomyces) oraz LF jako chelatorów żelaza w testach in vitro oraz in vivo. Podanie LF myszom przed zakażeniem w pewnym stopniu (choć nieistotnie) łagodziło przebieg infekcji, podczas gdy podanie DFO znacznie ją nasilało. Co istotne, LF znacznie stymulowała tworzenie H2O2 oraz zabijanie bakterii przez aktywowane makrofagi, drugi z chelatorów osłabiał właściwości bójcze fagocytów i zmniejszał wybuch tlenowy. Indukowane obecnością żelaza wytwarzanie H2O2 z udziałem oksydazy NADPH jest zasadniczym mechanizmem efektorowym w kontroli infekcji Salmonella. Uzyskane dane wyraźnie wskazują, że przyczyną zaostrzenia infekcji było ograniczenie dostępności żelaza dla procesów tworzenia RFT. Jak konkludują autorzy, DFO wchodzi do komórek, wiąże zatem zarówno żelazo zewnątrz- jak i wewnątrzkomórkowe, tymczasem LF chelatuje jedynie żelazo na zewnątrz komórek, pulę wewnątrzkomórkową zostawiając nienaruszoną, co pozwala na efektywne tworzenie bakteriobójczych RFT.
W niektórych testach okazało się, że LF wysycona Fe w różnym stopniu działała podobnie skutecznie na drobnoustroje. Ludzka i bydlęca LF powodowały aglutynację komórek Micrococcus luteus, niezależnie od obecności jonów żelaza w medium i stopnia wysycenia cząsteczek [67]. W testach na myszach wolna od żelaza i wysycona żelazem w 20% BLF były tak samo skuteczne w hamowaniu kolonizacji żołądka przez H. pylori [87]. Bydlęca apo- i holo-LF dodane do zawiesiny neutrofilów mysich lub ludzkich zwiększały ich działanie wobec komórek C. albicans [65]. W testach in vitro zanotowano hamowanie przez apo- i holo-BLF adhezji i internalizacji komórek E. coli do komórek nabłonkowych linii HeLa [55]. W podobnych testach bydlęca LF w postaci wolnej od jonów oraz wysyconej manganem lub żelazem hamowała internalizację komórek L. monocytogenes do komórek nabłonkowych linii HT-29 oraz Caco-2 [11]. W zapobieganiu bakteriemii u szczurzych noworodków doustna terapia rHLF była równie efektywna, gdy białko podawano w postaci ubogowysyconej (około 10%), jak i całkowicie wysyconej żelazem (rHLF+FeSO4) [33].
Podobna siła działania postaci wysyconych i niewysyconych metalem przemawia za dodatkowymi, innymi niż chelatowanie żelaza sposobami działania LF na drobnoustroje. Wiadomo, że białko może m.in. bezpośrednio uszkadzać ściany komórek bakteryjnych i grzybowych oraz zaburzać ich metabolizm, hamować procesy adhezji patogenów do tkanek czy rozkładać enzymatycznie czynniki wirulencji drobnoustrojów. Działanie pośrednie białka obejmuje aktywację różnych składników układu odpornościowego gospodarza (m.in. komórek NK, makrofagów, limfocytów) i wzmocnienie ich działania wobec patogenów. Należy jednak pamiętać, że stopień wysycenia żelazem cząsteczki LF poprzez zmianę konformacyjną jej struktury [7] może m.in. wpływać na zdolność do interakcji z niektórymi powierzchniowymi cząsteczkami na komórkach gospodarza [21,56], jak i drobnoustrojów [11,13]. Stąd odmienna siła działania różnych postaci białka w sytuacjach, które nie dotyczą bezpośrednio pozbawienia patogenów dostępu do żelaza, jako cennego składnika odżywczego.
Ilustracją powyższych rozważań może być obserwacja, że stopień wysycenia białka żelazem ma znaczenie w bezpośrednim uszkadzającym działaniu na ścianę komórkową bakterii. Apo-HLF dodana do hodowli bakterii Gram-ujemnych: E. coli i S. typhimurium działała bakteriostatycznie oraz zwiększała bakteriobójcze działanie antybiotyku ryfampiny [35]. Aktywność LF była znoszona przez wysycenie żelazem. Mechanizm działania LF polegał na uszkadzaniu struktury ściany komórkowej bakterii poprzez uwalnianie cząsteczek lipopolisacharydu (LPS) i zmianę przepuszczalności. Jak wiadomo, anionowe cząsteczki LPS są stabilizowane w obrębie ściany komórkowej przez jony dodatnie (wapnia, magnezu i żelaza) [29]. Przez wiązanie tych jonów LF zaburza zatem stabilność tej struktury. Co ważne, może w ten sposób ułatwiać dostęp lizozymu do komórek bakterii Gram-ujemnych i zwiększać jego aktywność bakteriobójczą. W badaniach Ellisona i Ghiela LF i lizozym użyte osobno działały bakteriostatycznie wobec V. cholerae, S. typhimurium i E. coli [34]. Użyte razem wykazywały zależny od dawki efekt bakteriobójczy, który był znoszony przez wysycenie LF żelazem oraz dodatek jonów wapnia do medium. LF, poza jonami żelaza może wiązać również jony innych metali (m.in. manganu, miedzi, cynku i wapnia) [20,71]. Obecność nadmiaru wspomnianych jonów spowodowała „zapełnienie” cząsteczki LF i utratę jej aktywności.
Warto w końcu zwrócić uwagę na aktywność przeciwwirusową LF zależną od stopnia wysycenia żelazem i innymi jonami. Różne postaci bydlęcej LF (niewysycona jonami, Fe-BLF, Mn-LF oraz Zn-BLF) hamowały namnażanie wirusa zapalenia wątroby typu B (HBV) na wrażliwej linii komórkowej, choć największą aktywność zanotowano dla Zn-BLF [52,98]. Podobnie białko wysycone cynkiem najskuteczniej hamowało infekcję komórek poliowirusem [59], a żelazo i mangan z kolei zwiększały aktywność natywnej LF wobec HIV [69]. Apo- i holo-LF w testach in vitro hamowały małpi rotawirus, choć postać niewysycona żelazem odznaczała się znacznie większą aktywnością i mniejszą toksycznością [81]. LF wysycona żelazem traciła zdolność do hamowania infekcji cytomegalowirusem (CMV) ludzkich fibroblastów [6]. Bydlęca LF w różnych postaciach (apo-LF, Zn-LF, Fe-LF, Mn-LF) hamowała replikację ludzkich wirusów Herpes simplex 1 i 2 we wrażliwej linii komórkowej. Wskaźnik ID50 dla LF wysyconej żelazem w 10% wynosił 28 µg/ml, podczas gdy dla LF wysyconej w 90% – tylko 11,5 µg/ml, postać holo-LF była zatem bardziej aktywna niż postać apo-LF. Metale w postaci cytrynianów nie miały dużego wpływu hamującego, można zatem wnioskować, że działanie LF nie było wynikiem dostarczania jonów metali do komórek [58].
Odmienna aktywność przeciwwirusowa różnych postaci LF może się wiązać, jak już wspomniano, z różnym powinowactwem do receptorów niektórych komórek, np. nabłonka przewodu pokarmowego [56] i monocytów/makrofagów [21]. Jednym z mechanizmów aktywności przeciwwirusowej LF jest bowiem wiązanie i blokowanie cząsteczek powierzchniowych na komórkach, tak by nie mogły być wykorzystane przez cząsteczki wirusa [57]. Spotykana w niektórych przypadkach większa aktywność postaci Zn-LF może wynikać z dostarczania cynku do komórek docelowych. Zwiększona dostępność jonów Zn2+ jest prawdopodobnie przyczyną uszkodzonej replikacji niektórych wirusów [36,44]. Z kolei większą aktywność postaci apo-LF obserwowaną wobec niektórych wirusów można prawdopodobnie tłumaczyć bardziej skuteczną sekwestracją jonów metali (głównie żelaza), które są kofaktorami wielu enzymów komórkowych wykorzystywanych przez wirusy oraz enzymów wirusowych, czego skutkiem będzie ograniczenie replikacji wirusów. Dokładny mechanizm antywirusowej aktywności różnych postaci LF wciąż oczekuje na wyjaśnienie.
Laktoferryna jako źródło żelaza dla drobnoustrojów
Jak już wspomniano, LF podobnie jak inne ustrojowe chelatory żelaza, może być źródłem tego pierwiastka dla niektórych patogenów. Badania wykazały, że holo-LF może stanowić źródło żelaza dla bakterii: H. pylori [42], H. influenzae [85], Legionella pneumophila [25], Aeromonas hydrophila [80], Bordetella pertussis [61], E. coli [19] oraz Moraxella sp. i Neisseria sp. [26,62]. Niektóre z tych bakterii rozwinęły na swojej powierzchni specjalne białka, które służą do wiązania białek gospodarza sekwestrujących żelazo i bezpośredniego nabywania tego pierwiastka. Na przykład w przypadku H. pylori jest to białko o masie 70 kDa, które pojawia się na powierzchni bakterii, gdy rosną one w medium pozbawionym żelaza. Potwierdzono ponadto ekspresję białka na szczepach H. pylori wyizolowanych od pacjentów [30]. Większość szczepów H. influenzae wyizolowanych z plwociny chorych na chroniczne zapalenie oskrzeli oraz z gardła osób zdrowych wiązała ludzką LF, wykorzystując do swojego wzrostu zawarte w niej żelazo [85]. Na powierzchni tych bakterii wykazano obecność białek receptorowych o masie 58 i 105/106 kDa wiążących odpowiednio TF i LF [76]. Stwierdzono dużą swoistość gatunkową wspomnianych białek bakteryjnych, co oznacza, że mogą one pozyskiwać żelazo wyłącznie z LF i/lub TF ludzkiej. Na przykład komórki H. pylori nie pobierały żelaza z bydlęcej LF, choć wykazano, że białko to w pewnym stopniu może być wiązane do białek receptorowych bakterii [30]. Ekspresja receptorów na powierzchni komórek bakteryjnych jest indukowana przez ograniczenie dostępności żelaza w środowisku wzrostu [10].
Inne bakterie mogą natomiast pozyskiwać żelazo z LF lub TF za pomocą sideroforów. Dla przykładu A. hydrophila (czynnik etiologiczny biegunki i posocznicy u osób z obniżoną odpornością) wytwarza amonabaktynę, która pobiera żelazo z TF i LF [80]. Stwierdzono również pozyskiwanie żelaza z tych białek przez enterochelinę jelitowych szczepów E. coli [19]. Siderofor wytwarzany przez P. aeruginosa (piowerdyna) może pobierać żelazo z TF, choć tylko w obecności bakteryjnej elastazy. LF nie była źródłem żelaza dla tej bakterii [32].
LF i TF mogą też stanowić źródło żelaza dla niektórych pasożytów, co wykazano dla Trichomonas vaginalis [68] oraz Leishmania chagasi [92]. W pierwszym przypadku białko było wiązane do komórek pasożytów, ale nie ulegało internalizacji. Ekspozycja T. vaginalis na LF znacznie (ponad 6-krotnie) podnosiła ich metabolizm [68]. Pasożyt ten kolonizuje bogatą w LF śluzówkę pochwy, a objawy zakażenia są najbardziej nasilone podczas krwawienia miesięcznego, kiedy stężenia LF i żelaza w wydzielinach pochwy wzrastają. Zakażenie męskiej cewki moczowej, gdzie występuje LF, ale niewiele jest żelaza, ulega samoograniczeniu [88]. Mechanizmy pozyskiwania żelaza z LF przez L. chagasi nie są wyjaśnione. Nie wykazano roli sideroforów ani cięcia proteolitycznego białka. Wiadomo, że pasożyt może nabywać żelazo również z innych źródeł: transferryny i hemu [92].
Jak się okazało, sam organizm może sprzyjać uwalnianiu żelaza z białek chelatujących i udostępnianiu go patogenom. Taką aktywność mają katecholaminy – hormony wydzielane w większych ilościach podczas stresu, do których zalicza się: epinefrynę (adrenalinę), norepinefrynę (noradrenalinę) oraz dopaminę. Inkubacja E. coli z norepinefryną w obecności holo-LF lub holo-TF stymulowała wzrost bakterii poprzez uwalnianie żelaza związanego z białkami [38]. Cząsteczki hormonu tworzyły stabilne kompleksy z LF, TF oraz osoczową albuminą, prowadząc do redukcji związanego jonu Fe3+ do jonu Fe2+ i jego uwolnienia z cząsteczki chelatora [73]. Autorzy badań sugerują znaczenie tych interakcji w klinice, na przykład podczas rozwoju indukowanej stresem sepsy w wyniku nadmiernego rozwoju komensalnych jelitowych szczepów E. coli. Ma to tym większe znaczenie, że związki podobne strukturalnie do amin katecholowych (np. izoprenalina i dobutamina) używane są powszechnie w klinice, m.in. w chorobach układu krążenia.
LF zarówno we krwi, jak i wydzielinach śluzówek występuje w postaci ubogiej w żelazo, nie stanowi więc ważnego źródła pierwiastka dla patogenów. Niemniej jednak w pewnych sytuacjach (podczas zapalenia) może sekwestrować żelazo i wtedy istnieje niebezpieczeństwo, że związane żelazo zostanie wykorzystane przez drobnoustroje. Należy podkreślić, że siderofory wspomnianych bakterii mogą wiązać również żelazo wolne, LF wydaje się zatem nie stanowić dodatkowego zagrożenia, a uwzględniając inne mechanizmy jej działania przeciwbakteryjnego, raczej ogranicza niż sprzyja wzrostowi tych patogenów. Może natomiast ułatwiać wzrost tych bakterii, dla których stanowi jedyne źródło żelaza (nietworzących sideroforów), zwłaszcza wtedy, gdy inne aspekty działania LF nie przeważą tego negatywnego. Cytowane wyniki pochodzą z badań in vitro, są więc konieczne testy in vivo badające „zachowanie” białka w określonych sytuacjach w organizmie i potwierdzające bezpieczeństwo stosowania LF w terapii. W świetle dotychczasowych badań, w schorzeniach wywołanych wymienionymi patogenami, białko (szczególnie ludzkie) powinno być aplikowane z ostrożnością.
Rola laktoferryny w uogólnionych stanach zapalnych
Spadek stężenia żelaza we krwi jest obserwowany podczas infekcji i stanów zapalnych, w tym uogólnionych. Jest to tzw. hipoferremia związana z zapaleniem. Jednocześnie występuje akumulacja tego pierwiastka w magazynach ustrojowych (wątrobie i śledzionie) i jego ograniczone dostarczanie do szpiku kostnego, czemu towarzyszy rozwój anemii (tzw. anemia podczas zapalenia lub chorób chronicznych) [9,89,91]. Jak wskazują niektóre badania, jednym z czynników odpowiedzialnych za to zjawisko może być LF. Zagulski i wsp. przeprowadzili badania przeżywalności królików z sepsą po podaniu E. coli [95]. Bydlęca LF podana dożylnie znacznie przedłużała przeżycie zwierząt, co łączyło się ze spadkiem stężeń osoczowego żelaza. Jak wynika z tych badań, we wczesnych etapach ochrony przed bakteriemią i szokiem septycznym może mieć znaczenie chelatowanie żelaza ustrojowego przez LF, gdyż podobnie działało inne białko chelatujące żelazo – ferrytyna. Podana dożylnie 24 h przed letalną dawką bakterii chroniła zwierzęta przed śmiercią w sposób zależny od dawki [54]. Ochronne działanie LF potwierdzono również u myszy z bakteriemią E. coli [94]. Pojedyncza dożylna dawka LF 24 h przed zakażeniem znacznie zmniejszała śmiertelność, powodując czasowy spadek (o 45%) stężeń osoczowego żelaza. Jednakże podanie żelaza myszom krótko przed lub po zakażeniu tylko nieznacznie zmniejszało ochronne działanie LF, co sugeruje bardziej kompleksowy mechanizm tego działania. Sekwestracja żelaza, choć może mieć pewne znaczenie w ochronnym działaniu LF w zapaleniu, nie jest jego głównym elementem. Jak wykazały liczne późniejsze badania skuteczność LF opiera się przede wszystkim na jej działaniu regulacyjnym na różne składniki układu odpornościowego zaangażowane w rozwój zapalenia i sepsy [48].
W latach 70. ub. wieku powstała hipoteza włączająca LF w powstawanie hipoferremii i niedokrwistości w stanach zapalnych [84]. Zgodnie z jej założeniami LF uwolniona z aktywowanych neutrofilów odbiera żelazo transferrynie lub współzawodniczy z nią o wiązanie żelaza uwolnionego podczas rozpadu hemoglobiny oraz komórek bakteryjnych i odpornościowych gospodarza i przenosi je do układu siateczkowo-śródbłonkowego (RES), głównie w wątrobie, gdzie jest ono magazynowane i czasowo niedostępne dla procesów erytropoezy. Uwolnione po degradacji LF żelazo jest gromadzone w postaci ferrytyny, stanowiąc tzw. powolny szlak obrotu żelaza, w którym okres połowicznego obrotu pierwiastka wynosi około 7 dni. Ta droga obrotu żelaza ma większe znaczenie w zapaleniu, podczas gdy w stanie zdrowia istnieje równowaga pomiędzy szlakiem „szybkim” (z okresem połowicznego obrotu 24 min) a szlakiem „powolnym” [84]. W warunkach prawidłowych uwalniane żelazo wiąże się z TF, która przenosi je prawie w całości do komórek erytropoetycznych. Konkurencyjne wychwytywanie żelaza przez LF neutrofilową doprowadza zatem do czasowego niedoboru tego pierwiastka. Warto pamiętać, że LF może „odbierać” żelazo TF już w słabo zakwaszonym środowisku (pH~6,5) [84]. Stężenie osoczowego żelaza spada szybko już we wczesnym okresie rozwoju choroby, osiągając wartość nawet o połowę niższą niż podczas zdrowia, a czasem żelazo jest w ogóle niewykrywalne we krwi. Podczas zdrowienia stężenie żelaza szybko wraca do normy [89]. Podobny spadek stężeń osoczowego żelaza wykazano w testach laboratoryjnych, m.in. po podaniu zwierzętom LF [84,95], IL-1 [47], bakterii i endotoksyny (LPS) [41]. Dożylne podanie prosiętom LPS lub zawiesiny komórek E. coli znacznie obniżało (o 60%) wyjściowe stężenie osoczowego żelaza już w ciągu pierwszych odpowiednio 30 i 120 min. W tym samym czasie zanotowano znaczny (3-20-krotny) wzrost stężeń osoczowej LF [41]. Można tu zatem obserwować wczesny mechanizm ochrony przez zakażeniem i endotoksemią z udziałem endogennej LF. Redukcja stężenia żelaza podczas zapalenia może być zatem spowodowana przez LF uwalnianą z aktywowanych neutrofilów lub syntetyzowaną de novo w tkankach objętych zapaleniem. Takie samo działanie będzie miała LF egzogenna dostarczona do ustroju w celach profilaktycznych lub leczniczych.
Znacznie nowsze badania nad metabolizmem żelaza wykazały, że głównym czynnikiem odpowiedzialnym za rozwój hipoferremii podczas zapalenia jest hepcydyna – białko wytwarzane przez hepatocyty i uwalniane do krążenia [9,91]. Hepcydyna ogranicza wchłanianie żelaza z enterocytów do krążenia oraz jego uwalnianie z puli makrofagowej, obniżając osoczowe stężenia tego pierwiastka. Stężenia hepcydyny są regulowane przez status żelaza w ustroju: jego niedobór obniża, a nadmiar zwiększa wytwarzanie hepcydyny. Co ważne, infekcja i zapalenie również łączą się ze zwiększonym wytwarzaniem tego białka, prowadząc do czasowej anemii. Te zdarzenia wiążą homeostazę żelaza z odpowiedzią zapalną. W regulacji wytwarzania hepcydyny podczas zapalenia uczestniczy wiele czynników, m.in. STAT (signal transducer and activator of transcription), kompleks hemojuweliny i białka morfogenicznego kości (HJV/MBP), kompleks białka HFE z receptorem TF (HFE/TFR) i inne. Są one indukowane w odpowiedzi na cytokiny prozapalne (IL-6, TNF-α) [91]. Główna rola hepcydyny w indukcji hipoferremii w zapaleniu nie wyklucza jednak udziału w omówionym mechanizmie regulacyjnym innych elementów, w tym również laktoferryny. Potwierdzają to niedawne badania, które wykazały, że TNF-α oraz IFN-γ mogą promować hipoferremię w zapaleniu poprzez mechanizmy niezależne od indukcji hepcydyny, m.in. w wyniku stymulacji syntezy ferrytyny i zmniejszenia uwalniania tkankowych zapasów żelaza (ryc. 1) [91].

Ryc. 1. Mechanizm powstawania hipoferremii podczas zapalenia. W stanie zdrowia odpowiednie (zgodne z fizjologicznym zapotrzebowaniem ustroju) ilości żelaza są wchłaniane z jelita oraz uwalniane z zapasów tkankowych (głównie z komórek układu RES wątroby). Podczas zapalenia lub przewlekłej choroby obniżają się stężenia żelaza we krwi z jednoczesną jego akumulacją w magazynach ustrojowych i ograniczonym dostarczaniem do szpiku kostnego, czemu towarzyszy rozwój anemii (tzw. anemia podczas zapalenia lub chorób chronicznych). Czynniki wpływające na obniżenie wchłaniania żelaza i jego uwalniania z komórek magazynujących to: hepcydyna, cytokiny (IL-6, IFN-γ, TNF-α), regulujące ekspresję hepcydyny i ferrytyny oraz laktoferryna
Omówiony mechanizm hipoferremii w zapaleniu może skutecznie eliminować z patogennych i potencjalnie patogennych grup mikroorganizmów wszystkie te, które nie wykształciły zdolności współzawodnictwa o żelazo z białkami chelatującymi gospodarza (takie drobnoustroje stanowią ponad 90% wszystkich mikroorganizmów). Usunięcie żelaza z płynów ustrojowych uniemożliwia również jego udział w procesach tworzenia toksycznych rodników tlenowych, które odgrywają ważną niekorzystną rolę w procesach zapalnych. Uwzględniając powyższe fakty rola opisanego mechanizmu mikrobiostatycznego i przeciwzapalnego jest nie do przecenienia.
Podsumowanie
Żelazo jest istotnym składnikiem odżywczym dla organizmów eukariotycznych i prokariotycznych. Umożliwia prawidłowy przebieg procesów metabolicznych, zatem jego niedobór uszkadza funkcje wielu komórek, w tym m.in. procesy oddechowe, proliferację i różnicowanie komórek, funkcje odpornościowe. Żelazo jest też istotnym kofaktorem tworzenia bardzo reaktywnych cząsteczek, jakimi są RFT. W warunkach fizjologicznych odgrywają ważną rolę w ustroju, w tym uczestniczą w zabijaniu patogenów. Gdy jest ich jednak za dużo stają się toksyczne, uszkadzając liczne składniki organizmu. Nadmiar żelaza w organizmie gospodarza ułatwia też jego nabywanie przez patogenne drobnoustroje, stymulując ich wzrost. Żelazo zatem jest również istotnym elementem interakcji gospodarz-patogen. Ilości dostępnego żelaza w ustroju muszą więc podlegać bardzo precyzyjnej regulacji, tak by gospodarz korzystał z żelaza, ale uniknął toksyczności a jednocześnie ograniczył jego dostępność dla patogenów (potrzeby ustroju vs przeładowanie; tworzenie potrzebnych RFT vs toksyczność). Organizm „zainwestował” w stworzenie precyzyjnego systemu kontroli pobierania, przepływu, dostępności i reaktywności żelaza, co jest tym ważniejsze, że w zasadzie brak systemu usuwania żelaza z organizmu, a kontrola jego poziomów odbywa się głównie podczas wchłaniania. LF i inne białka chelatujące żelazo są bardzo istotnymi elementami systemu homeostazy żelaza w ustroju. LF uczestniczy nie tylko w procesach absorpcji w jelicie, transporcie i magazynowaniu żelaza w komórkach ustroju, ale też reguluje jego dostępność w reakcjach tworzenia wolnych rodników tlenowych oraz dostępność dla drobnoustrojów pasożytujących w organizmie. Te dwie ostatnie funkcje stanowią ważny element przeciwmikrobiologicznej aktywności LF. Udział LF w procesach tworzenia RFT zostanie dokładnie omówiony w III części artykułu (w przygotowaniu).
PIŚMIENNICTWO
[1] Abdallah F.B., El Hage Chahine J.M.: Transferrins: iron release from lactoferrin. J. Mol. Biol., 2000; 303: 255-266
[PubMed]
[2] Abe F., Tateyma M., Shibuya H., Azumi N., Ommura Y.: Experimental candidiasis in iron overload. Mycopathologia, 1985; 89: 59-63
[PubMed]
[3] Alexander J., Limaye A.P., Ko C.W., Bronner M.P., Kowdley K.V.: Association of hepatic iron overload with invasive fungal infection in liver transplant recipients. Liver Transpl., 2006; 12: 1799-1804
[PubMed] [Full Text PDF]
[4] Almeida R.S., Wilson D., Hube B.: Candida albicans iron acquisition within the host. FEMS Yeast Res., 2009; 9: 1000-1012
[PubMed]
[5] Altes A., Remacha A.F., Sarda P., Sancho F.J., Sureda A., Martino R., Briones J., Brunet S., Canals C., Sierra J.: Frequent severe liver iron overload after stem cell transplantation and its possible association with invasive aspergillosis. Bone Marrow Transplant., 2004; 34: 505-509
[PubMed]
[6] Andersen J.H., Osbakk S.A., Vorland L.H., Traavik T., Gutteberg T.J.: Lactoferrin and cyclic lactoferricin inhibit the entry of human cytomegalovirus into human fibroblasts. Antiviral Res., 2001; 51: 141-149
[PubMed]
[7] Anderson B.F., Baker H.M., Norris G.E., Rice D.W., Baker E.N.: Structure of lactoferrin: crystallographic structure of analysis and refinement at 2.8A resolution. J. Mol. Biol., 1989; 209; 711-734
[PubMed]
[8] Andersson Y., Lindquist S., Lagerqvist C., Hernell O.: Lactoferrin is responsible for the fungistatic effect of human milk. Early Hum. Dev., 2000; 59: 95-105
[PubMed]
[9] Andrews N.C.: Forging a field: the golden age of iron biology. Blood, 2008; 112: 219-230
[PubMed]
[10] Andrews S.C., Robinson A.K., Rodriguez-Quinones F.: Bacterial iron homeostasis. FEMS Microbiol. Rev., 2003; 27: 215-237
[PubMed]
[11] Antonini G., Catania M.R., Greco R., Longhi C., Pisciotta M.G., Seganti L., Valenti P.: Anti-invasive activity of bovine lactoferrin against Listeria monocytogenes. J. Food Prot., 1997; 60: 267-271
[12] Arnold R.R., Brewer M., Gauthier J.J.: Bactericidal activity of human lactoferrin: sensitivity of a variety of microorganisms. Infect. Immun., 1980; 28: 893-898
[PubMed] [Full Text HTML] [Full Text PDF]
[13] Arnold R.R., Russell J.E., Champion W.J., Brewer M., Gauthier J.J.: Bactericidal activity of human lactoferrin: differentiation from the stasis of iron deprivation. Infect. Immun., 1982; 35: 792-799
[PubMed] [Full Text HTML] [Full Text PDF]
[14] Artym J.: Udział laktoferryny w gospodarce żelazem w organizmie. Część I. Wpływ laktoferryny na wchłanianie, transport i magazynowanie żelaza. Postepy Hig. Med. Dosw., 2008; 62: 599-612
[PubMed] [Full Text HTML] [Full Text PDF]
[15] Barluzzi R., Saleppico S., Nocentini A., Boelaert J.R., Neglia R., Bistoni F., Blasi E.: Iron overload exacerbates experimental meningoencephalitis by Cryptococcus neoformans. J. Neuroimmunol., 2002; 132: 140-146
[PubMed]
[16] Bartosz G.: Druga twarz tlenu. Wolne rodniki w przyrodzie. Wydawnictwo Naukowe PWN, Warszawa, 2003
[17] Baynes R.D., Bezwoda W.R.: Lactoferrin and the inflammatory response. Adv. Exp. Med. Biol., 1994; 357: 133-141
[PubMed]
[18] Bhimani R.S., Vendrov Y., Furmanski P.: Influence of lactoferrin feeding and injection against systemic staphylococcal infections in mice. J. Appl. Microbiol., 1999; 86: 135-144
[PubMed]
[19] Brock J.H., Pickering M.G., McDowall M.C., Deacon A.G.: Role of antibody and enterobactin in controlling growth of Escherichia coli in human milk and acquisition of lactoferrin- and transferrin-bound iron by Escherichia coli. Infect. Immun., 1983; 40: 453-459
[PubMed] [Full Text HTML] [Full Text PDF]
[20] Brodie A.M., Ainscough E.W., Baker E.N., Baker H.M., Shongwe M.S., Smith C.A.: Synergism and substitution in the lactoferrins. Adv. Exp. Med. Biol., 1994; 357: 33-44
[PubMed]
[21] Broxmeyer H.E., deSousa M., Smithyman A., Ralph P., Hamilton J., Kurland J.I., Bognacki J.: Specificity and modulation of the action of lactoferrin, a negative regulator of myelopoiesis. Blood, 1980; 55: 324-333
[PubMed] [Full Text PDF]
[22] Bullen J.J.: The significance of iron in infection. Rev. Infect. Dis., 1981; 3: 1127-1138
[PubMed]
[23] Bullen J.J., Rogers H.J., Griffiths E.: Role of iron in bacterial infection. Curr. Top. Microbiol. Immunol., 1978; 80: 1-35
[PubMed]
[24] Byrd T.F., Horwitz M.A.: Interferon gamma-activated human monocytes downregulate transferrin receptors and inhibit the intracellular multiplication of Legionella pneumophila by limiting the availability of iron. J. Clin. Invest., 1989; 83: 1457-1465
[PubMed] [Full Text HTML] [Full Text PDF]
[25] Byrd T.F., Horwitz M.A.: Lactoferrin inhibits or promotes Legionella pneumophila intracellular multiplication in nonactivated and interferon gamma-activated human monocytes depending upon its degree of iron saturation. Iron-lactoferrin and nonphysiologic iron chelates reverse monocyte activation against Legionella pneumophila. J. Clin. Invest., 1991; 88: 1103-1112
[PubMed] [Full Text HTML] [Full Text PDF]
[26] Campagnari A.A., Shanks K.L., Dyer D.W.: Growth of Moraxella catarrhalis with human transferrin and lactoferrin: expression of iron-responsible proteins without siderophore production. Infect. Immun., 1994; 62: 4909-4914
[PubMed] [Full Text HTML] [Full Text PDF]
[27] Collins H.E.: The role of iron in infections with intracellular bacteria. Immunol. Lett., 2003; 85: 193-195
[PubMed]
[28] Collins H.L., Kaufmann S.H., Schaible U.E.: Iron chelation via deferoxamine exacerbates experimental salmonellosis via inhibition of the nicotinamide adenine dinucleotide phosphate oxidase-dependent respiratory burst. J. Immunol., 2002; 168: 3458-3463
[PubMed] [Full Text HTML] [Full Text PDF]
[29] Coughlin R.T., Tonsager S., McGroarty E.J.: Quantitation of metal cations bound to membranes and extracted lipopolysaccharide of Escherichia coli. Biochemistry, 1983; 22: 2002-2007
[PubMed]
[30] Dhaenens L., Szczebara F., Husson M.O.: Identification, characterization, and immunogenicity of the lactoferrin-binding protein from Helicobacter pylori. Infect. Immun., 1997; 65: 514-518
[PubMed] [Full Text HTML] [Full Text PDF]
[31] Doherty C.P.: Host-patogen interactions: the role of iron. J. Nutr., 2007; 137: 1341-1344
[PubMed] [Full Text HTML] [Full Text PDF]
[32] Doring G., Pfestorf M., Botzenhart K., Abdallah M.A.: Impact of proteases on iron uptake of Pseudomonas aeruginosa pyoverdin from transferrin and lactoferrin. Infect. Immun., 1988; 56: 291-293
[PubMed] [Full Text HTML] [Full Text PDF]
[33] Edde L., Hipolito R.B., Hwang F.F., Headon D.R., Shalwitz R.A., Sherman M.P.: Lactoferrin protects neonatal rats from gut-related systemic infection. Am. J. Physiol. Gastrointest. Liver Physiol., 2001; 281: 1140-1150
[PubMed] [Full Text HTML] [Full Text PDF]
[34] Ellison R.T.3rd, Giehl T.J.: Killing of Gram-negative bacteria by lactoferrin and lysozyme. J. Clin. Invest., 1991; 88: 1080-1091
[PubMed] [Full Text HTML] [Full Text PDF]
[35] Ellison R.T.3rd, Giehl T.J., LaForce F.M.: Damage of the outer membrane of enteric Gram-negative bacteria by lactoferrin and transferrin. Infect. Immun., 1988; 56: 2774-2781
[PubMed] [Full Text HTML] [Full Text PDF]
[36] Esposito J.J., Obljeski J.F.: Enterovirus type 70 virion and intracellular proteins. J. Virol., 1976; 18: 1160-1162
[PubMed] [Full Text HTML] [Full Text PDF]
[37] Evans D.M., Frazer I.H., Martin N.G.: Genetic and environmental causes of variation in basal levels of blood cells. Twin. Res., 1999; 2: 250-257
[PubMed]
[38] Freestone P.P., Lyte M., Neal C.P., Maggs A.F., Haigh R.D., Williams P.H.: The mammalian neuroendocrine hormone norepinephrine supplies iron for bacterial growth in the presence of transferrin or lactoferrin. J. Bacteriol., 2000; 182: 6091-6098
[PubMed] [Full Text HTML] [Full Text PDF]
[39] Goldoni P., Sinibaldi L., Valenti P., Orsi N.: Metal complexes of lactoferrin and their effect on the intracellular multiplication of Legionella pneumophila. Biometals, 2000; 13: 15-22
[PubMed]
[40] Groenink J., Almstahl A., Veerman E.C., Wikstrom M., Nieuw Amerongen A.V.: Salivary lactoferrin in relation to the colonisation of oral pathogens. 6th International Conference on Lactoferrin, Capri, Italy, 5-9 May 2003; Materiały zjazdowe: 69
[41] Gutteberg T.J., Rokke O., Andersen O., Jorgensen T.: Early fall of circulating iron and rapid rise of lactoferrin in septicemia and endotoxemia: an early defence mechanism. Scand. J. Infect. Dis., 1989; 21: 709-715
[PubMed]
[42] Husson M.O., Legrand D., Spik G., Leclerc H.: Iron acquisition by Helicobacter pylori: importance of human lactoferrin. Infect. Immun., 1993; 61: 2694-2697
[PubMed] [Full Text HTML] [Full Text PDF]
[43] Iglesias-Osma C., Gonzalez-Villaron L., San Miguel J.F., Caballero M.D., Vazquez L., de Castro S.: Iron metabolism and fungal infections in patients with haematological malignancies. J. Clin. Pathol., 1995; 48: 223-225
[PubMed] [Full Text HTML] [Full Text PDF]
[44] Inouye Y., Kanamori T., Yoshida T., Bu X., Shionoya M., Koike T. Kimura E.: Inhibition of human immunodeficiency virus proliferation by macrocyclic polyamines and their metal complexes. Biol. Pharm. Bull., 1994; 17: 243-250
[PubMed]
[45] Jenssen H., Hancock R.E.: Antimicrobial properties of lactoferrin. Biochemie, 2009; 91: 19-29
[PubMed]
[46] Kakhlon O., Cabantchik Z.I.: The labile iron pool: characterization, measurement, and participation in cellular processes. Free Radic. Biol. Med., 2002; 33: 1037-1046
[PubMed]
[47] Kampschmidt R.F., Mesecher M.: Interleukin-1 from P388D1: effects upon neutrophils, plasma iron, and fibrinogen in rats, mice, and rabbits. Proc. Soc. Exp. Biol. Med., 1985; 179: 197-200
[PubMed]
[48] Kruzel M.L.: Rola laktoferyny w rozwoju ostrych stanów zapalnych. Postepy Hig. Med. Dosw. 2003; 57: 377-404
[PubMed]
[49] Leclercq A., Martin L., Vergnes M.L., Ounnoughene N., Laran J.F., Giraud P., Carniel E.: Fatal Yersinia enterocolitica biotype 4 serovar O:3 sepsis after red blood cell transfusion. Tansfusion, 2005; 45: 814-818
[PubMed]
[50] Leitch E.C., Willcox M.D.: Synergic antistaphylococcal properties of lactoferrin and lysozyme. J. Med. Microbiol., 1998; 47: 837-842
[PubMed] [Full Text PDF]
[51] Levi S., Yewdall S.J., Harrison P.M., Santambrogio P., Cozzi A., Rovida E., Albertini A., Arosio P.: Evidence of H- and L-chains have co-operative roles in the iron-uptake mechanism of human ferritin. Biochem. J., 1992; 288: 591-596
[PubMed] [Full Text HTML] [Full Text PDF]
[52] Li S., Zhou H., Huang G., Liu N.: Inhibition of HBV infection by bovine lactoferrin and iron-, zinc-saturated lactoferrin. Med. Microbiol. Immunol., 2009; 198: 19-25
[PubMed]
[53] Lieu P.T., Heiskala M., Peterson P., Yang Y.: The roles of iron in health and disease. Mol. Aspects Med., 2001; 22: 1-87
[PubMed]
[54] Lipiński P., Jarząbek Z., Broniek S., Zagulski T.: Protective effect of tissue ferritins in experimental Escherichia coli infection of mice in vivo. Int. J. Exp. Pathol., 1991; 72: 623-630
[PubMed]
[55] Longhi C., Conte M.P., Seganti L., Polidoro M., Alfsen A., Valenti P.: Influence of lactoferrin on the entry process of Escherichia coli HB101 (pRI203) in HeLa cells. Med. Microbiol. Immunol., 1993; 182: 25-35
[PubMed]
[56] Lonnerdal B.: Lactoferrin receptors in intestinal brush border membranes. Adv. Exp. Med. Biol., 1994; 357: 171-175
[PubMed]
[57] Małaczewska J., Rotkiewicz Z., Siwicki A.K.: Laktoferyna – mechanizmy działania przeciwwirusowego. Medycyna Wet., 2006; 62: 1104-1107
[Full Text PDF]
[58] Marchetti M., Pisani S., Antonini G., Valenti P., Seganti L., Orsi N.: Metal complexes of bovine lactoferrin inhibit in vitro replication of herpes simplex virus type 1 and 2. Biometals, 1998; 11: 89-94
[PubMed]
[59] Marchetti M., Superti F., Ammendolia M.G., Rossi P., Valenti P., Seganti L.: Inhibition of poliovirus type 1 infection by iron-, manganese- and zinc-saturated lactoferrin. Med. Microbiol. Immunol., 1999; 187: 199-204
[PubMed]
[60] Masson P.L., Heremans J.F., Prignot J.J., Wauters G.: Immunohistochemical localization and bacteriostatic properties of an iron-binding protein from bronchial mucus. Thorax, 1966; 21: 538-543
[PubMed] [Full Text HTML] [Full Text PDF]
[61] Menozzi F.D., Gantiez C., Locht C.: Identification and purification of transferrin- and lactoferrin-binding proteins of Bordetella pertussis and Bordetella bronchiseptica. Infect. Immun., 1991; 59: 3982-3988
[PubMed] [Full Text HTML] [Full Text PDF]
[62] Mickelsen P.A., Blackman E., Sparling P.F.: Ability of Neisseria gonorrhoeae, Neisseria meningitidis, and commensal Neisseria species to obtain iron from lactoferrin. Infect. Immun., 1982; 35: 915-920
[PubMed] [Full Text HTML] [Full Text PDF]
[63] Mulayim B., Celik N.Y., Yanik F.F.: Helicobacter pylori infection detected by 14C-urea breath test is associated with iron deficiency anemia in pregnant women. J. Obstet. Gynaecol. Res., 2008; 34: 980-985
[PubMed]
[64] Nikawa H., Samaranayake L.P., Tenovuo J., Pang K.M., Hamada T.: The fungicidal effect of human lactoferrin on Candida albicans and Candida crusei. Arch. Oral. Biol., 1993; 38: 1057-1063
[PubMed]
[65] Okutomi T., Abe S., Tansho S., Wakabayashi H., Kawase K., Yamaguchi H.: Augmented inhibition of growth of Candida albicans by neutrophils in the presence of lactoferrin. FEMS Immunol. Med. Microbiol., 1997; 18: 105-112
[PubMed]
[66] Patruta S.I., Hörl W.H.: Iron and infection. Kidney Int. Suppl., 1999; 69: S125-S130
[PubMed]
[67] Perraudin J.P., Prieels J.P.: Lactoferrin binding to lysozyme-treated Micrococcus luteus. Biochim. Biophys. Acta, 1982; 718: 42-48
[PubMed]
[68] Peterson K.M., Alderete J.F.: Iron uptake and increased intracellular enzyme activity follow host lactoferrin binding by Trichomonas vaginalis receptors. J. Exp. Med., 1984; 160: 398-410
[PubMed] [Full Text HTML] [Full Text PDF]
[69] Puddu P., Borghi P., Gessani S., Valenti P., Belardelli F., Seganti L.: Antiviral effect of bovine lactoferrin saturated with metal ions on early steps of human immunodeficiency virus type 1 infection. Int. J. Biochem. Cell. Biol., 1998; 30: 1055-1062
[PubMed]
[70] Rich I.N.: The macrophage as a production site for hematopoietic regulator molecules: sensing and responding to normal and pathophysiological signals. Anticancer Res., 1988; 8: 1015-1040
[PubMed]
[71] Rossi P., Giansanti F., Boffi A., Ajello M., Valenti P., Chiancone E., Antonini G.: Ca2+ binding to bovine lactoferrin enhances protein stability and influences the release of bacterial lipopolysaccharide. Biochem. Cell Biol., 2002; 80: 41-48
[PubMed]
[72] Ryu S.Y., Jeong K.S., Kang B.N, Park S.J., Yoon W.K., Kim S.H., Kim T.H.: Modulation of transferrin synthesis, transferrin receptor expression, iNOS expression and NO production in mouse macrophages by cytokines, either alone or in combination. Anticancer Res., 2000; 20: 3331-3338
[PubMed]
[73] Sandrini S.M., Shergill R., Woodward J., Maralikuttan R., Haigh R.D., Lyte M., Freestone P.P.: Elucidation of mechanism by which catecholamine stress hormones liberate iron from innate immune defense proteins transferrin and lactoferrin. J. Bacteriol., 2010; 192: 587-594
[PubMed] [Full Text HTML] [Full Text PDF]
[74] Schaible U.E., Collins H.L., Priem F., Kaufmann H.E.: Correction of the iron overload defect in β-2-microglobulin knockout mice by lactoferrin abolishes their increased susceptibility to tuberculosis. J. Exp. Med., 2002; 196: 1507-1513
[PubMed] [Full Text HTML] [Full Text PDF]
[75] Schaible U.E., Kaufmann S.H.: Iron and microbial infection. Nat. Rev. Microbiol., 2004; 2: 946-953
[PubMed]
[76] Schryvers A.B.: Identification of the transferrin- and lactoferrin-binding proteins in Haemophilus influenzae. J. Med. Microbiol., 1989; 29: 121-130
[PubMed] [Full Text PDF]
[77] Singh P.K., Parsek M.R., Greenberg E.P., Welsh M.J.: A component of innate immunity prevents bacterial biofilm development. Nature, 2002; 417: 552-555
[PubMed]
[78] Soukka T., Tenovuo J., Lenander-Lumikari M.: Fungicidal effect of human lactoferrin against Candida albicans. FEMS Microbiol. Lett., 1992; 90: 223-228
[PubMed]
[79] Spik G., Cheron A., Montreuil J., Dolby J.M.: Bacteriostasis of a milk-sensitive strain of Escherichia coli by immunoglobulins and iron-binding proteins in association. Immunology, 1978; 35: 663-671
[PubMed] [Full Text HTML] [Full Text PDF]
[80] Stintzi A., Raymond K.N.: Amonabactin-mediated iron acquisition from transferrin and lactoferrin by Aeromonas hydrophila: direct measurement of individual microscopic rate constants. J. Biol. Inorg. Chem., 2000; 5: 57-66
[PubMed]
[81] Superti F., Ammendolia M.G., Valenti P., Seganti L.: Antirotaviral activity of milk proteins: lactoferrin prevents rotavirus infection in the enterocyte-like cell line HT-29. Med. Microbiol. Immunol., 1997; 186; 83-91
[PubMed]
[82] Sutak R., Lesuisse E., Tachezy J., Richardson D.R.: Crusade for iron: iron uptake in unicellular eukaryotes and its significance for virulence. Trends Microbiol., 2008; 16: 261-268
[PubMed]
[83] Torti F.M., Torti S.V.: Regulation of ferritin genes and protein. Blood, 2002; 99: 3505-3516
[PubMed] [Full Text HTML] [Full Text PDF]
[84] van Snick J.L., Masson P.L., Heremans J.F.: The involvement of lactoferrin in the hyposideremia of acute inflammation. J. Exp. Med., 1974; 140: 1068-1084
[PubMed] [Full Text HTML] [Full Text PDF]
[85] Vogel L., Geluk F., Jansen H., Dankert J., van Alphen L.: Human lactoferrin receptor activity in non-encapsulated Haemophilus influenzae. FEMS Microbiol. Lett., 1997; 156: 165-170
[PubMed]
[86] Wanachiwanawin W.: Infections in E-beta thalassemia J. Pediatr. Hematol. Oncol., 2000; 22: 581-587
[PubMed]
[87] Wang X., Hirmo S., Willen R., Wadstrom T.: Inhibition of Helicobacter pylori infection by bovine milk glycoconjugates in BALB/cA mouse. J. Med. Microbiol., 2001; 50: 430-435
[PubMed] [Full Text PDF]
[88] Weinberg E.D.: Human lactoferrin: a novel therapeutic with broad spectrum potential. J. Pharm. Pharmacol., 2001; 53: 1303-1310
[PubMed]
[89] Weinberg E.D.: Iron and infection. Microbiol. Rev., 1978; 42: 45-66
[PubMed] [Full Text HTML] [Full Text PDF]
[90] Weinberg E.D.: Suppression of bacterial biofilm formation by iron limitation. Med. Hypotheses, 2004; 63: 863-865
[PubMed]
[91] Wilson M.E., Vorhies R.W., Andersen K.A., Britigan B.E.: Acquisition of iron from transferrin and lactoferrin by the protozoan Leishmania chagasi. Infect. Immun., 1994; 62: 3262-3269
[PubMed] [Full Text HTML] [Full Text PDF]
[92] Wolach B., Coates T.D., Hugli T.E., Baehner R.L., Boxer L.A.: Plasma lactoferrin reflects granulocyte activation via complement in burn patients. J. Lab. Clin. Med., 1984; 103: 284-293
[PubMed]
[93] Zagulski T., Lipiński P., Zagulska A., Broniek S., Jarząbek Z.: Lactoferrin can protect mice against lethal dose of Escherichia coli in experimental infection in vivo. Br. J. Exp. Path., 1989; 70: 697-704
[PubMed]
[94] Zagulski T., Zagulska A., Jedra M., Jarząbek Z.: Rabbit plasma iron level after in vivo administration of lactoferrin. Prace i Mater. Zoot., 1985; 36: 95-105
[95] Zarember K.A., Cruz A.R., Huang C.Y. Gallin J.I.: Antifungal activities of natural and synthetic iron chelators alone and in combination with azole and polyene antibiotics against Aspergillus fumigatus. Antimicrob. Agents Chemother., 2009; 53: 2654-2656
[PubMed] [Full Text HTML] [Full Text PDF]
[96] Zarember K.A., Sugui J.A., Chang Y.C., Kwon-Chung K. J., Gallin J.I.: Human polymorphonuclear leukocytes inhibit Aspergillus fumigatus conidial growth by lactoferrin-mediated iron depletion. J. Immunol., 2007; 178: 6367-6373
[PubMed] [Full Text HTML] [Full Text PDF]
[97] Zhou H., Li S., Huang G., Liu N.: Study of inhibition effect of zinc-, iron-, and manganese-saturated bovine lactoferrin on hepatitis B virus DNA in vitro. Wei Sheng Yan Jiu, 2008; 37: 586-589
[PubMed]
Autorka deklaruje brak potencjalnych konfliktów interesów.