
Właściwości biologiczne białek powierzchniowych bakterii z rodzaju Lactobacillus
Barbara Buda 1 , Ewa Dylus 1 , Sabina Górska-Frączek 1 , Ewa Brzozowska 1 , Andrzej Gamian 1Streszczenie
Bakterie z rodzaju Lactobacillus to Gram-dodatnie bakterie, wśród których znajduje się wiele szczepów zakwalifikowanych do tzw. mikroflory probiotycznej. Probiotyki, jak wynika z definicji, to żywe mikroorganizmy, które korzystnie wpływają na zdrowie gospodarza. W pracy zebrano informacje na temat morfologii i fizjologii bakterii z rodzaju Lactobacillus. Przedstawiono budowę ściany komórkowej bakterii Gram-dodatnich. Opisano występującą u nich warstwę powierzchniową S (S-layer) zbudowaną z białek (SLP) o niewielkiej masie cząsteczkowej. Opisano wpływ czynników stresowych, takich jak wysoka temperatura, pH, enzymy soku żołądkowego i trzustkowego na ekspresję białek warstwy S, a także przedstawiono zdolność białek powierzchniowych do wiązania jonów metali ciężkich. Scharakteryzowano białka powierzchniowe biorące udział w regulacji wzrostu komórek nabłonka jelitowego. Omówiono właściwości białek powierzchniowych, wśród których można wymienić: zdolności adhezyjne do komórek nabłonkowych, zahamowanie inwazji patogenów z rodzajów m.in.: Shigella, Salmonella, Escherichia. Opisano zdolność obniżania przez białka powierzchniowe aktywności toksyn wydzielanych przez Clostridium.
Słowa kluczowe:Lactobacillus • probiotyki • ściana komórkowa • warstwa powierzchniowa S • białka powierzchniowe
Summary
Lactobacillus, a genus of Gram-positive bacteria, includes many strains of probiotic microflora. Probiotics, by definition, are living microorganisms that exert beneficial effects on the host organism. The morphology and physiology of the Lactobacillus bacterial genus are described. The structure of the cell wall of Gram-positive bacteria is discussed. The surface S-layer of Lactobacillus composed of proteins (SLP) with low molecular mass is presented. Cell surface proteins participating in the regulation of growth and survival of the intestinal epithelium cells are characterized. The influence of stress factors such as increased temperature, pH, and enzymes of gastric and pancreatic juice on SLP expression is described. The ability of binding of heavy metal ions by S-layer proteins is discussed. The characteristics of these structures, including the ability to adhere to epithelial cells, and the inhibition of invasion of pathogenic microflora of type Shigella, Salmonella, Escherichia coli and Clostridium and their toxins, are presented.
Key words:Lactobacillus • probiotics • cell wall • surface S-layer • surface proteins
Charakterystyka bakterii z rodzaju Lactobacillus
Bakterie z rodzaju Lactobacillus to nieprzetrwalnikujące mikroorganizmy, które wykazują znaczne zróżnicowanie pod względem morfologii. Komórki bakteryjne różnych gatunków Lactobacillus mają postać od długich, prostych i wąskich, poprzez wygięte, aż do krótkich i grubych laseczek, w zależności od wieku hodowli jak i składu medium hodowlanego (ryc. 1). Na podłożu stałym rosną w postaci małych (2-5 mm), przeważnie gładkich, śliskich lub matowych kolonii o barwie kremowej bądź bezbarwnej. Pod względem metabolizmu węglowodanowego szczepy z rodzaju Lactobacillus mogą należeć do jednej z trzech grup: bezwzględnie homofermentatywne, względnie heterofermentatywne oraz bezwzględnie heterofermentatywne. Głównym produktem szlaków metabolicznych jest kwas mlekowy. Ponadto dodatkowymi produktami mogą być kwas octowy, mrówkowy, etanol, dwutlenek węgla. Są to bakterie mikroaerofilne, nie trawią kazeiny, są katalazoujemne, nie wytwarzają siarkowodoru, a większość szczepów wytwarza niewielkie ilości rozpuszczalnego azotu. Optymalna temperatura wzrostu bakterii Lactobacillus mieści się w granicach 30-40 °C, natomiast pH zwykle 5,5-6,2, a najlepszym podłożem wzrostowym jest MRS agar (de Man, Rogosa and Sharpe). Analizując budowę ich ściany komórkowej, można je zakwalifikować do grupy bakterii Gram-dodatnich ze względu na obecność wielu warstw peptydoglikanu oraz braku błony zewnętrznej. Niektóre szczepy są zdolne do wytwarzania pozakomórkowych homo- i heteropolisacharydów (PS) [3].
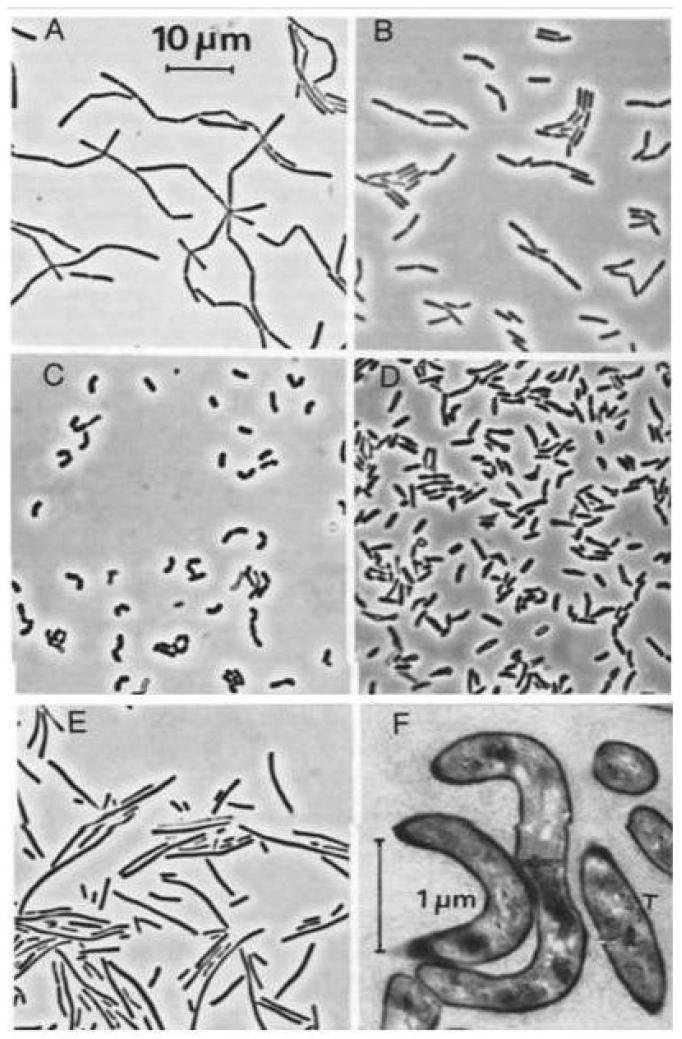
Ryc. 1. Zdjęcia spod mikroskopu kontrastowo-fazowego (A-E) oraz spod mikroskopu elektronowego (F) ukazujące różnice w morfologii komórek bakterii Lactobacillus: A – Lactobacillus gasseri, B – Lactobacillus agilis, C – Lactobacillus curvatus, D – Lactobacillus minor, E – Lactobacillus fermentum, F – inwolucyjna postać bakterii Lactobacillus wyizolowanych z ziaren kefirowych [3]
Gatunki bakterii z rodzaju Lactobacillus bytuje nie tylko w żywności pochodzenia roślinnego, czy też sfermentowanych produktach mlecznych, ale również w przewodzie pokarmowym ludzi oraz zwierząt [22].
Bakterie z rodzaju Lactobacillus jako probiotyki
Wśród bakterii kwasu mlekowego wiele szczepów z rodzaju Lactobacillus zakwalifikowano do grona probiotyków jako mikroflorę, która spożywana w odpowiednich ilościach wpływa korzystnie na zdrowie człowieka [31]. Zalecane dzienne spożycie dawki probiotyków (niezależne od sposobu podawania i rodzaju preparatu), wynosi 109-1011 komórek [35]. Do probiotycznych szczepów bakterii z rodzaju Lactobacillus można zaliczyć m.in.: L. rhamnosus, L. casei, L. bulgaricus, L. acidophilus. Ochrona przed zakażeniami poprzez modulację układu odpornościowego, obniżanie reakcji alergicznych oraz zdolność do zasiedlania nabłonka jelitowego, skłoniły do rozpoczęcia badań nad wykorzystaniem wybranych gatunków Lactobacillus jako składników preparatów terapeutycznych lub starterów w procesach produkcji żywności probiotycznej [2,19,32,51]. Opisano korzystny wpływ laseczek Lactobacillus w leczeniu zakażeń rotawirusowych, a także obniżaniu zdolności adhezyjnej i inwazyjnej mikroflory patogennej, takiej jak bakterie z rodzaju E. coli, Salmonella. Zaobserwowano, że szczepy probiotyczne obniżają ryzyko pojawienia się biegunki po terapii antybiotykowej, wpływają na regulację wzrostu, proliferację oraz spowalnianie apoptozy komórkowej w zapaleniach jelit [2,51].
W ostatnich latach położono nacisk na prowadzenie badań mających na celu ocenę wymiernych korzyści z wykorzystania bakterii probiotycznych, zwłaszcza z rodzajów Lactobacillus oraz Bifidobacterium. Wyniki najnowszych badań wskazują, że leczenie dużymi dawkami L. rhamnosus GG lub L. reuteri SD2222 znacznie skraca czas trwania ostrej biegunki u dzieci, a podawanie żywych bakterii L. acidophilus zmniejsza ryzyko rozwoju martwiczego nowotworu jelit u noworodków. Ponadto wykazano, że zastosowanie szczepów probiotycznych, takich jak L. acidophilus oraz L. johnsonii podczas terapii antybiotykowej łagodzi objawy zakażenia wywołanego przez Helicobater pylori [20,25]. Wyniki badań prowadzonych na modelach zwierzęcych wskazują, że probiotyki biorą udział w modulacji układu odpornościowego. Przykładem może być zmniejszenie częstości występowania alergii (wyprysków atopowych) u noworodków po podaniu bakterii L. rhamnosus GG [23]. Ponadto zaobserwowano łagodzenie objawów nietolerancji laktozy przy podawaniu produktu zawierającego osiem szczepów probiotycznych, w tym czterech z rodzaju Lactobacillus [35,52]. Odnotowano również, że spożywanie jogurtu zawierającego kultury bakterii L. acidophilus 145 zmniejsza w krwi stężenie cholesterolu LDL (low-density lipoproteins, czyli kompleksy białkowo-tłuszczowe o małej gęstości) [27,35]. Podobne wyniki dotyczące redukcji stężenia cholesterolu we krwi otrzymano w badaniach z wykorzystaniem modelu myszy z hipercholesterolemią. Siedmiodniowe podawanie myszom bakterii z rodzaju L. reuteri CRL 1098 (dawek o stężeniu 109 komórek/dzień), spowodowało obniżenie ogólnej zawartości cholesterolu o 38% [25]. Pojawiły się wyniki badań nad wykorzystaniem mikroflory probiotycznej w zapobieganiu infekcjom układu moczowo-płciowego, w leczeniu nowotworów pęcherza moczowego oraz hamowaniu tworzenia się kamieni nerkowych. Wykazano, że szczepy L. rhamnosus GR-1, L. fermentum RC-14 oraz L. crispatus CTV-05 działają korzystnie w zakażeniach moczowo-płciowych, a także w stanach zapalnych pochwy [20]. W jednej z prac ukazały się wyniki badań, w których obserwowano korzystny wpływ szczepu L. casei Shirota polegający na hamowaniu indukowanego chemicznie nowotworu pęcherza u myszy i szczurów [20]. Pojawiły się również doniesienia, że doustne podawanie bakterii L. casei zapobiega nawrotom powierzchniowego nowotworu pęcherza [35]. Większość z wymienionych badań wymaga potwierdzenia na większej grupie pacjentów.
Bardzo ważnym zagadnieniem jest poznanie mechanizmów działania bakterii probiotycznych. W tym celu prowadzone badania skupiają się głównie na poznaniu funkcji poszczególnych składników komórkowych bakterii Lactobacillus, a przede wszystkim ich udziału w procesie adhezji do komórek nabłonka jelitowego oraz właściwości terapeutycznych [32,51]. Niektóre czynniki, takie jak hydrofobowość powierzchni komórki, czy obecność kwasów tłuszczowych i białek, mogą mieć wpływ na adhezję komórek do tkanek gospodarza [2,26]. Pojawiło się wiele publikacji na temat zdolności adhezyjnych różnych gatunków bakterii z rodzaju Lactobacillus. W jednej z nich wykazano, że możliwe jest przetrwanie i kolonizacja nabłonka przewodu pokarmowego i dróg moczowo-płciowych przez doustne podawanie dwóch szczepów: L. rhamnosus GR-1 oraz L. fermentum RC-14. Pozwoliło to na rozpoczęcie badań nad stworzeniem nowego suplementu diety, który zapewni właściwą mikroflorę kobiecych dróg moczowo-płciowych oraz ochronę przed infekcjami [12]. Zdolności adhezyjne szczepów probiotycznych znalazły również swoje wykorzystanie w hamowaniu zakażeń przez patogenne komórki Escherichia coli. Udowodniono, że w wielu przypadkach jest to związane z wytwarzaniem bakteriocyn lub konkurencyjnym przyleganiem mikroflory probiotycznej do komórek nabłonka, przez co zablokowane są miejsca potencjalnego ataku przez komórki patogenu. W doświadczeniach in vitro wykazano, że gatunki bakterii, takich jak L. johnsonii La1 czy L. acidophilus LB hamują przyleganie patogenu do eukariotycznych linii komórkowych Caco-2 (linia pochodząca od gruczolakoraka 72-letniego mężczyzny służąca jako model do badań m.in. adhezji bakterii jelitowych oraz studiów nad inwazją patogenów bakteryjnych), poprzez konkurencyjną adhezję do nabłonka [16,31].
Niedawne doniesienia na temat budowy ściany komórkowej bakterii Gram-dodatnich, pozwalają na lepsze zrozumienie mechanizmów działania oraz właściwości mikroflory probiotycznej, w tym bakterii z rodzaju Lactobacillus. Szczegółowa analiza powierzchni komórki umożliwiła scharakteryzowanie struktur powierzchniowych oraz przypisanie im odpowiednich funkcji biologicznych.
Budowa ściany komórkowej bakterii Gram-dodatnich
Schematyczny przekrój przez ścianę komórkową bakterii z rodzaju Lactobacillus przedstawiono na ryc. 2.

Ryc. 2. Przekrój przez ścianę i błonę komórkową bakterii rodzaju Lactobacillus. LTA – kwasy lipotejchojowe, WTA – kwasy tejchowe (lipo-, wall-teichoic acids), EPS – egzopolisacharydy (exopolysaccharides), SDP – białka sortazozależne (sortase-dependent proteins) [30]
Ściana komórkowa bakterii Gram-dodatnich składa się głównie z peptydoglikanu, kwasów (lipo-) tejchojowych, polisacharydów oraz białek. Wewnętrzna warstwa ściany zawiera sieć peptydoglikanu – mureiny, która zbudowana jest z łańcuchów polisacharydowych złożonych z podjednostek: kwasu muraminowego oraz N-acetyloglukozaminy, połączonych wiązaniem β-1,4-glikozydowym. Cała konstrukcja utrzymywana i stabilizowana jest przez krótkie peptydy. W związku z dużą gęstością usieciowania oraz małą zdolnością zmiany konformacji, mureina jest dość sztywna i wytrzymała na rozciąganie. Kwasy tejchojowe tworzą różnego typu struktury, wśród których najczęściej występującymi są fosforany polioli (glicerolu lub rybitolu) albo polimery fosforanów glikozydowanych polioli zazwyczaj podstawionymi resztami glukozy i/lub D-alaniny, które są kowalencyjnie połączone z mureiną ściany komórkowej przez jednostkę glikozydową [21]. Kwasy lipotejchojowe różnią się od tejchojowych tym, że zakotwiczone są w membranie cytoplazmatycznej poprzez fragment lipidowy. Obydwa rodzaje kwasów przejawiają silny polielektrolityczny charakter przez obecność silnie kwasowych grup fosforanowych [41]. Oprócz kwasów tejchojowych i lipotejchojowych występują również kwasy tejchuronowe, pozbawione reszt fosforanowych i składające się z monomerów cukrowych połączonych wiązaniami glikozydowymi. Ich ujemny ładunek jest warunkowany obecnością grup karboksylowych kwasu urunowego lub glukuronowego [21].
Egzopolisacharydy (EPS) związane ze ścianą komórkową bakterii Lactobacillus mają charakter kwaśny lub neutralny. Wyróżniamy polisacharydy połączone ze ścianą (w sposób kowalencyjny lub niekowalencyjny), a także polisacharydy wydzielane do środowiska zewnętrznego [18,21]. Ze względu na ich dużą ilość na zewnętrznej powierzchni komórki, decydują one w znacznym stopniu o właściwościach komórki. Duża ilość wytwarzanych polisacharydów pozwala także na ich praktyczne wykorzystanie. Na przykład w przemyśle spożywczym mogą służyć jako środki stabilizujące, emulgujące, żelujące lub wiążące wodę [41,50].
Na powierzchni ściany komórkowej bakterii Gram-dodatnich znajduje się wiele rodzajów białek. Jednymi z nich są białka swoiście rozpoznawane przez sortazę. Zawierają one motyw LPXTG (X oznacza dowolny aminokwas), który jest umiejscowiony na C-końcu cząsteczki. Sortaza (StrA) rozcina wiązanie peptydowe między resztami treoniny i glicyny w motywie LPXTG, a następnie przyłącza białko w sposób kowalencyjny do grup aminowych peptydoglikanu w ścianie komórkowej [43]. Opisano dziesięć białek zależnych od sortazy (SDP), które wyizolowano ze szczepu L. salivarius UCC 118. Wśród nich znalazły się homologi: trzech białek wiążących śluz, białka wiążącego kolagen, białka powierzchniowego enterokoków, białka wiążącego aglutyninę śliny, homolog proteinazy, homolog białka wiążącego się z nabłonkiem jelitowym oraz dwa białka hipotetyczne [47].
Omawiając białka ściany komórkowej bakterii Gram-dodatnich, należy również zwrócić uwagę na białka wydzielane na zewnątrz komórki. W tej grupie można wyróżnić bakteriocyny lub białka działające bójczo na pokrewne organizmy. Są to białka niewielkich rozmiarów rozpoznające receptory powierzchniowe atakowanej komórki. Mechanizm ich działania jest dość zróżnicowany i może obejmować wnikanie do komórki przez wytworzone pory, niszczenie komórkowego materiału genetycznego, czy też hamowanie syntezy peptydoglikanu [45]. Wyróżnia się cztery główne grupy bakteriocyn: I – lantybiotyki (A i B), II – małe, termostabilne bakteriocyny (a wśród nich podklasy, takie jak: pediocynopodobne, sec-zależne (wydalane za pomocą białek sec), dwupeptydowe), III – duże, bakteriocyny termolabilne oraz IV – bakteriocyny, które do pełnej aktywności wymagają obecności w swojej strukturze komponentu lipidowego lub węglowodanowego. Przykładem bakteriocyny wysokocząsteczkowej może być helwetycyna J wytwarzana przez L. helvetius 481 oraz kaseicyna 80 wytwarzana przez L. casei B80 [5,44]. Inne szczepy wytwarzające bakteriocyny, to np. L. sakei wytwarzająca bakteriocyny ST22Ch, ST153Ch oraz ST154Ch, a także szczepy L. plantarum, wytwarzające wiele typów plantarycyny [44]. Ostatnie doniesienia wskazują, że bakteriocyny nie są jedynymi białkami wydzielanymi przez bakterie Lactobacillus. Szczep L. rhamnosus GG wydziela kilka rodzajów białek, a wśród nich: inhibitor elastazy, hydrolazę bakteryjną ściany komórkowej oraz homologi białek powierzchniowych. Szczep L. casei ATCC 334 wytwarza białka p40 i p75, które prawdopodobnie biorą udział w regulacji proliferacji nabłonka oraz hamowaniu apoptozy komórkowej podczas infekcji pokarmowych [40].
Innym rodzajem białek występujących na powierzchni komórki są adhezyny bakteryjne. W ciągu ostatnich lat udało się zidentyfikować wiele białek adhezyjnych na powierzchni bakterii z rodzaju Lactobacillus. Wśród nich znajduje się dehydrogenaza aldehydu 3-fosfoglicerynowego (GAPDH). Jest to enzym cytoplazmatyczny, który uczestniczy w fosforylacji oksydacyjnej aldehydu 3-fosfoglicerynowego. Z ostatnio przeprowadzonych badań wynika, że GAPDH występuje na powierzchni komórek różnych gatunków bakterii z rodzaju Lactobacillus, m.in.: L. plantarum LA 318, 423 i 299v oraz L. crispatus ST1. Jest to niezwykła i niewytłumaczona jak dotąd kwestia, gdyż enzym ten nie ma ani N-końcowej sekwencji sygnałowej, ani żadnych motywów pozwalających na wydzielenie białka z komórki i zakotwiczenie go na powierzchni ściany [28,39]. Wykazano, że GAPDH bierze udział w adhezji do błony śluzowej jelita [28] oraz ma zdolność rozpoznawania antygenów grupy krwi A i B [29]. Innym przykładem białka adhezyjnego jest MAPP (mucin adhesion-promoting protein). Białko o wielkości 29 kDa zlokalizowane na powierzchni komórek bakterii z rodzaju L. fermentum 104R, odpowiadające za adhezję do śluzówki jelita (prosiąt i świń). Prawdopodobnie MAPP jest oligomerycznym białkiem, które zawiera dwa lub więcej łańcuchów polipeptydowych czy też podjednostek białkowych, które w sposób niekowalencyjny wiążą się z powierzchnią bakterii [37]. Następnym znanym białkiem adhezyjnym jest czynnik elongacji Tu, wyizolowany z powierzchni L. johnsonii NCC533. Przypuszcza się, że białko Tu bierze udział w adherencji komórek L. johnsonii do nabłonka jelitowego oraz białek błony śluzowej, a także indukuje odpowiedź immunologiczną w stanach zapalnych jelita [17]. Wśród adhezyn białkowych można wymienić jeszcze α-enolazę (Eno A1) o masie cząsteczkowej 48 kDa, którą zlokalizowano na powierzchni komórek L. plantarum WCFS1. Enolaza do tej pory znana była jako enzym szlaku Embdena-Meyerhofa-Parnasa, w którym katalizuje przemianę 2-fosfoglicerynianu w fosfoenolopirogronian. Wyeksponowana na powierzchni komórek licznych szczepów bakterii Gram-dodatnich, w tym z rodzaju Lactobacillus, enolaza wykazuje duże powinowactwo do plazminogenu (białka wytwarzanego przez wątrobę i wydzielanego do osocza jako nieaktywny prekursor proteazy serynowej) [42] oraz ma zdolność wiązania fibronektyny (glikoproteiny biorącej udział w procesach naprawczych zniszczonych tkanek) [8].
Na powierzchni komórki obecne są również białka o dużej masie cząsteczkowej. Jednym z nich jest białko Mub o masie 352 kDa. Przypuszcza się, że Mub ma N-końcowy peptyd sygnałowy, dzięki któremu białko jest kierowane na szlak sekrecyjny. Na jego C-końcu znajduje się domniemany motyw kotwiczący w ścianie (LPQTG) oraz inne motywy łączące z błoną komórkową. Białko Mub zawiera dwa typy powtarzających się motywów aminokwasowych: sześć kopii Mub1 oraz osiem powtórzeń Mub2. W jednym z badań, za pomocą swoistych przeciwciał oraz mikroskopii fluorescencyjnej, białko Mub zostało zlokalizowane na powierzchni L. reuterii 1063. W pracy zasugerowano, że bierze ono udział w adhezji komórek do śluzówki jelita [38].
Najnowsze badania potwierdzają również obecność białek piliny u probiotycznego gatunku bakterii L. rhamnosus GG, które biorą udział w oddziaływaniach z komórką gospodarza. Obecność białek piliny została potwierdzona metodami immunoblotingu oraz mikroskopii elektronowej [46]. Białka piliny są najlepiej scharakteryzowane u bakterii Gram-ujemnych. Dopiero w ostatnich latach udało się przeprowadzić dokładną analizę białek piliny u bakterii Gram-dodatnich. Pili u tej grupy mikroorganizmów zbudowane są z kowalencyjnie ze sobą związanych polimerów (powtarzających się domen immunoglobulinopodobnych), do których podjednostki pilinowe przyłączone są za pomocą wiązań izopeptydowych [24].
Najbardziej zewnętrzną warstwą u większości szczepów bakterii Lactobacillus jest krystaliczna, dwuwymiarowa warstwa S (S-layer) (ryc.2). Struktura ta zbudowana jest głównie z białek, wśród których część stanowią glikoproteiny. Masy cząsteczkowe białek warstwy S mieszczą się w granicach 25-71 kDa, podczas gdy u innych rodzajów bakterii i Archea mogą osiągać masą do 200 kDa [9]. Białka warstwy powierzchniowej stanowią 10-15% ogólnej zawartości białek w komórce [9,26,41]. Cechą charakterystyczną białek warstwy powierzchniowej Lactobacillus jest ich wysoki punkt izoelektryczny (pI), który wynosi 9,35-10,40 i nadaje im charakter silnie zasadowy, podczas gdy u innych mikroorganizmów mają one charakter słabo kwaśny [9,32,41]. Potwierdzono istnienie dwóch genów kodujących białka warstwy S: SlpA oraz SlaB [9]. Obecność warstwy S potwierdzono u szczepów: L. helvetius, L. brevis, L. acidophilus, L. crispatus, L. amylovorus, L. gallinarum, L. gasseri, L. johnsonii [9,26]. Niedawno, w celu określenia struktury drugorzędowej białek warstwy powierzchniowej wykorzystano metody spektroskopii FTIR. Zaletą tej metody jest to, że można poddać analizie białka w różnym stanie skupienia, np. w postaci krystalicznej lub rozpuszczonej. Ponadto mogą być one umieszczone na różnych powierzchniach, m.in.: na polimerach, metalach czy podłożach bioceramicznych [34].
Według zebranych informacji, białka powierzchniowe mają dwa strukturalne i funkcjonalne regiony. Jeden z nich odpowiada za przymocowanie białka do ściany komórkowej, drugi natomiast za utrzymanie struktury białka na powierzchni. U wielu bakterii Gram-dodatnich białka powierzchniowe zawierają na N-końcu motyw SLH, odpowiedzialny za kotwiczenie podjednostek białkowych w ścianie komórkowej. Motyw SLH nie znajduje się wyłącznie u bakterii Gram-dodatnich, można go znaleźć również u wielu przedstawicieli bakterii Gram-ujemnych, Archea, u organizmów eukariotycznych, a nawet wirusów [21]. W przypadku białek powierzchniowych niemających motywu SLH, przypuszcza się, że wiązanie białka ze ścianą komórkową zachodzi między resztami zasadowych aminokwasów białka a ujemnie naładowanymi węglowodanami ściany komórkowej [21].
Ze szczepów L. johnsonii oraz L. gasserii wyizolowano białka, które stanowią czynniki wspomagające agregację (APF). Ich wielkość wahała się między 30-40 kDa [48]. Z kolei u L. reuteri JCM1081 scharakteryzowano białko powierzchniowe o masie cząsteczkowej 29 kDa. Białko to miało zdolność wiązania do składników śluzu jelitowego [50].
Zdolność wiązania jonów metali przez białka warstwy S
Żywność oraz woda są głównymi źródłami potencjalnego zakażenia metalami, takimi, które są toksyczne dla człowieka. Ich połączenia nieulegające rozkładowi mają tendencję do gromadzenia się w organizmie, co może powodować poważne skutki zdrowotne. Jedną z metod usuwania metali ze środowiska jest bioremediacja, czyli wykorzystanie czynników biologicznych do usuwania lub neutralizacji zanieczyszczeń. Wykorzystanie inaktywowanych mikroorganizmów jako biosorbentów do usuwania ciężkich metali z wody wydaje się metodą efektywną oraz ekonomicznie opłacalną. Wykorzystuje się różne gatunki bakterii kwasu mlekowego do usuwania kadmu oraz ołowiu. W celu zrozumienia mechanizmu biosorpcji oraz przypisanie jej odpowiedniej strukturze komórkowej, rozpoczęto badania nad oddziaływaniami między strukturami powierzchniowymi komórki z metalami. Wśród białek nadających zdolności wiązania metali, białka powierzchniowe jako jedyne mają wiele miejsc swoistego ich wiązania, dzięki czemu mogą absorbować kilka metali jednocześnie. W jednej publikacji opisano za pomocą spektroskopii FTIR analizę oddziaływania między białkami powierzchniowymi wyizolowanymi z dwóch szczepów: agregującego L. kefir CIDCA 8348 i nieagregującego L. kefir JCM 5818 z jonami kadmu, cynku, ołowiu i niklu. Przeprowadzono również analizę wpływu powyższych interakcji na zmiany struktury drugorzędowej białek warstwy S. Wykonane analizy pozwoliły na stwierdzenie, że metale wiążą się do grup karboksylowych reszt kwasu asparaginowego oraz glutaminowego wchodzących w skład białek powierzchniowych. Badania wykazały również, że szczep agregujący miał mniejszą zdolność wiązania metali, niż szczep nieagregujący, u którego liczba miejsc wiązania mniejszych jonów (niklu i cynku) była większa. Ponadto zauważono, że oddziaływanie bakterii z metalami wywołuje zmiany w strukturze drugorzędowej białek, a zwłaszcza zwiększenie liczby struktur β-harmonijki i zmniejszenie α-helis, zwłaszcza przy oddziaływaniu z dużymi jonami metali. Zmiany te prowadzą do zwiększenia sztywności, siły oraz wydajności rozpraszania energii podczas odkształcania komórki [13,36].
Wpływ warunków stresowych na ekspresję białek powierzchniowych
Oprócz funkcji adhezyjnych, białka powierzchniowe bakterii z rodzaju Lactobacillus pełnią także funkcję stabilizującą kształt komórki oraz chronią przed wrogimi dla komórki mikroorganizmami, a także czynnikami środowiskowymi [1,11,26,53]. Ochronną funkcję białek powierzchniowych wykazano poprzez wpływ zmian w ich strukturze drugorzędowej na właściwości jakie wykazują w stanie natywnym. W jednej z prac opisano badania dotyczące wpływu podwyższonej temperatury na strukturę białek powierzchniowych. Analizę przeprowadzono za pomocą metod FTIR na białkach wyizolowanych z wybranych szczepów L. kefir oraz L. brevis. Wykazano, że obecność struktury β-harmonijki, zwiększa zdolność agregowania, natomiast obecność α-helisy nie ma szczególnego znaczenia w mechanizmie agregacji u szczepów bakterii z rodzaju Lactobacillus. Ponadto większa zawartość struktury β-harmonijki i jednocześnie niewielka α-helisy, wpływała na wzrost odporności na wysoką temperaturę [34]. Wpływ warunków stresowych, takich jak wysoka temperatura, niskie pH, enzymy trawienne na syntezę białek powierzchniowych został opisany w 2012 roku przez Khaleghiego oraz Kermanshahiego [26]. Jest to bardzo istotna kwestia, gdyż szczepy Lactobacillus stosowane jako probiotyki muszą być zdolne do przeżycia i wzrostu w trudnych warunkach środowiskowych w przewodzie pokarmowym (zmiany kwasowości), czy też oporne na wszelkie czynniki zewnętrzne podczas przetwarzania w cyklu technologicznym (zmiany temperatury, osmotyczne czy stres oksydacyjny). Sok żołądkowy zawiera kwas solny, który tworzy ekstremalnie niskie pH. Na czczo w żołądku pH wynosi 1,5, a podczas spożywania posiłków może wzrosnąć do 3,0-5,0. W jelicie pH jest wyższe niż w żołądku, ale w dalszym ciągu mikroflora probiotyczna musi być niewrażliwa na soki trzustkowe oraz żółć. Ponadto innymi czynnikami mającymi wpływ na przeżycie i wzrost mikroflory są: stale regenerujący się nabłonek, przeciwciała, ruchy perystaltyczne jelit związane z przepływem treści oraz lepka warstwa śluzu [26]. Udowodniono, że niektóre czynniki, takie jak: kwasy lipotejchojowe, hydrofobowość powierzchni, zdolność do agregacji czy też obecność białek powierzchniowych (S-layer protein), zwiększają siłę oddziaływania z komórkami gospodarza [2,26]. Pojawiły się dowody na to, że wytwarzanie białek powierzchniowych zależy od składu medium hodowlanego. W jednej z prac analizowano wpływ zmian pH oraz temperatury na wytwarzanie białek powierzchniowych przez szczep L. acidophilus ATCC 4356. Badania wykazały, że białka były wytwarzane w warunkach niekorzystnych dla wzrostu bakterii (temperatura, pH), co może wskazywać na ich właściwości ochronne [26]. Z kolei inne badania w podobnych zakresach temperaturowych, wskazują na taką samą ilość białek powierzchniowych syntetyzowanych w ciągu wszystkich faz życia komórki [11]. Prowadzono także badania nad wpływem różnych enzymów na wrażliwość białek z L. acidophilus M92. Wykazano, że warstwa powierzchniowa jest wrażliwa na działanie proteinazy K, ale nie wpływa na nią pankreatyna oraz pepsyna, które są składnikami soku żołądkowego. Potwierdzono również rolę białek warstwy S w mechanizmie adhezji do komórek nabłonka [11]. Wykazano odnawialność warstwy S po jej usunięciu i umieszczeniu komórek w świeżej pożywce MRS. Natomiast nie zaobserwowano wzrostu bakterii Lactobacillus acidophilus M92 w temperaturze 50°C po usunięciu warstwy powierzchniowej, podczas gdy przy jej obecności szczep rozwijał się w tej temperaturze. Wykazano również, że mają one właściwości m.in. ochronne tylko wtedy, gdy znajdują się na komórce (in situ). Może się to wiązać ze zmianą konformacji białek podczas procesu izolacji. Przykładowo, wyizolowane białka wykazywały wrażliwość na trypsynę, podczas gdy były one na nią odporne, gdy znajdowały się na powierzchni komórki [11]. W kolejnych badaniach dotyczących warunków wpływających na funkcjonowanie białek powierzchniowych wykorzystano kilka szczepów L. kefir, gdzie sprawdzano zdolność do antagonizowania toksyn z Clostridium difficile [7]. Przeprowadzono wiele reakcji, podczas których sprawdzano wpływ wysokich temperatur, inhibitorów białkowych oraz swoistych przeciwciał powierzchniowych na wydajność hamowania negatywnych skutków działalności toksyn na komórki nabłonka jelitowego przez białka powierzchniowe bakterii z rodzaju L. kefir. Wykazano, że jedynie swoiste przeciwciała hamowały właściwości białek warstwy S, podczas gdy zarówno wysoka temperatura jak i inhibitory białkowe nie miały większego wpływu, co stanowi kolejne potwierdzenie tego, że temperatura nie wpływa znacząco na blokowanie aktywności białek powierzchniowych [7,11,14].
Oddziaływania białek powierzchniowych z mikroflorą patogenną
Jedną z ważniejszych właściwości jakie mają bakterie probiotyczne jest obniżanie zdolności adhezyjnych, inwazyjnych mikroflory patogennej oraz zmniejszanie aktywności toksyn. Podczas wielu badań udowodniono, że to białka warstwy S są odpowiedzialne za tę właściwość. W jednej publikacji wykazano wpływ bakterii wyizolowanych z kefiru – L. kefir CIDCA 8321, na komórki Salmonella enetritidis [14]. Bakterie L. kefir hamowały inwazję patogenu Salmonella na linię komórkową Caco-2. Zdolność ta pojawiała się tylko po wcześniejszej preinkubacji obydwu mikroorganizmów, podczas której szczep Lactobacillus agregował z komórkami patogenu przez co hamował jego atak na komórki Caco-2. Dla porównania przeprowadzono inkubację komórek Salmonella z Lactobacillus kefir 83113, który nie ma zdolności agregujących i nie zahamował działalności patogenu. Wyniki jednoznacznie nasuwają wniosek, że agregacja z patogenem ma szczególne znaczenie podczas ochrony komórkowej, przez maskowanie, czy też blokowanie powierzchni komórek patogennych. Wspomniane wyniki sugerują rozpoczęcie badań w kierunku rozwoju produktów probiotycznych przeciwdziałających zakażeniom wywołanym przez szczepy Salmonella u ludzi i zwierząt [14,16]. W innych badaniach analizowano oddziaływanie białek powierzchniowych wyizolowanych z różnych szczepów bakterii z gatunku Lactobacillus kefir z toksynami bakteryjnymi Clostridium difficile TcdA oraz TcdB, które wywołują zmiany w cytoszkielecie atakowanej komórki. W kilku przypadkach zaobserwowano oddziaływanie między białkami powierzchniowymi wyizolowanymi z bakterii Lactobacillus a oczyszczonymi toksynami oraz znaczne obniżenie aktywności toksyn [7]. Podobne wyniki otrzymano dla szczepów z gatunków Lactobacillus gasseri, L. plantarum oraz L. acidophilus, które miały zdolność hamowania in vitro adhezji komórek Escherichia coli oraz Salmonella Typhimurium do komórek nabłonka jelitowego [4,6,10,32]. Badano również wpływ Lactobacillus na adhezję do komórek nabłonka jelitowego bakterii z rodzaju Shigella – patogenu, który jest jednym z głównych czynników chorób przewodu pokarmowego. Wykazano, że wyizolowane z tradycyjnej, fermentowanej, chińskiej żywności, szczepy L. paracasei subp. paracasei M5-L, L. rhamnosus J10-L oraz L. casei Q8-L i ich białka powierzchniowe w znacznym stopniu hamowały adhezję Shigella sonnei do komórek HT29 [16,53].
Najnowsze doniesienia opisują białka powierzchniowe jako środek zapobiegawczy przed infekcją JUNV (wirus Junin). W doświadczeniach wykorzystano białka warstwy S wyizolowane ze szczepu L. acidophilus ATCC 4353 (mający status GRAS), komórki linii 3T3 ekspresjonujące receptor DC-SIGN lub jego homolog L-SIGN oraz komórki linii Vero do badań in vitro. Arenawirus JUNV jest czynnikiem etiologicznym argentyńskiej gorączki krwotocznej, choroby, która wywołuje powikłania neurologiczne oraz krwotoczne. Atakuje on komórki zawierające białko receptorowe (DC-SIGN) rozpoznające glikany mannozy i fruktozy znajdujące się na powierzchni mikroorganizmów oraz wirusów. Badano również homolog DC-SIGN, receptor L-SIGN, o podobnych funkcjach oraz zbliżonej sekwencji aminokwasowej. Wyniki wykazały, że białka powierzchniowe z L. acidophilus ATCC 4353 powodowały redukcję infekcji przez JUNV na komórkach z receptorem DC-SIGN. Nie zaobserwowano inhibicji na komórkach mających receptor L-SIGN. Przeprowadzono również eksperyment, w którym białka powierzchniowe Lactobacillus były dodane po zainfekowaniu komórek. Nie zaobserwowano zahamowania, co może świadczyć o tym, że białka warstwy S biorą udział we wczesnym stadium replikacji wirusa. Zauważono również, iż wraz ze wzrostem koncentracji białek powierzchniowych, maleje poziom infekcji wirusem Junin [15,32].
W innych doniesieniach literaturowych wykazano wpływ rozpuszczalnych białek powierzchniowych L. rhamnosus p75 oraz p40 na wzrost komórek nabłonkowych jelita. W chorobie zapalnej jelit (IBD – inflammatory bowel disease) dochodzi do zaburzenia integralności komórek nabłonka poprzez infiltrację komórek lub ich apoptozę. W eksperymentach wykorzystano szczep Lactobacillus rhamnosus GG (LGG), który jest najlepiej przeanalizowanym szczepem pod kątem IBD. Wykazano, że LGG zapobiega apoptozie indukowanej przez cytokiny w ludzkich i mysich nabłonkach jelitowych przez aktywację kinazy Akt i inhibicję MAPK (p38 mitogen-activated protein kinase). Wyizolowano dwa białka z L. rhamnosus GG – p75 i p40 i wykazano, że powodowały one inhibicję apoptozy i wspomagały wzrost komórek nabłonka, aktywowały kinazę Akt oraz redukowały aktywność czynnika martwicy nowotworowej (TNF) [51].
Zgromadzona dotychczas wiedza na temat właściwości białek warstwy powierzchniowej, to pierwszy krok w kierunku wykorzystania ich w celach terapeutycznych oraz zapobiegania infekcji przewodu pokarmowego. Właściwości immunomodulacyjne, adhezyjne oraz hamujące aktywność mikroflory patogenicznej mogą być wykorzystane do stworzenia doustnych szczepionek. Ponadto białka powierzchniowe mogą być wykorzystane nie tylko w mikrobiologii lekarskiej, ale również w dziedzinach biotechnologicznych oraz w ochronie środowiska, np. poprzez zwiększenie wydajności usuwania metali ciężkich z wody. Dzięki temu uda się zmniejszyć prawdopodobieństwo zakażenia metalami oraz zwiększą się możliwości leczenia i zapobiegania infekcjom układu pokarmowego. Jest to niezwykle istotne, gdyż nieleczone zapalenia jelitowe mogą prowadzić do zmian onkologicznych, a w konsekwencji nawet do śmierci człowieka.
PIŚMIENNICTWO
[1] Ashida N., Yanagihara S., Shinoda T., Yamamoto N.: Characterization of adhesive molecule with affinity to Caco-2 cells in Lactobacillus acidophilus by proteome analysis. J. Biosci. Bioeng., 2011; 112: 333-337
[PubMed]
[2] Beganović J., Frece J., Kos B., Lebos Pavunc A., Habjanič K., Sušković J.: Functionality of the S-layer protein from the probiotic strain Lactobacillus helveticus M92. Antonie van Leeuwenhoek, 2011; 100: 43-53
[PubMed]
[3] Bergey’s Manual of Systematic Bacteriology, wyd. 2, t. 3: The Firmicutes. Springer, New York 2009, 465-510
[4] Bernet M.F., Brassart D., Neeser J.R., Servin A.L.: Lactobacillus acidophilus LA1 binds to cultured human intestinal cell lines and inhibits cell attachment and cell invasion by enterovirulent bacteria. Gut, 1994; 35: 483-489
[PubMed] [Full Text HTML] [Full Text PDF]
[5] Błaszczyk U.: Bakteriocyny – właściwości i zastosowanie. Laboratorium – Przegląd Ogólnopolski, 2008; 10: 28-32
[Full Text PDF]
[6] Candela M., Perna F., Carnevali P., Vitali B., Ciati R., Gionchetti P., Rizzello F., Campieri M., Brigidi P.: Interaction of probiotic Lactobacillus and Bifidobacterium strains with human intestinal epithelial cells: adhesion properties, competition against enteropathogens and modulation of IL-8 production. Int. J. Food Microbiol., 2008; 125: 286-292
[PubMed]
[7] Carasi P., Trejo F.M., Pérez P.F., De Antoni G.L., Serradell Mde L.: Surface proteins from Lactobacillus kefir antagonize in vitro cytotoxic effect of Clostridium difficile toxins. Anaerobe, 2012; 18: 135-142
[PubMed]
[8] Castaldo C., Vastano V., Siciliano R.A., Candela M., Vici M., Muscariello L., Marasco R., Sacco M.: Surface displaced alfa-enolase of Lactobacillus plantarum is a fibronectin binding protein. Microb. Cell Fact., 2009; 8: 14
[PubMed] [Full Text HTML] [Full Text PDF]
[9] Chen X., Chen Y., Li X., Chen N., Fang W.: Characterization of surface layer proteins in Lactobacillus crispatus isolate ZJ001. J. Microbiol. Biotechnol., 2009; 19: 1176-1183
[PubMed] [Full Text PDF]
[10] Coconnier M.H., Bernet M.F., Kernéis S., Chauviere G., Fourniat J., Servin A.L.: Inhibition of adhesion of enteroinvasive pathogens to human intestinal Caco-2 cells by Lactobacillus acidophilus strain LB decreases bacterial invasion. FEMS Microbiol. Lett., 1993; 110: 299-305
[PubMed]
[11] Frece J., Kos B., Svetec I. K., Zgaga Z., Mrša V., Šuškovic J.: Importance of S-layer proteins in probiotic activity of Lactobacillus acidophilus M92. J. Appl. Microbiol., 2005; 98: 285-292
[PubMed]
[12] Gardinera G.E., Heinemanna C., Barojab M.L., Brucea A. W., Beuerman D., Madrenas J., Reid G.: Oral administration of the probiotic combination Lactobacillus rhamnosus GR-1 and L. fermentum RC-14 for human intestinal applications. Int. Dairy J., 2002; 12: 191-196
[13] Gerbino E., Mobili P., Tymczyszyn E., Fausto R., Gómez-Zavaglia A.: FTIR spectroscopy structural analysis of the interaction between Lactobacillus kefir S-layers and metal ions. J. Mol. Structure, 2011; 987: 186-192
[14] Golowczyc M.A., Mobili P., Garrote G.L., Abraham A.G., De Antoni G.L.: Protective action of Lactobacillus kefir carrying S-layer protein against Salmonella enterica serovar Enteritidis. Int. J. Food Microbiol., 2007; 118: 264-273
[PubMed]
[15] Gómez R.M., Jaquenod de Giusti C., Sanchez Vallduvi M.M., Frik J., Ferrer M.F., Schattner M.: Junín virus. A XXI century update. Microbes Infect., 2011; 13: 303-311
[PubMed]
[16] Grajek W., Olejnik A., Stanaszek K.: Kultury komórkowe nabłonka jelitowego jako model do badania transportu transnabłonkowego. Biotechnologia, 2006; 73: 148-165
[Full Text PDF]
[17] Granato D., Bergonzelli G.E., Pridmore R.D., Marvin L., Rouvet M., Corthésy-Theulaz I.E.: Cell surface-associated elongation factor Tu mediates the attachment of Lactobacillus johnsonii NCC533 (La1) to human intestinal cells and mucins. Infect. Immun., 2004; 72: 2160-2169
[PubMed] [Full Text HTML] [Full Text PDF]
[18] Górska S., Grycko P., Rybka J., Gamian A.: Exopolysaccharides of lactic acid bacteria: structure and biosynthesis. Postępy Hig. Med. Dośw., 2007; 61: 805-818
[PubMed] [Full Text HTML] [Full Text PDF]
[19] Górska S., Jarząb A., Gamian A.: Bakterie probiotyczne w przewodzie pokarmowym człowieka jako czynnik stymulujący układ odpornościowy. Postępy Hig. Med. Dośw., 2009; 63: 653-667
[PubMed] [Full Text HTML] [Full Text PDF]
[20] Hoesl C.E., Altwein J.E.: The probiotic approach: An alternative treatment option in urology. Eur. Urol., 2005; 47: 288-296
[PubMed]
[21] Hynönen U.: Structural and functional characterization of the surface layer protein of Lactobacillus brevis ATCC 8287 – academic dissertation. Department of Basic Veterinary Sciences, University of Helsinki, 2009
[Full Text PDF]
[22] Jakava-Viljanen M.: Characterization of porcine-specific surface (S-) layer protein carrying Lactobacillus species, S-layer proteins and the adhesin of Escherichia coli F18 fimbriae – potential applications for veterinary medicine – academic dissertation. Food Safety Authority Evira, Helsinki 2007
[Full Text PDF]
[23] Kalliomäki M., Salminen S., Poussa T., Arvilommi H., Isolauri E.: Probiotics and prevention of atopic disease: 4-year follow-up of a randomised placebo-controlled trial. Lancet, 2003; 361: 1869-1871
[PubMed]
[24] Kang H.J., Baker E.N.: Structure and assembly of Gram-positive bacterial pili: unique covalent polymers. Curr. Opin. Struct. Biol., 2012; 22: 200-207
[PubMed]
[25] Kaur I.P., Chopra K., Saini A.: Probiotics: potential pharmaceutical applications. Eur. J. Pharm. Sci., 2002; 15: 1-9
[PubMed]
[26] Khaleghi M., Kermanshahi R.K.: Effect of environmental stresses on s-layer production in Lactobacillus acidophilus ATCC 4356. Adv. Appl. Biotechnol.; 2012, 209-225
[Full Text HTML] [Full Text PDF]
[27] Kiessling G., Schneider J., Jahreis G.: Long-term consumption of fermented dairy products over 6 months increases HDL cholesterol. Eur. J. Clin. Nutr., 2002; 56: 843-849
[PubMed] [Full Text HTML] [Full Text PDF]
[28] Kinoshita H., Uchida H., Kawai Y., Kawasami T., Wakahara N., Matsuo H., Watanabe M., Kitazawa H., Ohnuma S., Miura K., Horii A., SaitoT.: Cell surface Lactobacillus plantarum LA 318 glyceraldehyde-3-phosphate dehydrogenase (GAPDH) adheres to human colonic mucin. J. Appl. Microbiol., 2008;104: 1667-1674
[PubMed]
[29] Kinoshita H., Wakahara N., Watanabe M., Kawasaki T., Matsuo H., Kawai Y., Kitazawa H., Ohnuma S., Miura K., Horii A., Saito T.: Cell surface glyceraldehyde-3-phosphate dehydrogenase (GAPDH) of Lactobacillus plantarum LA 318 recognizes human A and B blood group antigens. Res. Microbiol., 2008; 159: 685-691
[PubMed]
[30] Lebeer S., Vanderleyden J., De Keersmaecker S.C.: Genes and molecules of Lactobacilli supporting probiotic action. Microbiol. Mol. Biol. Rev., 2008; 12: 728-764
[PubMed] [Full Text HTML] [Full Text PDF]
[31] Maragkoudakisa P.A., Zoumpopouloua G., Miarisa C., Kalantzopoulosa G., Potb B., Tsakalidou E.: Probiotic potential of Lactobacillus strains isolated from dairy products. Int. Dairy J., 2006; 16: 189-199
[32] Martínez M.G., Acosta M.P., Candurra N.A., Ruzal S.M.: S-layer proteins of Lactobacillus acidophilus inhibits JUNV infection. Biochem. Biophys. Res. Commun., 2012; 422: 590-595
[PubMed]
[33] Matijašić, B.B., Narat M., Peternel M.Z., Rogelj I.: Ability of Lactobacillus gasseri K7 to inhibit Escherichia coli adhesion in vitro on Caco-2 cells and ex vivo on pigs’ jejunal tissue. Int. J. Food Microbiol., 2006; 107: 92-96
[PubMed]
[34] Mobili P., Londero A., Maria T.M., Eusebio M.E., De Antoni G.L., Fausto R., Gómez-Zavaglia A.: Characterization of S-layer proteins of Lactobacillus by FTIR spectroscopy and differential scanning calorimetry. Vibrational Spectroscopy, 2009; 50: 68-77
[35] Mombelli B., Gismondo M.R.: The use of probiotics in medical practice. Int. J. Antimicrob. Agents, 2000; 16: 531-536
[PubMed]
[36] Pollmann L., Raff J., Merroun M., Fahmy K., Selenska-Pobell S.: Metal binding by bacteria from uranium mining waste piles and its technological applications. Biotechnol. Adv., 2006; 24: 58-68
[PubMed]
[37] Rojas M., Ascencio F., Conway P. L.: Purification and characterization of a surface protein from Lactobacillus fermentum 104R that binds to porcine small intestinal mucus and gastric mucin. Appl. Environ. Microbiol., 2002; 5: 2330-2336
[PubMed] [Full Text HTML] [Full Text PDF]
[38] Roos S., Jonsson H.: A high-molecular-mass cell-surface protein from Lactobacillus reuteri 1063 adheres to mucus components. Microbiology, 2002; 148: 433-442
[PubMed] [Full Text HTML] [Full Text PDF]
[39] Saad N., Urdaci M., Vignoles C., Chaignepain S., Tallon R., Schmitter J.M., Bressollier P.: Lactobacillus plantarum 299v surface-bound GAPDH: a new insight into enzyme cell walls location. J. Microbiol. Biotechnol., 2009; 19: 1635-1643
[PubMed] [Full Text PDF]
[40] Sánchez B., Schmitter J.M., Urdaci M.C.: Identification of novel proteins secreted by Lactobacillus rhamnosus GG grown in de Mann-Rogosa-Sharpe broth. Lett. Appl. Microbiol., 2009; 48: 618-622
[PubMed]
[41] Schär-Zammaretti P., Ubbink J.: The cell wall of lactic acid bacteria: surface constituents and macromolecular conformations, Biophys. J., 2003; 85: 4076-4092
[PubMed] [Full Text HTML] [Full Text PDF]
[42] Seweryn E., Pietkiewicz J., Szamborska A., Gamian A.: Enolaza na powierzchni komórek eukariota i prokariota jako receptor plazminogenu ludzkiego. Postępy Hig. Med. Dośw., 2007; 61: 672-682
[PubMed] [Full Text HTML] [Full Text PDF]
[43] Strzelecki J., Sadowy E., Hryniewicz W.: Białka powierzchniowe enterokoków odpowiedzialne za oddziaływanie z tkankami gospodarza. Postępy Mikrobiol., 2011; 50, 31-42
[Full Text PDF]
[44] Todorov S.D.: Bacteriocins from Lactobacillus plantarum – production, genetic organization and mode of action. Braz. J. Microbiol., 2009; 40: 209-221
[Full Text HTML] [Full Text PDF]
[45] Todorov S.D., Vaz-Velho M., Gombossy de Melo Franco B.D., Holzapfel W.H.: Partial characterization of bacteriocins produced by three strains of Lactobacillus sakei, isolated from salpicao, a fermented meat product from North-West of Portugal. Food Control, 2013; 30: 111-121
[Full Text HTML] [Full Text PDF]
[46] Tripathi P., Beaussart A., Andre G., Rolain T., Lebeer S., Vanderleyden J., Holsc P., Dufrene Y.F.: Towards a nanoscale view of lactic acid bacteria. Micron, 2012; 43: 1323-1330
[PubMed]
[47] Van Pijkeren J.P., Canchaya C., Ryan K.A., Li Y., Claesson M.J., Sheil B., Steidler L., O’Mahony L., Fitzgerald G.F., Van Sinderen D., O’Toole P.W.: Comparative and functional analysis of sortase-dependent proteins in the predicted secretome of Lactobacillus salivarius UCC118. Appl. Environ. Microbiol., 2006; 72: 4143-4153
[PubMed] [Full Text HTML] [Full Text PDF]
[48] Ventura M., Jankovic I., Walker D.C., Pridmore R.D., Zink R.: Identification and characterization of novel surface proteins in Lactobacillus johnsonii and Lactobacillus gasseri. Appl. Environ. Microbiol., 2002; 68: 6172-6181
[PubMed] [Full Text HTML] [Full Text PDF]
[49] Vliegenthart J.F., van den Berg D.J., Robijn G.W., Janssen A.C., Giuseppin M.L., Vreeker R., Kamerling J.P., Ledeboer A.M.: Production of a novel extracellular polysaccharide by Lactobacillus sake 0-1 and characterization of the polysaccharide. Appl. Environ. Microbiol., 1995; 64: 2840-2844
[PubMed] [Full Text PDF]
[50] Wang B, Wei H., Yuan J., Li Q., Li Y., Li N., Li J.: Identification of a surface protein from Lactobacillus reuteri JCM1081 that adheres to porcine gastric mucin and human enterocyte-like HT-29 cells. Curr. Microbiol., 2008; 57: 33-38
[PubMed] [Full Text HTML] [Full Text PDF]
[51] Yan F., Cao H., Cover T. L., Whitehead R., Washington M.K., Polk D.B.: Soluble proteins produced by probiotic bacteria regulate intestinal epithelial cell survival and growth. Gastroenterology, 2007; 132: 562-575
[PubMed] [Full Text HTML] [Full Text PDF]
[52] Yesovitch R., Cohen A., Szilagyi A.: Failure to improve parameters of lactose maldigestion using the multiprobiotic product VSL3 in lactose maldigesters: a pilot study. Can. J. Gastroenterol., 2004; 18: 83-86
[PubMed]
[53] Zhang Y.C., Zhang L.W., Tuo Y.F., Guo C.F., Yi H.X., Li J.Y., Han X., Du M.: Inhibition of Shigella sonnei adherence to HT-29 cells by lactobacilli from Chinese fermented food and preliminary characterization of S-layer protein involvement. Res. Microbiol., 2010; 161: 667-672
[PubMed]
Autorzy deklarują brak potencjalnych konfliktów interesów.