
Wpływ kotyniny na komórki linii niedrobnokomórkowego raka płuc A549
Jakub Nowak 1 , Alina Grzanka 2 , Maciej Gagat 2 , Agnieszka Żuryń 2Streszczenie
Celem niniejszej pracy była próba określenia wpływu kotyniny na linię komórkową A549. Komórki linii A549 poddano 24-godzinnej inkubacji z kotyniną w dawkach 18 ng/ml i 36 ng/ml. Komórki kontrolne inkubowano w podobnych warunkach bez dodatku kotyniny. Oznaczono ich żywotność, barwiono hematoksyliną, zatopiono w eponie, oglądano w mikroskopie elektronowym oraz zastosowano metodę fluorescencyjną w celu oceny F-aktyny. W wyniku działania kotyniny dochodzi do zmian cytoszkieletu, o czym świadczą zmiany organizacji F-aktyny. Zaobserwowano również, że komórki linii A549 prawdopodobnie ulegają śmierci w procesie apoptozy oraz katastrofy mitotycznej. Ponadto wyniki uzyskane na poziomie ultrastrukturalnym sugerują, że kotynina działa nie tylko na jądro komórkowe i wpływa na kształt komórki poprzez działanie na cytoszkielet aktynowy, ale również odgrywa istotną rolę w zmianach, które obejmują pozostałe organella komórkowe. Dotychczas oceniano przede wszystkim wpływ działania składników dymu papierosowego, głównie nikotyny, stąd zasadne było podjęcie badań z wykorzystaniem kotyniny. Uzyskane wyniki pozwalają przypuszczać, że kotynina poprzez F-aktynę może wpływać na wielkość i kształt komórek linii niedrobnokomórkowego raka płuc. Sugeruje się, że komórki linii A549 w wyniku działania kotyniny mogą ulegać apoptozie i katastrofie mitotycznej.
Słowa kluczowe:kotynina • niedrobnokomórkowy rak płuc • aktyna • linia komórkowa A549
Summary
The aim of this investigation was to determine the influence of cotinine on the non-small-cell lung cancer line A549. The material for the study was the A549 cell line. The cells were subjected to 24-h incubation with cotinine at doses of 18 and 36 ng/ml. Control cells were incubated under analogous conditions without the addition of cotinine. Cell viability was determined and the cells were Mayer hematoxylin stained, embedded in epon, and observed under an electron microscope. A fluorescent method was used to evaluate F-actin. Cotinine’s action resulted in alteration of the cytoskeleton, with changes in the organization of F-actin. Also observed was A549 cell death, presumably due to the activation of apoptotic and mitotic catastrophe pathways. Moreover, the results at the ultrastructural level suggest that cotinine influences not only the nucleus and cell shape by acting on the actin cytoskeleton, but also plays a crucial role in changes which include the remaining cellular organelles. Until now, research has been focused on determining the action of constituents of cigarette smoke, mainly nicotine. From this point of view, studies on cotinine seem justified. Results of the study allow us to suppose that cotinine, through F-actin, can influence the size and shape of non-small-cell lung cancer cells. There are some suggestions that cells of the A549 cell line can undergo death through apoptotic and mitotic catastrophe pathways as a result of cotinine’s action.
Key words:cotinine • non-small-cell lung cancer • actin • A549 cell line
Abbreviations:
COT – cotinine; CSE – cigarette smoke extract; CYP2A6 – cytochrome P450 2A6; DAPI – 4’,6- diaminoino-2-fenylindol; FMO – flavin-containing monooxygenase; HBSS – Hank’s Buffered Salt Solution; HE – H&E (hematoxylin and eosin) stain; NIC – nicotine; NSCLC – non-small-cell lung cancer; RT – room temperature; TGF-β – tumor growth factor β; TRITC – tetramethylrhodamineisothyocyanate; UV – ultraviolet; WHO – World Health Organization
INTRODUCTION
Lung cancer is the most frequent malignant tumor, causing ca. 1.3 million deaths all around the world [26]. At present it is believed that the reason for the high incidence of lung cancer is long-term exposure to airborne carcinogens, mostly tobacco smoke. Over 3000 substances, known carcinogens and including vinyl chloride, methane, alkaloids (nicotine), aromatic hydrocarbons, and heavy metals, have been identified in the composition of cigarette smoke. In patients with lung cancer who have never smoked, some factors such as passive exposure to tobacco smoke (secondhand smoking) and genetic factors have been considered [18]. Cigarette smoking is one of the most significant factors responsible for the increased risk of morbidity due to non-small-cell lung cancer, which comprises about 80% of lung cancers. The concentration of cotinine, the main metabolite of nicotine, depends on the degree of the organism’s exposure to cigarette smoke [21]. It was established that about 70-80% of the nicotine which enters the human organism undergoes transformation to cotinine. This process occurs above all in the liver. Cotinine is produced in a two-stage process in which cytochrome P450 2A6 (CYP2A6) and cytoplasmic aldehyde oxidase are involved [8,16]. The effects of lung cancer treatment are unsatisfactory, which is reflected in statistical data on patient survival rates and prognoses [26,27]. At the moment the fight against addiction to cigarette smoking has the greatest significance in decreasing the number of deaths caused by lung cancer. However, there is no one universal therapy which would give chances of cure. It is possible that the interrelation of cigarette smoking, cotinine concentration, and the influence of cotinine on non-smallcell lung cancer cells would allow a better understanding of the mechanisms connected with this type of cancer in the future. Studies involving cotinine in the A549 cell line would allow us to create a research model for understanding the mechanisms of immortalization and also to create medications that inhibit and eliminate the cells responsible for the malignancy of lung tumors in the future.
MATERIAL AND METHODS
Cell culture
The study material was the A549 cell line (non-smallcell lung cancer line ATCC CL 185), kindly provided by Kopiński, Ph.D. (Nicolaus Copernicus University, Collegium Medicum in Bydgoszcz). The cells were grown in DMEM+GlutaMAX (Gibco) supplemented with 10% (m/v) fetal bovine serum (Gibco) and antibiotics (a solution of penicillin and streptomycin) at 37°C in a 5% CO2 atmosphere with stable humidity. Before starting the culture, the density of cells was 5×105 cells/ml. The cells were incubated with cotinine for 24 h. The concentrations of cotinine used were 18 and 36 ng/ml. Control cells were grown under the same conditions but without the addition of cotinine to the medium. The experiment was repeated three times.
Assessment of A549 cell viability
Five to ten µl of A549 cell suspension in culture medium was mixed on a microscope slide with a freshly prepared solution of 2% tripan blue diluted in 0.9% NaCl (1:1 solution) and covered with a coverslip. The number of dying blue-stained cells was assessed after 3 minutes.
HE staining: examination of A549 cell line morphology in light microscopy
Cells were fixed with 4% paraformaldehyde in HBSS (15 min, RT) and afterwards rinsed with PBS (3×5 min, RT). Then the cells were embedded in 0.1 M glycine solution (5 min, RT). Following a double rinsing in PBS, the cells were stained with Mayer’s hematoxylin (5 min, RT). Then the cells were rinsed for 20 minutes under running water. After that the cells were dehydrated in a series of alcohols and xylenes. The fixed preparations were examined using an Eclipse E800 microscope (Nikon, Tokyo, Japan) with computer imaging system analysis (NIS-Elements software version 4.20, Nikon Instruments).
Immunofluorescence: analysis of F-actin organization in fluorescence microscopy
Cells were fixed with 4% paraformaldehyde in HBSS (15 min, RT) and afterwards rinsed with PBS (3×5 min, RT). Then the cells were incubated with 0.1 M glycine (5 min, RT) and rinsed in PBS (3×5 min, RT). To stain the material, phalloidin conjugated with a derivative of rodamin (phalloidin/TRITC, Sigma) was used. The cells were incubated in darkness with 10 µM of stock phalloidin/TRITC solution diluted 1:5 in 20% methanol (20 min, RT). Nuclei were counterstained with DAPI (Sigma) at a dilution of 1:25,000 (10 min, RT). In the last stage, the material was embedded in Gelvatol (Monsanto). For a control, serum derived from unimmunized mice instead of the first antibody (phalloidin/TRITC) was used to confirm reaction specificity. F-actin was examined in an Eclipse E800 microscope with a Y-FL fluorescence attachment (Nikon, Tokyo, Japan). Computer analysis of fluorescent imaging was done with the use of NIS-Elements software version 4.20 (Nikon Instruments).
Electron microscopy: ultrastructural analysis of A459 cells
For ultrastructural analysis, cells were fixed for 1 hour in 3.6% glutaralehyde and then moved to 0.1 M cacodylate buffer (pH 7.4). Following postfixing for 1.5 hour with 2% osmium tetrooxide in 0.1M cacodylate buffer, the material was dehydrated in a graded series of alcohols and embedded in Epon 212. Semi-thin sections were made of all preparations using a Reichert OmU3 ultramicrotome. Parts of the material selected from the semi-thin sections were cut into ultrathin sections and then stained with uranyl acetate and lead citrate. The prepared material was analyzed with n JEM 100 CX electron microscope (JEOL, Tokyo, Japan).
Statistical analysis
All results were subjected to statistical analysis. After 24 hours of treatment, the number of dead cells and the total number of cells were analyzed. Quantitative experiments were analyzed by the Mann-Whitney U test. Statistical analysis was carried out with Graph Pad Prism software version 4.0. Diagrams were made with MS Excel.
RESULTS
A549 cell line viability after cotinine treatment
Statistical analysis did not show statistically significant differences in the average percentage of surviving A549 cells after treatment with the two concentrations of cotinine, which was confirmed by data obtained from counting dead cells after 24 hours of incubation with cotinine at doses of 18 and 36 ng/ml (viability: 91.09% and 92.9%, respectively, compared with 90.05% in control cells, Figure 1).
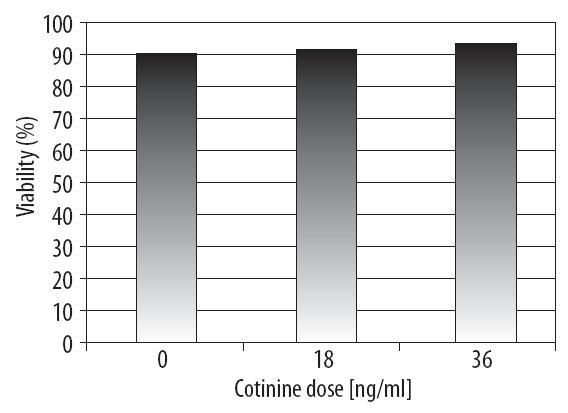
Figure 1. The influence of cotinine dose on the average percentage of surviving A549 cells
However, there were statistically significant differences in the total number of cells after 24 hours of incubation with cotinine. The quantitative studies showed that the control had the highest number of NSCLC cells (ca. 68×104). After COT treatment at a dose of 18 ng/ml, the number of cells decreased (to ca. 45×104), whereas the lowest number of cells was observed at 36 ng/ml (Figure 2).
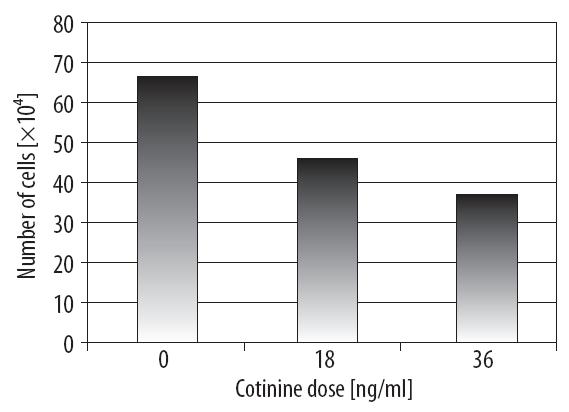
Figure 2. The influence of cotinine dose on the total number of cells
Light microscopic examination of cell morphology
Changes in shape, number, and size of the cotinine-treated cells were observed compared with the controls (Figure 3A). HE staining seemed to reveal decreased total numbers of cells, increased intracellular spaces, and cell shrinkage occurring dose dependently. Cells treated with cotinine showed a tendency to form clusters (Figures 3B, 3C). In some cells after COT treatment, mostly at a dose of
36 ng/ml, multiplied nucleolus were observed in comparison with the control cells as well as characteristic blebs, probably containing chromatin (Figure 3C).
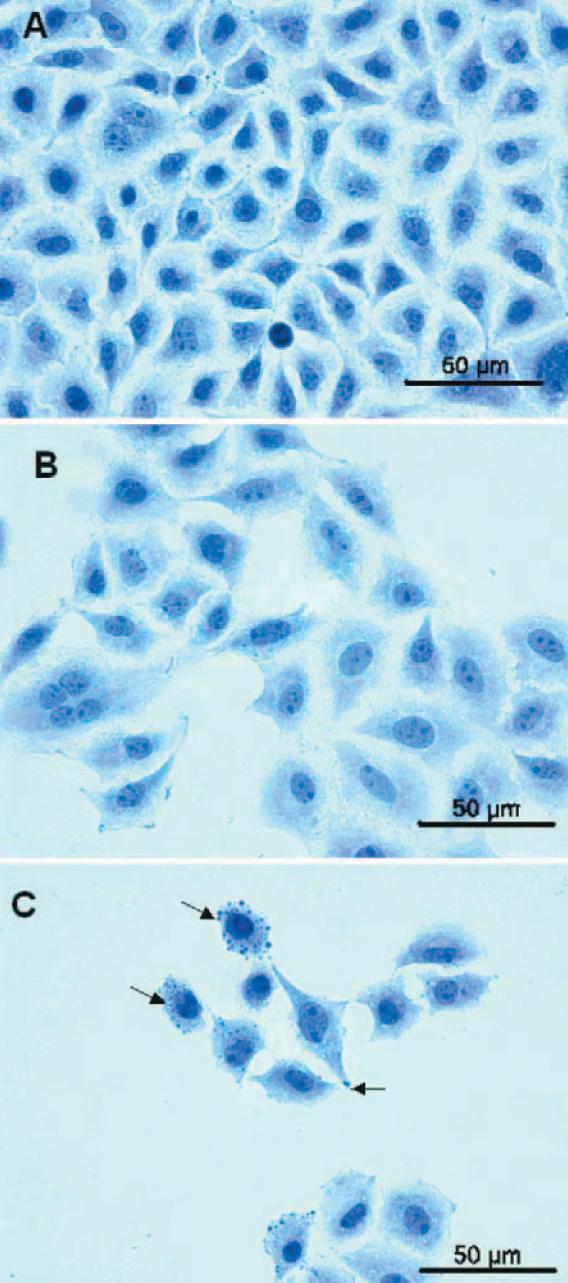
Figure 3. Light microscopy; A – Control: A549 cells incubated without cotinine. Close cell-cell contacts were observed, B – A549 cells incubated with cotinine at a dose of 18 ng/ml. Increased intracellular spaces and multiplied nucleolus in the area of the nucleus were observed (arrows), C – A549 cells incubated with cotinine at a dose of 36 ng/ml. Cells with condensed chromatin and apoptotic blebs were observed (arrows)
Analysis of F-actin organization in fluorescence microscopy
Fluorescence microscopic studies showed the effect of cotinine on F-actin distribution in the A549 cells. In the control cells, F-actin was evenly distributed. The nuclei were located in the central region of the cell and had regular shapes (Figure 4A). However, after 24 hours of treatment with COT (18 and 36 ng/ml) they were shrunken (Figures 4B, 4C). Besides cells shrinkage, large cells appeared with strongly developed stress fibers. In the area of these cells, lobe-shaped nuclei and micronuclei were observed (Figure 4B). In the majority of both types of cells, i.e. the shrunken and large cells, F-actin accumulation was observed in the central part of the cell, in the place of the nucleus.

Figure 4. Fluorescence microscopy; A – Control: A549 cells incubated without cotinine (phalloidin/TRITC – F-actin, DAPI – nuclei). Regular-shaped nuclei in the central part of the cell (arrows) and evenly distributed F-actin in the area of the cell were observed, B – A549 cells incubated with cotinine at a dose of 18 ng/ml (phalloidin/TRITC – F-actin, DAPI – nuclei). Micronuclei in the area of the cell (arrow), C – A549 cells incubated with cotinine at a dose of 36 ng/ml (phalloidin/TRITC – F-actin, DAPI – nuclei). Shrunken cells were observed
Ultrastructural analysis of A459 cells
Light microscopy studies were supplemented by ultrastructural analysis of the A549 cells, which allowed for a more accurate observation of changes in the cell area after COT treatment. The most significant changes were observed after COT treatment at a dose of 36 ng/ml. At the electron microscopic level, round nuclei were observed in the control cells (Figure 5A). However, after using COT, changes in the shape of the nucleus occurred, i.e. the nuclei became dose-dependently less regular and more lobe-shaped (Figures 5B, 5C). With increasing dose, a greater number of nucleoli, mostly in the cells treated with the higher dose of COT, was also observed (Figure 5C). Changes in the cytoplasmic area were also observed, such as damage to mitochondria and morphological alterations of the endoplasmic reticulum. Mitochondria were swollen, with a visible disappearance of cristae. Endoplasmic reticulum canals were dilated. In these cells there were increased numbers of lysosomes compared with the control cells (Figure 5C). Studies at the electron microscopy level showed that all the above changes we observed intensified with increasing dose of COT.

Figure 5. Electron microscopy; A – Control: A549 cells incubated without cotinine. Regularly shaped nucleus and lack of changes in cellular organelles were observed. Magnification ×12,000, B – A549 cells incubated with cotinine at a dose of 18 ng/ml. Irregularly shaped nucleus, swollen mitochondria (arrow a) and lysosomes (arrow b) were observed. Magnification ×13,000, C – A549 cells incubated with cotinine at a dose of 36 ng/ml. Nucleoli in the area of the nucleus (arrow a), dilated endoplasmic reticulum canals (arrow b), swollen mitochondria (arrow c), and lysosomes (arrow d) were observed. Magnification ×12,000
DISCUSSION
The human non-small-cell lung cancer cell line A549 was derived in the early 1970s. In the 1980s it was used above all in investigations with herpes simplex virus [19]. Since then, this cell line has been found to be useful in many different studies. Foster et al. [12] pointed out that due to their properties, which are characteristic of pulmonary alveolar type II cells, non-small-cell lung cancer cells constitute an attractive research model for the characterization of the metabolism and mechanisms of drug transport to the pulmonary epithelium. Nowadays, the NSCLC A549 cell line is used in studies on the influence of chemical agents on cancerous processes. Xiao-hua et al. [25] demonstrated that cantharidin exerts an inhibitory effect on the proliferation of A549 cells. Tai et al. [20] used A549 cells in experiments with an enzyme responsible for biological prostaglandin inactivation. The NSCLC A549 cell line also constituted a model in research conducted by Katakura which was related to the cellular senescence program and its application to anticancer therapy. Katakura showed that TGF-β is a cellular senescence-inducing factor. He identified 86 independent senescence-associated genes in A549-derived cells and established novel cell lines from A549 cells [10].
Many studies used A549 cells as a model for establishing the influence of elements of cigarette smoke. Hoshino et al. showed that cigarette smoke extract at a concentration of 5% and lower initiates apoptosis in the A549 cell line. However, at higher concentration (10%) it is responsible for necrosis [7]. It is necessary to remember that cigarette smoke contains 4000 different agents, and each of them appears in different concentrations depending on the smoking conditions and the kind of cigarette [2]. Similar results were obtained by Ramage et al. which showed an increased number of apoptotic cells after cigarette smoke and nicotine treatment [17]. The influence of cigarette smoke on cellular senescence in non-small-cell lung cancer was studied by Tsuji et al. Using a liquid cigarette smoke extract solution, they demonstrated that a sublethal dose of cigarette smoke causes the appearance of a phenotype indicative of cellular senescence in pulmonary alveoli cells [22]. However, Klein et al. suggested the occurrence of a mitotic catastrophe as a response to stress conditions, which can also lead to cellular senescence [11].
In recent years, the NSCLC A549 cell line has become the model cell line for testing many cancer drugs and agents that cause lung cancer. Statistical data show that smoking cigarettes is the main cause of morbidity due to lung cancer [26]. It is known that nicotine, one of the main constituents of cigarette smoke, undergoes transformation to cotinine with the participation of cytochrome P450 [1,8,23,29]. In vitro studies on the A549 cell line showed that nicotine causes an increase in the number of apoptotic cells [7,17]. However, Dasgupta et al. suggested that nicotine inhibits apoptotic processes induced by taxol, cisplatin, and gemcytabin [4]. There are also observations that nicotine inhibits apoptosis induced by opioids, etoposide, cisplatin, and UV [9,13,14,28].
In the present study we investigated the influence of cotinine, the main metabolite of nicotine synthesized in the organism, on the non-small-cell lung cancer A549 cell line. Research to date has been focused on evaluating the actions of constituents of cigarette smoke, mainly nicotine [5,7,17,22]. Cotinine, as a derivative of nicotine, is a good object for studies related to its influence on cells derived from the human organism, especially because its concentration in smokers is significantly higher [1,3,8,12,16,23,29]. >From this point of view, studies that aim to explain a relationship between cotinine’s action and changes in the A549 cell line seem justified. Our study is the first to estimate the influence of cotinine on F-actin in A549 cells.
The results obtained in this study indicate that cotinine, at the doses applied, significantly influences NSCLC cells. The morphological changes observed after cotinine treatment may suggest a cellular response to stress conditions. Cells with a phenotype indicative of mitotic catastrophe were also detected. Observations similar to those we obtained, among others, with the use of microtubule stabilization factors [4,6,15,24] suggest that cotinine may act in a similar way. However, to confirm this, further studies using additional research methods are required.
CONCLUSIONS
1. The results allow us to conclude that cotinine significantly influences the structure and number of non-small-cell lung cancer A549 cells.
2. On the basis of changes in the area of the cytoskeleton at the classical fluorescence microscopy level, it is possible to suppose that F-actin causes changes in the shape and size of A549 cells after cotinine treatment. Intensification of these changes was observed with increased cotinine dose.
3. The results on cotinine’s action in the A549 cell line included the presence of cells with apoptotic features as well as cells with a mitotic catastrophe phenotype, which implies that cells of the A549 line may undergo two types of death, i.e. with activation of pathways leading to apoptosis and mitotic catastrophe.
4. Significant changes in the non-small-cell lung cancer cells were observed at the level of electron microscopy. The changes were intensified with an increase in cotinine dose and involved both the nucleus and cytoplasmic organelles. This allows the suggestion that cotinine causes changes not only within the cytoskeleton and nucleus, but also in other cellular organelles.
REFERENCES
[1] Bao Z., He X.Y., Ding X., Prabhu S., Hong J.Y.: Metabolism of nicotine and cotinine by human cytochrome P450 2A13. Drug Metab. Dispos., 2005; 33: 258-261
[PubMed] [Full Text HTML] [Full Text PDF]
[2] Burns D.M.: Cigarettes and cigarette smoking. Clin. Chest Med., 1991; 12: 631-642
[PubMed]
[3] Cook D.G., Whincup P.H., Jarvis M.J., Strachan D.P., Papacosta O., Bryant A.: Passive exposure to tobacco smoke in children aged 5-7 years: individual, family, and community factors. Br. Med. J., 1994; 308: 384-389
[PubMed] [Full Text HTML] [Full Text PDF]
[4] Dasgupta P., Kinkade R., Joshi B., DeCook C., Haura E., Chellapan S.: Nicotine inhibits apoptosis induced by chemotherapeutic drugs by up-regulating XIAP and survivin. Proc. Natl. Acad. Sci. USA, 2006; 103: 6332-6337
[PubMed] [Full Text HTML] [Full Text PDF]
[5] Foster K.A., Oster C.G., Mayer M.M., Avery M.L., Audus K.L.: Characterization of the A549 cell line as a type II pulmonary epithelial cell model for drug metabolism. Exp. Cell Res., 1998; 243: 359-366
[PubMed]
[6] Honore S., Kamath K., Braguer D., Wilson L., Briand C., Jordan M.A.: Suppression of microtubule dynamics by discodermolide by a novel mechanism is associated with mitotic arrest and inhibition of tumor cell proliferation Mol. Cancer Ther., 2003; 2: 1303-1311
[PubMed] [Full Text HTML] [Full Text PDF]
[7] Hoshino Y., Mio T., Nagai S., Miki H., Ito I., Izumi T.: Cytotoxic effects of cigarette smoke extract on an alveolar type II cell-derived cell line. Am. J. Physiol. Lung Cell. Mol. Physiol., 2001; 281: L509-L516
[PubMed] [Full Text HTML] [Full Text PDF]
[8] Hukkanen J., Jacob P.3rd, Benowitz N.L.: Metabolism and disposition kinetics of nicotine. Pharmacol. Rev., 2005; 57: 79-115
[PubMed] [Full Text HTML] [Full Text PDF]
[9] Jin Z., Gao F., Flagg T., Deng X.: Nicotine induces multi-site phosphorylation of Bad in association with suppression of apoptosis. J. Biol. Chem., 2004; 279: 23837-23844
[PubMed] [Full Text HTML] [Full Text PDF]
[10] Katakura Y.: Molecular basis for the cellular senescence program and its application to anticancer therapy. Biosci. Biotechnol. Biochem., 2006; 70: 1076-1081
[PubMed] [Full Text PDF]
[11] Klein L.E., Freeze B.S., Smith A.B.3rd, Horwitz S.B.: The microtubule stabilizing agent discodermolide is a potent inducer of accelerated cell senescence. Cell Cycle, 2005; 4: 501-507
[PubMed] [Full Text HTML] [Full Text PDF]
[12] Kołodziejczyk J.: Wpływ biernego palenia na stan zdrowia osób niepalących. Kosmos, 2002; 51: 47-55
[13] Mai H., May W.S., Gao F., Jin Z., Deng X.: A functional role for nicotine in Bcl2 phosphorylation and suppression of apoptosis. J. Biol. Chem., 2003; 278: 1886-1891
[PubMed] [Full Text HTML] [Full Text PDF]
[14] Maneckjee R., Minna J.D.: Opioid and nicotine receptors affect growth regulation of human lung cancer cell lines. Proc. Natl. Acad. Sci. USA, 1990; 87: 3294-3298
[PubMed] [Full Text HTML] [Full Text PDF]
[15] Masterson J.C., O’Dea S.: 5-Bromo-2-deoxyuridine activates DNA damage signalling responses and induces a senescence-like phenotype in p16-null lung cancer cells. Anticancer Drugs, 2007; 18: 1053-1068
[PubMed]
[16] Nakajima M., Yamagishi S., Yamamoto H., Yamamoto T., Kuroiwa Y., Yokoi T.: Deficient cotinine formation from nicotine is attributed to the whole deletion of the CYP2A6 gene in humans. Clin. Pharmacol. Ther., 2000; 67: 57-69
[PubMed]
[17] Ramage L., Jones A.C., Whelan C.J.: Induction of apoptosis with tobacco smoke and related products in A549 lung epithelial cells in vitro. J. Inflamm., 2006; 3: 3
[PubMed] [Full Text HTML] [Full Text PDF]
[18] Samet J.M., Wiggins C.L., Humble C.G., Pathak D.R.: Cigarette smoking and lung cancer in New Mexico. Am. Rev. Respir. Dis., 1988; 137: 1110-1113
[PubMed]
[19] Smith C.D., Craft D.W., Shiromoto R.S., Yan P.O.: Alternative cell line for virus isolation. J. Clin. Microbiol., 1986; 24: 265-268
[PubMed] [Full Text PDF]
[20] Tai H.H., Tong M., Ding Y.: 15-hydroxyprostaglandin dehydrogenase (15-PGDH) and lung cancer. Prostaglandins Other Lipid Mediat., 2007; 83: 203-208
[PubMed] [Full Text HTML] [Full Text PDF]
[21] Travis W.D., Travis L.B., Devesa S.S.: Lung cancer. Cancer, 1995; 75 (Suppl. 1): 191-202
[PubMed]
[22] Tsuji T., Aoshiba K., Nagai A.: Cigarette smoke induces senescene in alveolar epithelial cells. Am. J. Respir. Cell Mol. Biol., 2004; 31: 643-649
[PubMed] [Full Text HTML] [Full Text PDF]
[23] Tutka P., Mosiewicz J., Wielosz M.: Pharmacokinetics and metabolism of nicotine. Pharmacol. Rep., 2005; 57: 143-153
[PubMed] [Full Text PDF]
[24] Vogel C., Hager C., Bastians H.: Mechanisms of mitotic cell death induced by chemotherapy-mediated G2 checkpoint abrogation. Cancer Res., 2007; 67: 339-345
[PubMed] [Full Text HTML] [Full Text PDF]
[25] Wang X., Yin Y., Sui C., Meng F., Ma P., Jiang Y.: Inhibitory effect of cantharidin on proliferation of A549 cells. Chinese J. Cancer Res., 2007; 19: 283-286
[Abstract]
[26] WHO Cancer Feb 2006 (10.04.2008)
http://www.who.int/mediacentre/factsheets/fs297/en/
[27] WHO The world health report 2004 – changing history (10.04.2008)
http://www.who.int/whr/2004/en/
[28] Wu X.H., Wang H.F., Liu Y.F., Lu X.Y., Wang J.J., Li K.: Histone adduction with nicotine: a bio-AMS study. Radiocarbon, 1997; 39: 293-297
[Abstract] [Full Text PDF]
[29] Yildiz D.: Nicotine, its metabolism and an overview of its biological effects. Toxicon, 2004; 43: 619-632
[PubMed]
The authors have no potential conflicts of interest to declare.