
Złoty środek na otyłość? Udział laktoferryny w metabolizmie glukozy i lipidów
Jolanta Artym 1Streszczenie
Otyłość, dyslipidemia, hiperglikemia/cukrzyca typu 2 i nadciśnienie tętnicze stanowią razem tzw. zespół metaboliczny. Częstość występowania tych poważnych zaburzeń metabolicznych wiąże się ze stylem życia i szybko wzrasta w bogatych krajach uprzemysłowionych. Schorzenia te stanowią nie tylko poważny problem zdrowotny, ale też społeczny i ekonomiczny, angażując w profilaktykę i leczenie rzesze praktyków różnych specjalności (lekarzy, dietetyków, psychologów). Od kilkunastu lat trwają badania nad możliwością użycia naturalnych białek pochodzących z mleka w zapobieganiu i leczeniu wspomnianych chorób metabolicznych. Jednym z intensywniej badanych białek jest laktoferryna (LF), białko obecne w mleku i wydzielinach komórkowych ssaków. Pierwsze testy wykazujące korzystny wpływ LF na metabolizm lipidów oraz otyłość przeprowadzono w Japonii w 2003 r. na zaledwie kilku ochotnikach. Kolejne próby to zarówno badania na zwierzętach, jak i randomizowane próby kliniczne, których wyniki są poparte odpowiednimi testami in vitro. Po doustnym podaniu LF obserwowano m.in.: spadek masy ciała, zmniejszenie obwodu pasa i ilości trzewnej tkanki tłuszczowej, obniżenie osoczowych i wątrobowych stężeń kwasów tłuszczowych, triglicerydów oraz cholesterolu. Mechanizm działania LF może obejmować kilka aspektów: hamowanie procesów adipogenezy, ograniczenie absorpcji triglicerydów pokarmowych, zwiększenie poziomu antyaterogennej frakcji cholesterolu HDL oraz hamowanie akumulacji utlenionej postaci cholesterolu LDL w makrofagach i ochronę przed tworzeniem „komórek piankowatych”. Laktoferryna ponadto zwiększa wrażliwość komórek na działanie insuliny, również w warunkach, gdy odpowiedź na insulinę jest obniżona (w stanie zapalnym). Dodatkowo LF reguluje aktywność białek systemu IGF (insulin-like growth factor). Uzyskane dotychczas dane wskazują, że LF jest obiecującym, naturalnym, całkowicie nietoksycznym środkiem, który (np. jako suplement diety) może być stosowany w długoterminowej profilaktyce oraz terapii zaburzeń metabolicznych, takich jak: dyslipidemia, otyłość, insulinooporność/cukrzyca typu 2.
Słowa kluczowe:laktoferryna • zespół metaboliczny • otyłość • dyslipidemia • insulinooporność • hiperglikemia • cukrzyca typu 2 • nadciśnienie tętnicze • cholesterol • triglicerydy • stan zapalny
Summary
Obesity, dyslipidemia, hyperglycemia/type II diabetes and hypertension together constitute the so-called metabolic syndrome. Frequency of occurrence of these serious metabolic disturbances is associated with life style and is on the rise in prosperous industrialized countries. These diseases represent not only a serious health problem but also social and economic ones, and involve in prophylaxis and treatment various specialists (physicians, dieticians and psychologists). For about two decades research has been conducted on the possibility to apply milk-derived proteins in prevention and treatment of the above mentioned metabolic diseases. Lactoferrin (LF), a protein present in milk and excretory fluids of mammals, is one of the most intensively studied milk proteins for therapeutic application. Initial trials revealing an advantageous effect of LF on lipid metabolism and obesity enrolled only a few volunteers and were performed in Japan in 2003. Subsequent trials were conducted on animals as well as in clinics, and the positive results were supported by in vitro tests. After oral administration of LF, decreases of body weight, waist measurement, visceral fat tissue, plasma and liver fatty acid concentrations, triglycerides and cholesterol were registered. The mechanism of LF action may involve several processes, such as inhibition of adipogenesis, decrease of dietary triglyceride absorption, elevation of HDL cholesterol possessing anti-atherogenic properties, inhibition of accumulation of oxidized LDL cholesterol forms in macrophages and protection against formation of foam cells. LF also increases the susceptibility of cells to insulin action, including in conditions when the response to insulin is lowered (during inflammation). In addition, LF regulates activity of insulin-like growth factor (IGF). The data collected to date indicate that LF is a promising, completely nontoxic, natural remedy which (as for example a food supplement) may be applied in long-term prophylaxis and therapy of metabolic disturbances, such as dyslipidemia, obesity and insulin resistance/type II diabetes.
Key words:lactoferrin • metabolic syndrome • obesity • dyslipidemia • insulin resistance • hyperglycemia • type II diabetes • hypertension • cholesterol • triglycerides • inflammation
Wykaz skrótów:
ac-LDL – acetylowane (acetylated) LDL; AGE – produkty glikacji białek (advanced glycation end products); Ala – alanina; apo-E – apolipoproteina E; apo-LF – laktoferryna wolna od żelaza; Arg – arginina; BLF – laktoferryna bydlęca; BMI – wskaźnik masy ciała (body mass index); CRP – białko C-reaktywne (C-reactive protein); HDL – lipoproteiny o dużej gęstości (high density lipoproteins); HLF – laktoferryna ludzka; holo-LF – laktoferryna wysycona żelazem; IDL – lipoproteiny o pośredniej gęstości (intermediate density lipoproteins); IL – interleukina; i.v. – dożylnie (intravenously); LDL – lipoproteiny o małej gęstości (low density lipoproteins); LF – laktoferryna; LRP1 – białko związane z receptorem LDL (LDL receptor related protein 1); LPS – lipopolisacharyd, endotoksyna (lipopolysaccharide, endotoxin); Lys – lizyna; MCP-1 – białko chemotaktyczne dla monocytów (monocyte chemoattractant protein-1); ox-LDL – utlenione (oxidized) LDL; PAI-1 – inhibitor aktywatora plazminogenu 1 (plasminogen activator inhibitor 1); p.o. – doustnie (per os); RFT – reaktywne formy tlenu; rHLF – rekombinacyjna laktoferryna ludzka; s.c. – podskórnie (subcutaneously); SNP – polimorfizm typu pojedynczego nukleotydu (single-nucleotide polymorphism); TG – triglicerydy (trójglicerydy); Thr – treonina; VLDL – lipoproteiny o bardzo małej gęstości (very low density lipoproteins); WHR – współczynnik talia-biodro (waist-hip ratio).
Wstęp
Zaburzenia gospodarki lipidowej (dyslipidemia), otyłość, cukrzyca typu 2 oraz nadciśnienie to poważne problemy zdrowotne w uprzemysłowionych krajach Zachodu. O skali problemu świadczą liczby: otyłość dotyka aż 2 mld ludzi na świecie, około 260 mln cierpi natomiast na cukrzycę typu 2 (w Unii Europejskiej to 28 mln, a w Polsce – 1,8 mln osób) [56]. Główne przyczyny wymienionych zaburzeń to: złe odżywianie, brak wysiłku fizycznego, stres psychiczny, palenie tytoniu oraz starzenie się społeczeństw, a skutki to m.in.: miażdżyca, zawał mięśnia sercowego, udar mózgu, angiopatie. Schorzenia te stanowią poważny problem zdrowotny, społeczny i ekonomiczny, angażując w profilaktykę i leczenie praktyków różnych specjalności (lekarzy, dietetyków, psychologów). Bezsprzeczna zależność zaburzeń metabolicznych od ogólnie pojętego „stylu życia”, umożliwia profilaktykę i terapię tych schorzeń przez jego modyfikację, która powinna objąć długotrwałą zmianę zwyczajów żywieniowych, zwiększenie aktywności fizycznej, zaprzestanie palenia i unikanie długotrwałego napięcia psychicznego.
W wielu ośrodkach światowych trwają badania nad możliwością użycia naturalnych związków (w tym pochodzących z mleka) w zapobieganiu i leczeniu wspomnianych chorób metabolicznych. Ich zastosowanie byłoby szczególnie korzystne, gdyż włączone do codziennej diety i długotrwale zażywane nie dawałoby działań niepożądanych. Już na początku lat 90. ubiegłego wieku zwrócono uwagę na białka mleka krowiego, które jak się okazało, skutecznie obniżały osoczowe poziomy lipidów [77]. To zasugerowało zbadanie poszczególnych białek mleka odpowiedzialnych za ten efekt.
Jednym z białek intensywniej badanych pod tym kątem jest laktoferryna (LF), obecna w wydzielinach komórkowych oraz granulocytach ssaków. LF wydzielnicza, którą możemy znaleźć w siarze, mleku, łzach, ślinie, wydzielinie dróg oddechowych, dróg rodnych, soku żołądkowym i innych chroni nasz organizm przed atakiem drobnoustrojów pochodzących ze środowiska zewnętrznego. Białko obecne w neutrofilach może być uwalniane podczas degranulacji tych komórek, co zwiększa jego stężenie w tkankach i krążeniu w czasie infekcji, zapalenia i urazów [34]. LF jest istotnym składnikiem odporności wrodzonej organizmu. Wykazuje aktywność wobec różnorodnych bakterii Gram-ujemnych i -dodatnich, wirusów otoczkowych i bezotoczkowych oraz różnych rodzajów grzybów i pasożytów [27]. Uczestniczy ponadto w metabolizmie żelaza [1], regulacji hematopoezy [2], wykazuje działanie chemioprewencyjne i przeciwnowotworowe [18] oraz immunoregulatorowe, hamując bądź stymulując odpowiedź odpornościową ustroju [35]. Jak dotąd laktoferrynę przebadano w wielu testach laboratoryjnych, przedklinicznych i klinicznych. Uzyskane wyniki wskazują na jej przydatność w profilaktyce i terapii chorób autoimmunizacyjnych i neoplastycznych, alergii, zakażeń, sepsy, bakteriemii i endotoksemii, ran, osteoporozy, anemii z niedoboru żelaza oraz odnowie funkcji układu immunologicznego po chemioterapii [72].
Dzięki licznym badaniom przeprowadzonym w ciągu ostatnich kilkunastu lat, wspomniane wyżej plejotropowe działanie LF możemy obecnie uzupełnić o korzystny wpływ na gospodarkę lipidową i energetyczną ustroju. W pracy omówiono dane historyczne oraz wyniki najnowszych badań w tym zakresie.
Zespół metaboliczny: dyslipidemia, otyłość, cukrzyca typu 2, nadciśnienie tętnicze
Mianem hiperlipidemii określamy podwyższone stężenia cholesterolu i/lub triacylogliceroli (triglicerydów, TG) powyżej wartości uznawanych za prawidłowe, czyli odpowiednio: 5,2 mmol/l (200 mg/dl) i 2,0 mmol/l (180 mg/dl). Dyslipidemia (dyslipoproteinemia) to pojęcie szersze, podkreślające kliniczne znaczenie małych stężeń HDL (high density lipoproteins) często towarzyszące podwyższonym poziomom innych lipoprotein. Podstawową metodą profilaktyki i leczenia tych schorzeń jest odpowiednia dieta, która pozwala uzyskać poprawę lub normalizację wskaźników metabolicznych u dużej części osób. Głównym założeniem diety w hiperlipidemii jest ograniczenie ogólnej podaży tłuszczów (szczególnie tłuszczów nasyconych), a zwiększenie spożycia tłuszczów wielonienasyconych [63].
Otyłość powstaje wskutek utrzymującego się przez dłuższy czas dodatniego bilansu energetycznego (nadmierna podaż energii wobec zbyt małego zużycia) i charakteryzuje się nadmiernym zwiększeniem masy ciała. Prawidłowość masy określa się najczęściej według tzw. wskaźnika BMI (body mass-index), który powinien mieścić się w zakresie 20-25 kg/m2 (gdy mieści się w zakresie 25-30 kg/ m2 mówimy o nadwadze, gdy jest wyższy niż 30 oraz 40 kg/m2 oznacza odpowiednio otyłość i otyłość chorobliwą). W przebiegu otyłości dochodzi do zwiększenia objętości i masy tkanki tłuszczowej, w wyniku zarówno hipertrofii (powiększenia rozmiarów), jak i hiperplazji (zwiększenia liczby na skutek podziałów) komórek tłuszczowych (adipocytów). Adipocyty powstają z fibroblastów podczas prawidłowego rozwoju i w stanach patologicznych. Stanowią miejsce magazynowania i uwalniania kwasów tłuszczowych, służących za źródło energii. Poprzez wydzielane cząsteczki regulatorowe (tzw. adipokiny, adipocytokiny) regulują ponadto wiele procesów metabolicznych, m.in. przemiany glukozy i lipidów, równowagę energetyczną, apetyt, odporność, ciśnienie krwi czy angiogenezę [25].
Właściwości fizjologiczne adipocytów zgromadzonych w różnych częściach ciała różnią się, czym można tłumaczyć odrębności kliniczne różnych typów otyłości. Co istotne, następstwa zdrowotne otyłości brzusznej (wisceralnej, centralnej, androidalnej), w której tkanka tłuszczowa gromadzi się w jamie brzusznej (w sieci większej, krezce jelita, torebkach nerek, pod otrzewną i w przestrzeni pozaotrzewnowej), są znacznie poważniejsze niż otyłości pośladkowo-udowej (obwodowej; akumulacja tłuszczu w tkance podskórnej, głównie okolicy bioder i pośladków). Jest to zespół skomplikowanych zaburzeń metabolicznych, cechujący się m.in. dyslipidemią, hiperinsulinemią i nadciśnieniem. Te dwa rodzaje tkanki tłuszczowej różnią się udziałem w metabolizmie glukozy i lipidów. Tkanka tłuszczowa podskórna gromadzi zapasy energii w adipocytach, w postaci triglicerydów, w sytuacji nadmiernego dowozu energii (dieta wysokokaloryczna) i przy jej zbyt małym zużyciu (brak aktywności fizycznej). Działa więc jako naturalny, metaboliczny bufor lub „metaboliczny zlew” dla nadmiaru tłuszczu. Adipocyty podskórne zachłannie wychwytują wolne kwasy tłuszczowe i TG w czasie hiperlipidemii poposiłkowej. Gdy zdolność magazynowania tłuszczu w tej tkance się wyczerpie lub zdolność do tworzenia nowych adipocytów jest ograniczona z powodu predyspozycji genetycznych lub chronicznego stresu (podniesiony poziom kortyzolu) dochodzi do odkładania się zapasów tłuszczu poza jego naturalnym magazynem, w obrębie jamy brzusznej, a także w wątrobie i tkance mięśniowej (tzw. ektopowe depozyty tłuszczu) [25]. Mniejsze adipocyty podskórne (tzw. tkanka tłuszczowa żółta) charakteryzują się większą wrażliwością na działanie insuliny i większą aktywnością lipogenną. Tymczasem większe komórki z tkanki tłuszczowej brzusznej są bardziej aktywne metabolicznie i bardziej „skłonne” do lipolizy oraz wydzielania adipokin „atakujących” i prozapalnych: IL-6, TNF-α, białka C-reaktywnego (C-reactive protein, CRP), angiotensynogenu, inhibitora aktywatora plazminogenu 1 (plasminogen activator inhibitor 1, PAI-1), adiponektyny, leptyny i rezystyny. Czynniki te zmniejszają wrażliwość tkanek na insulinę oraz mogą stymulować proces zapalny, pogorszenie funkcji śródbłonka naczyniowego, tworzenie zakrzepów w naczyniach i sprzyjać rozwojowi miażdżycy [30,62]. Udział tkanki tłuszczowej podskórnej i brzusznej w metabolizmie przedstawiono na ryc. 1. Istnieją obserwacje sugerujące, że jednym z czynników wiążących nadmierny pobór kalorii i dodatni bilans energetyczny z opornością na insulinę i cukrzycą typu 2 jest nieodpowiedni rozwój podskórnej tkanki tłuszczowej, z jednoczesnym tworzeniem ektopowych depozytów tłuszczu w wątrobie i mięśniach. Ważną rolę regulacyjną w tych procesach przypisuje się adiponektynie [32], o której jeszcze będzie mowa. Akumulację tłuszczu w tkance podskórnej stymulują estrogeny, co ma znaczenie protekcyjne. Niedobór tych hormonów u kobiet w okresie menopauzalnym przyczynia się do gromadzenia zapasów tłuszczu brzusznego, zwiększając zagrożenie dyslipidemią i cukrzycą typu 2 [25].

Ryc. 1. Nadmierna podaż energii w posiłkach prowadzi do akumulacji TG w adipocytach tkanki podskórnej, która działa jako „metaboliczny zlew”. Gdy zdolność tej tkanki do magazynowania tłuszczu zostanie przekroczona lub jest nieprawidłowa (np. wskutek predyspozycji genetycznych lub adaptacji do długotrwałego stresu) tłuszcz gromadzi się poza tkanką podskórną, w postaci tłuszczu brzusznego lub ektopowych depozytów w wątrobie i mięśniach, co poprzez wydzielane adipokiny predysponuje do zaburzeń przemiany lipidów i glukozy oraz rozwoju miażdżycy (na podstawie [25], zmodyfikowano)
W profilaktyce i leczeniu otyłości najważniejsze jest ograniczenie kaloryczności posiłków, z jednoczesnym ich zbilansowaniem, by uniknąć niedoborów ważnych składników odżywczych, a także wzmożony wysiłek fizyczny (czyli zwiększone wydatkowanie energii tak, by doszło do deficytu energetycznego).
W cukrzycy typu 2 (insulinoniezależnej) może dochodzić zarówno do zmniejszenia ilości wydzielanej insuliny endogennej, jak i/lub zmniejszenia liczby i wrażliwości receptorów insulinowych na komórkach (zjawisko oporności). Oznacza to, że komórki docelowe nie reagują właściwie na prawidłowe, a nawet zwiększone ilości insuliny krążącej we krwi. Nie otrzymują niezbędnej porcji energii w postaci glukozy, a cukier pozostający przez długi czas w krążeniu (hiperglikemia) działa toksycznie na różne tkanki. Oporność na insulinę wzrasta wraz z przyrostem masy ciała – około 80% osób z tym typem cukrzycy to osoby otyłe. Tkanka tłuszczowa, jak już wspomniano, ma znaczenie w rozwoju oporności na insulinę, jest bowiem nie tylko magazynem tłuszczu, ale też narządem endokrynnym, wydzielającym liczne adipokiny [30,62]. W cukrzycy dochodzi zarówno do upośledzenia syntezy kwasów tłuszczowych, jak i ich nadmiernego spalania. Zahamowanie lipogenezy, a wzmożenie lipolizy prowadzi do zwiększenia ilości wolnych kwasów tłuszczowych we krwi. Ponadto obserwuje się wzrost stężeń triglicerydów, fosfolipidów i cholesterolu. Hiperlipidemia cukrzycowa może przyspieszać proces aterogenezy (powstawania miażdżycy). Leczenie tego typu cukrzycy obejmuje: dietę, zastosowanie doustnych leków hipoglikemizujących (zwiększających wydzielanie insuliny endogennej oraz wychwytywanie glukozy przez tkanki obwodowe) lub insuliny, w razie potrzeby, oraz leczenie wysiłkiem fizycznym (poprzez trening leczniczy).
Z opornością na insulinę wiąże się nadciśnienie tętnicze krwi, co nie dziwi, gdy uwzględnimy wazodylatacyjne (rozszerzające naczynia krwionośne) działanie insuliny, regulację reabsorpcji sodu w nerkach oraz wpływ na aktywność sympatycznego układu nerwowego [12,16]. Do wzrostu ciśnienia krwi przyczyniają się również kwasy tłuszczowe, które obkurczają naczynia krwionośne [69].
Od ponad 20 lat trwają badania, które pozwoliły powiązać ze sobą omawiane schorzenia (otyłość, dyslipidemię, insulinooporność/cukrzycę typu 2, nadciśnienie) w jeden zespół chorobowy, określany jako „zespół metaboliczny” (metabolic syndrome), a dawniej jako „zespół X”. Wyodrębnienie tego zespołu ma głównie znaczenie praktyczne, gdyż może pomóc w sprawnej diagnostyce osób zagrożonych wspomnianymi schorzeniami. Przez niektórych badaczy i praktyków pojęcie zespołu metabolicznego jest jednak krytykowane, m.in. ze względu na brak opracowania jednoznacznych wskaźników diagnostycznych [10,61]. Obecnie szacuje się, że zespół metaboliczny dotyczy około 25-30% populacji światowej.
Centralnym zaburzeniem metabolizmu stwierdzanym u wszystkich pacjentów jest oporność komórek na insulinę połączona ze słabym, ale przewlekłym stanem zapalnym [9,10]. Wykazano bezpośrednią korelację między krążącymi cytokinami zapalnymi (TNF-α, IL-6) a insulinoopornością, ale przyczyna chronicznego zapalenia nie została, jak dotąd, dokładnie poznana. Uważa się, że dochodzi do stałej aktywacji układu immunologicznego przez różne czynniki zewnętrzne, takie jak: bogatotłuszczowa dieta, molekularne wzorce związane z patogenami (pathogen associated molecular patterns, PAMP), np. lipopolisacharyd (LPS, endotoksyna), produkty glikacji białek (advanced glycation end products; AGE), infekcje, stres psychiczny, stres oksydacyjny [44]. Być może odpowiedź rozpoczyna się od rekrutacji neutrofilów do trzewnej tkanki tłuszczowej, do której dochodzi po posiłkach bogatotłuszczowych [13]. Znaczenie może mieć też tzw. metaboliczna endotoksemia, charakteryzująca się wzrostem osoczowych stężeń endotoksyny 2-3-krotnie ponad stan fizjologiczny podczas długotrwałej diety bogatej w tłuszcze [6]. U myszy dieta taka zwiększała udział bakterii Gram-ujemnych wśród mikroflory jelitowej, co może się przyczyniać do obserwowanej endotoksemii na skutek przenikania bakterii przez barierę jelitową. Długotrwała infuzja s.c. małych dawek LPS powodowała podobne skutki: insulinemię, glikemię, oporność na insulinę, przyrost tkanki tłuszczowej oraz wzrost poziomu markerów zapalenia [6].
W zespole metabolicznym stwierdza się zaburzenia funkcji komórek immunologicznych, głównie granulocytów obojętnochłonnych, z czym można wiązać dużo większą podatność pacjentów z cukrzycą na zakażenia. Ta zależność nie dziwi, gdy weźmiemy pod uwagę, że insulina działa regulująco na komórki odpornościowe [73]. Charakterystyczne jest, że wraz z opornością na insulinę zmienia się wytwarzanie białek odpornościowych w granulocytach. Zaburzenia te dotyczą: LF, białka bakteriobójczego zwiększającego przepuszczalność (bactericidal/increasing permeability protein, BPI), rozpuszczalnej postaci receptora CD14 (soluble, sCD14), a-defensyn i niektórych składników płucnego surfaktantu (surfactant protein D, SP-D) [44]. Obniżone stężenia tych białek w krążeniu korelują z obniżoną odpornością pacjentów i stanowią swoiste markery cukrzycy typu 2. Wydaje się, że w celu kompensacji organizm zwiększa wytwarzanie innych przeciwmikrobiologicznych białek, takich jak: LBP (LPS-binding protein), NGAL (neutrophil gelatinase-associated lipocalin) oraz SP-A (surfactant protein A). Nieprawidłowo działające składniki odporności wrodzonej nie buforują skutecznie ataku na organizm różnych czynników zewnętrznych (LPS, AGE, RFT i innych), a to nie pozwala na wygaszenie stanu zapalnego. Ten z kolei zmienia metabolizm, prowadząc do rozwoju oporności na insulinę/cukrzycy typu 2, otyłości, dyslipidemii, dysfunkcji wątroby. Te zaburzenia ponownie nasilają opisane zmiany w układzie odpornościowym. Dochodzi zatem do rozwoju „błędnego koła”, gdzie niekorzystne procesy w układzie odpornościowym i metabolizmie wzajemnie się nasilają (ryc. 2).

Ryc. 2. Zaburzenia metaboliczne prowadzące do rozwoju zespołu metabolicznego z główną rolą przewlekłego stanu zapalnego i insulinooporności. Najnowsze badania łączą rozwój zespołu metabolicznego z przewlekłym stanem zapalnym. Przyczyna słabego chronicznego stanu zapalnego nie jest znana, ale prawdopodobnie układ immunologiczny jest pobudzany przez różne czynniki zewnętrzne (bogatotłuszczowa dieta, endotoksyna i inne PAMP, AGE, infekcje, stres psychiczny, stres oksydacyjny). Szczególne znaczenie należy przypisać zbyt kalorycznej i bogatej w tłuszcze diecie. Wzrasta wytwarzanie czynników prozapalnych (IL-6, TNF-α, CRP, PAI-1), które uszkadzają komórki ß trzustki oraz interferują z aktywnością insuliny, zmniejszając wrażliwość komórek na jej działanie, co z kolei hamuje przeciwzapalne działanie insuliny. Prowadzi to do dalszego nasilenia stanu zapalnego, rozwoju insulinooporności i w konsekwencji cukrzycy typu 2, co z kolei nasila zaburzenia lipidowe. Rozwija się zespół metaboliczny, na który składają się: otyłość, zaburzenia lipidowe, cukrzyca typu 2 i nadciśnienie [6,9,13,44]
Regulacyjne działanie LF obserwowane w testach przedklinicznych i klinicznych
Początkowo uwagę na możliwy udział LF w gospodarce lipidowej organizmu zwrócili uczeni japońscy, przeprowadzając testy przedkliniczne oraz wstępne małe próby kliniczne. Pierwsze badanie objęło zaledwie kilkuosobową grupę zdrowych ochotników. Bydlęca LF podana w postaci kapsułek powleczonych warstwą chroniącą przed działaniem soku żołądkowego obniżała stężenie cholesterolu i triglicerydów we krwi [33,59]. LF podano osobom (n=4) po incydencie spożycia nadmiernej ilości alkoholu, który spowodował znaczny (do ponad 500 mg/dl) wzrost stężenia triglicerydów we krwi, co jest jednym z toksycznych skutków spożycia etanolu. Do wzrostu stężenia TG dochodzi wskutek zwiększonej syntezy oraz zahamowania β-oksydacji (utleniania) kwasów tłuszczowych, co powoduje ich akumulację, a w konsekwencji zwiększoną syntezę TG. U osób długotrwale spożywających alkohol TG gromadzą się w hepatocytach, wywołując stłuszczenie wątroby. LF w ilości 450 mg/dzień (w trzech dawkach podzielonych) obniżyła poziom triglicerydów do stężeń prawidłowych w ciągu 1-2 dni, podczas gdy u osób z grupy kontrolnej nie powrócił do normy w ciągu kolejnych 7 dni [33]. W drugim badaniu LF podawana w dawce 450 mg/dzień przez 4 tygodnie obniżyła poziom cholesterolu u 6 spośród 8 testowanych ochotników. W kilku przypadkach spadek był znaczny (np. z wartości 280 na 210 mg/dl). U dwu osób z wyjściowo niskim stężeniem cholesterolu (170 mg/dl) LF nie dała żadnych zmian [33,59]. Mechanizm obserwowanego działania białka nie jest znany, ale jak sugerują autorzy, może się wiązać z przyspieszeniem procesów podstawowego metabolizmu i spalania tłuszczów, a tym samym redukcji zapasów tłuszczu w organizmie. Potwierdzeniem tych przypuszczeń jest podniesienie podstawowej temperatury ciała, zwłaszcza po posiłku oraz spadek masy ciała i obwodu pasa u osób przyjmujących LF. W ciągu 1-2 miesięcy u części osób stwierdzono redukcję masy o 2-6 kg i obwodu pasa o 2-3 cm (bez zastosowania specjalnej diety i ćwiczeń fizycznych). Działanie LF może również obejmować przerwanie wątrobowo-jelitowego krążenia kwasów żółciowych, o czym jeszcze będzie mowa.
Skuteczność LF potwierdziły następnie testy na zwierzętach. BLF zastosowana jako dodatek do diety (w ilości 10 g/kg paszy) w ciągu 4 tygodni zredukowała poziom osoczowego i wątrobowego cholesterolu i TG u myszy [66]. Jednocześnie wzrósł poziom tzw. „dobrego cholesterolu” (związanego z frakcją HDL). Karmienie LF obniżało absorpcję triglicerydów z diety, ale tylko nieznacznie zwiększało wydalanie kwaśnych steroli z kałem, być może w wyniku zmniejszenia ponownego wyłapywania kwasów żółciowych w jelicie i powrotu do wątroby. Doustne podawanie LF (10 mg/dzień) myszom z genetycznie uwarunkowaną otyłością chroniło przed wzrostem masy ciała oraz obniżało poziom cholesterolu w krążeniu [51]. U szczurów, którym podawano paszę z dodatkiem 0,2% BLF przez 10 miesięcy obserwowano obniżenie osoczowych poziomów TG o ponad 30% w porównaniu ze zwierzętami kontrolnymi [67]. Skuteczny okazał się również dodatek LF do diety u myszy z otyłością wywołaną wcześniejszym karmieniem dietą bogatotłuszczową [55]. LF przyspieszała spadek masy ciała, redukowała ciężar oraz zawartość % tkanki tłuszczowej, jak również wielkość komórek tłuszczowych. U myszy otyłych obserwowano ponadto podwyższone stężenie glukozy we krwi, które było istotnie obniżane przez LF. Myszy karmione LF przyswajały mniejsze ilości tłuszczu z pokarmu, czym można być może, tłumaczyć działanie białka. W różnym stopniu skuteczne były też inne białka serwatki mleka (α-laktoalbumina i β-laktoglobulina) oraz mieszanina różnych białek serwatkowych, ale nie kazeina.
Znacznie nowsze badanie kliniczne, również w Japonii, objęło grupę 26 osób cierpiących na otyłość typu brzusznego (BMI >25 kg/m2; obszar tłuszczu trzewnego (VFA) > 100 cm2) [54]. Jak już wspomniano, ten typ otyłości jest szczególnie groźny, gdyż towarzyszy mu zespół metaboliczny z dyslipidemią, hiperinsulinemią, obniżoną tolerancją glukozy, nadciśnieniem, podwyższonym stężeniem kwasu moczowego i fibrynogenu. Chorzy ci są zagrożeni wczesnym rozwojem cukrzycy i miażdżycy. Grupie badanej przez 8 tygodni podawano BLF (300 mg/dzień) w postaci tabletek z osłonką chroniącą przed trawieniem w żołądku, grupa kontrolna otrzymywała placebo. Zanotowano istotny spadek: masy ciała (średnio o 1,5 kg), wskaźnika BMI (o 0,6 kg/m2), obwodu bioder (o 2,6 cm), obwodu pasa (o 4,4 cm) oraz VFA (o 14,6 cm2). Autorzy badań sugerują, że jedynie LF podana per os w postaci chronionej przed działaniem soków żołądkowych może być skuteczna, co potwierdzają w późniejszych testach in vitro [53]. BLF poddana trawieniu przez żołądkową pepsynę nie hamowała akumulacji lipidów w szczurzych preadipocytach z krezkowej i podskórnej tkanki tłuszczowej. LF trawiona jelitową trypsyną była aktywna, podobnie jak natywne białko. Uzyskane wyniki wskazują, że LF, aby skutecznie hamować akumulację lipidów w tkance tłuszczowej, nie może być wcześniej zdegradowana przez soki żołądkowe.
Ciekawych wyników dostarczyła niewielka (n=36), randomizowana próba kliniczna przeprowadzona przez badaczy koreańskich na młodych osobach cierpiących z powodu trądziku zwykłego (łojotokowego, acne vulgaris) [31]. W tym schorzeniu dochodzi do nadmiernego wydzielania łoju (sebum) przez gruczoły łojowe na skutek działania hormonów androgenowych, nadmiernego rogowacenia ujścia gruczołu łojowego oraz wtórnego zapalenia mieszka łojowo-włosowego. W patogenezie zmian zapalnych dużą rolę odgrywają Propionobacterium acnes – bakterie beztlenowe żyjące w głębszych strukturach gruczołów łojowych, które wydzielają liczne substancje, w tym enzymy i czynniki chemotaktyczne, przyciągające komórki odpornościowe. Czynnikiem dodatkowo nasilającym stan zapalny jest łój, a szczególnie wolne kwasy tłuszczowe, uwolnione z niego pod wpływem działania bakteryjnych lipaz. Zatem redukcja łojotoku jest jednym z głównych celów leczenia trądziku. We wspomnianym teście, dwie grupy osób (n=18+18) przyjmowały odpowiednio mleko fermentowane zawierające bakterie probiotyczne (Lactobacillus bulgaricus i Streptococcus thermophilus) wzbogacone lub nie w bydlęcą laktoferrynę (200 mg). Po upływie 4, 8 i 12 tygodni oceniano skuteczność terapii. W obu grupach obserwowano wyraźną poprawę: mniejszą liczbę i nasilenie zmian trądzikowych, w tym zmian zapalnych. Korzystniejsze okazało się użycie mleka z probiotykami i LF. Co ważne, obserwowano zmniejszone wydzielanie sebum, spadek ogólnej ilości lipidów, a szczególnie TG i wolnych kwasów tłuszczowych, co korelowało ze złagodzeniem objawów trądziku. Nie obserwowano żadnych działań niepożądanych, w tym zmiany pH i stopnia nawilżenia skóry [31]. Skuteczność LF w leczeniu trądziku pospolitego potwierdziła także najnowsza próba kliniczna (n=39), w której młodym osobom podawano BLF (100 mg/dzień) w postaci tabletek do ssania. Stwierdzono redukcję ogólnej liczby zmian i zmian zapalnych, nie badano jednak innych parametrów, w tym dotyczących gospodarki lipidowej skóry [50].
Istnieją obecnie dwa doniesienia na temat hipotensyjnego działania LF. Białko bydlęce podane dożylnie szczurom obniżało (o 35%) ciśnienie tętnicze krwi, ale nie wpływało na częstość akcji serca [21]. Działanie to było zależne od komórek śródbłonka naczyń i mediowane prawdopodobnie przez wytwarzany przez nie tlenek azotu (NO•) – znany czynnik relaksacyjny wywołujący wazodylatację. Hipotensyjne działanie LF obejmowało raczej centralny, a nie obwodowy układ opioidergiczny, gdyż było znoszone przez nalokson (antagonistę receptorów μ-opioidowych), który przechodzi przez barierę krew-mózg. W teście in vitro zanotowano, zależne od stężenia, relaksacyjne działanie LF na wyizolowaną aortę piersiową. Również peptydy pochodzące z laktoferrycyny B (N-końcowego fragmentu cząsteczki BLF) podane p.o. szczurom obniżały ciśnienie krwi. W teście in vitro obserwowano hamowanie kurczenia naczyń krwionośnych oraz hamowanie aktywności enzymu konwertującego angiotensynę I do angiotensyny II [57]. Enzym ten to najsilniejszy znany związek hipertensyjny, który zwiększa ciśnienie krwi, kurcząc naczynia krwionośne.
Regulacyjne działanie LF w obserwacjach klinicznych
Niezwykle ciekawe obserwacje dotyczące potencjalnego udziału endogennej LF w metabolizmie lipidów i glukozy u ludzi poczynili badacze z Hiszpanii. W próbie liczącej 229 mężczyzn rasy kaukaskiej (94 osoby z prawidłową i 135 osób z obniżoną tolerancją glukozy) badali związek między poziomem krążącej LF a chronicznym zapaleniem i parametrami metabolicznymi [46]. Poziom LF był niższy u pacjentów z obniżoną wrażliwością na insulinę/cukrzycą typu 2. Był odwrotnie skorelowany z BMI, współczynnikiem talia-biodro (waist-hip ratio, WHR), stężeniem TG oraz glukozy na czczo. Dodatnio natomiast korelował z poziomem HDL w osoczu, wrażliwością komórek na insulinę oraz funkcją śródbłonka naczyniowego u pacjentów otyłych z nietolerancją glukozy. Co ważne, poziom LF korelował ujemnie z markerami zapalenia i metabolicznej endotoksemii (np. z LBP, białkiem wiążącym LPS). W hodowlach pełnej krwi wydzielanie LF znacznie spadało pod wpływem stymulacji prozapalną IL-6, a wzrastało po zwiększeniu wrażliwości komórek na insulinę (zastosowanie roziglitazonu). W testach in vitro na komórkach monocytarnych linii THP-1 indukowanych endotoksyną, ludzka LF obniżała ekspresję IL-6, IL-8 oraz białka chemotaktycznego dla monocytów (MCP-1, monocyte chemoattractant protein-1) przez hamowanie aktywności czynnika transkrypcyjnego NF-κB. Jak sugerują autorzy badania, obniżony poziom krążącej LF może odgrywać rolę w chronicznym zapaleniu u osób z nietolerancją glukozy, z kolei stan zapalny obniża poziom LF [46].
Kolejna obserwacja objęła 390 mężczyzn, w tym 188 zdrowych i 202 z nietolerancją glukozy [45]. Stwierdzono występowanie dwóch niezależnych niesynonimicznych polimorfizmów typu pojedynczego nukleotydu (single-nucleotide polymorphism, SNP) w genie LF, które powodowały zmianę dwu reszt aminokwasowych w N-końcowym regionie cząsteczki i były związane z większym ryzykiem dyslipidemii u pacjentów z nietolerancją glukozy. Pierwszy SNP polegał na zamianie reszty Ala na resztę Thr w pozycji 11 dojrzałego białka (rs – reference sequence 1126477), czyli pozycji 29 niedojrzałego łańcucha białkowego. Drugi natomiast „zamieniał” resztę Lys na Arg w pozycji 29 dojrzałego białka (rs1126478), czyli pozycji 47 niedojrzałego łańcucha białkowego. Autorzy sugerują, że powstałe niewielkie modyfikacje cząsteczki białka mogą wpływać na jego interakcje z receptorami LRP1 lub cząsteczkami utlenionych lipoprotein o małej gęstości (ox-LDL), zmieniając w ten sposób stężenia krążących lipidów i tempo ich usuwania z krążenia. Być może obecność reszt Thr i Arg w pozycjach odpowiednio 11 i 29 białka faworyzuje jego silniejsze wiązanie do receptorów LRP1 na komórkach, co może blokować m.in. wychwytywanie cząsteczek HDL przez adipocyty lub wiązanie ox-LDL do receptorów zmiataczowych na makrofagach. Warto zauważyć, że SNP w pozycji 29 genu HLF powiązano również z większą podatnością na zapalenie ozębnej [71,75]. Dodatnio naładowany fragment z końca N cząsteczki LF decyduje również o właściwościach przeciwmikrobiologicznych i przeciwzapalnych białka, stąd modyfikacje w jego obrębie mogą wpływać na siłę działania LF. Jak wiadomo zespół metaboliczny wiąże się z przewlekłym zapaleniem i endotoksemią, zatem ten aspekt aktywności LF może mieć niebagatelne znaczenie. W cytowanym badaniu [45], podobnie jak w poprzedniej obserwacji, poziom krążącej LF korelował ujemnie z BMI, stężeniem TG i glukozy we krwi, a dodatnio ze stężeniem HDL. Nie wpływał na reaktywność ściany naczyń krwionośnych (mierzoną jako wazodylatację zależną od śródbłonka) u osób zdrowych, ale korelował dodatnio z tym wskaźnikiem u otyłych pacjentów z nietolerancją glukozy.
Równie ciekawych wyników dostarczyła obserwacja kliniczna u 45 pacjentów z chorobliwą otyłością (średni BMI 53,4 kg/m2) [15]. Zanotowano korelację poziomu krążącej LF z poziomem lipidów oraz parametrami stresu oksydacyjnego i zapalenia po spożyciu bogatotłuszczowego posiłku. Po takim posiłku znacznie wzrasta osoczowy poziom TG (tzw. poposiłkowa hiperlipidemia), co nasila stres oksydacyjny, pogarsza funkcję śródbłonka naczyniowego i sprzyja rozwojowi choroby wieńcowej. U osób, u których po posiłku poziom LF wzrósł bardziej, niższe było stężenie wolnych kwasów tłuszczowych oraz mniejszy spadek poziomu HDL. Co istotne, mniej nasilone były procesy peroksydacji lipidów, większa aktywność katalazy i reduktazy glutationu oraz całkowita aktywność antyoksydacyjna, co świadczy o złagodzeniu stresu oksydacyjnego. Niższe poziomy CRP wskazują natomiast na ograniczenie stanu zapalnego. W tym samym badaniu izolowano od pacjentów tkankę tłuszczową, którą następnie inkubowano z dodatkiem LF. Zanotowano znacznie większe przeżycie komórek oraz niższe stężenia TG w medium hodowlanym.
Powyższe obserwacje dostarczają ważnych dowodów na udział LF w gospodarce węglowodanowej i lipidowej ustroju. Spadek poziomu krążącego białka łączy się z nasileniem stanu zapalnego i obniżoną tolerancją glukozy, integralnymi składowymi zespołu metabolicznego. Wzrost poziomu LF chroni przed nadmierną lipidemią, stresem oksydacyjnym i zapaleniem po spożyciu dużej ilości tłuszczu.
Mechanizmy regulacyjnego działania LF
Jednym z możliwych mechanizmów działania LF jest wpływ na adipogenezę (tworzenie komórek tłuszczowych), który potwierdzono w licznych testach in vitro. Większość wyników wskazuje na antyadipogenne działanie białka. W pierwszym z badań LF hamowała różnicowanie w kierunku adipocytów mysich komórek linii MC3T3-G2/PA6 wyprowadzonych pierwotnie z kości czaszki [76]. Działaniu LF towarzyszyło obniżenie ekspresji genów białek uczestniczących w adiopogenezie: adiponektyny (adiponectin, ADIPOQ) i aP2 (adipocyte/macrophage fatty acid-binding protein) oraz czynnika transkrypcyjnego C/EBPα (CCAAT/enhancer binding protein α), który należy do głównych regulatorów procesu różnicowania adipocytów. Mniejsze było ponadto wytwarzanie izoformy γ receptora aktywowanego proliferacją peroksysomalną (peroxisome proliferator-activated receptor, PPARγ). Ekspresja tego receptora jądrowego stanowi jeden z wcześniejszych etapów różnicowania adipocytów. Kiedy w fibroblastach ulega on nadekspresji, komórki te różnicują się w kierunku adipocytów, a sygnały regulujące PPARγ mogą regulować metabolizm lipidów i glukozy. Aktywacja PPARγ stymuluje wytwarzanie i wydzielanie przez komórki tłuszczowe adiponektyny, hormonu, który reguluje wiele procesów metabolicznych, w tym katabolizm kwasów tłuszczowych i wykorzystanie glukozy w wątrobie i mięśniach, pośrednio wpływając na wrażliwość komórek na insulinę. Poziom adiponektyny koreluje odwrotnie z ilością tkanki tłuszczowej w organizmie – zmniejszenie otyłości zwiększa stężenie tego hormonu we krwi. Adiponektyna należy do adipokin „ochronnych” działając korzystnie nie tylko na przemiany lipidów i glukozy, ale też wykazując działanie przeciwzapalne i przeciwmiażdżycowe [25]. A zatem hamowanie jej wytwarzania przez LF nie wydaje się korzystne, ale może być niewątpliwie uznane za marker hamowania adipogenezy.
W dwu kolejnych testach LF hamowała tworzenie adipocytów z komórek linii 3T3-L1 (preadipocytów) wyprowadzonych z mysiego zarodka [47,51]. Jednocześnie z zahamowaniem różnicowania komórek i tworzenia kropelek tłuszczu, obserwowano obniżenie ekspresji markerów lipogenezy: syntazy kwasów tłuszczowych (fatty acid synthase, FASN), α-karboksylazy acetylo-CoA (acetyl-coenzyme A carboxylase α, ACC) oraz PPARγ. Stwierdzono ponadto zwiększoną aktywację kinazy białkowej aktywowanej AMP (AMP-activated protein kinase, AMPK) poprzez fosforylację reszty Thr172 w obrębie jednostki katalitycznej białka. Ta AMPK katalizuje fosforylację ACC, która w ten sposób traci swoją aktywność, hamując adipogenezę. LF nasilała również ekspresję i hipofosforylację białka Rb (retinoblastoma), które w stanie hipofosforylacji jest aktywne i może przerywać adipogenezę [47].
Co ciekawe, nieco późniejsze badania tej samej grupy dały inne wyniki [48]. Były to testy in vitro na ludzkich preadipocytach wyizolowanych z tkanki tłuszczowej podskórnej i wisceralnej stymulowanych w odpowiedniej hodowli do różnicowania w kierunku komórek tłuszczowych. Dodatek HLF (1 i 10 µM) stymulował proces adipogenezy, co łączyło się ze zwiększeniem ekspresji genów i wytwarzania białek związanych z tym procesem: FASN, PPARγ, ACC, adiponektyny, adipocytowego białka wiążącego kwasy tłuszczowe (fatty acid binding protein 4, adipocyte, FABP4) oraz STAMP2 (six-transmembrane protein of prostate 2). Dodatek LF obniżał ponadto aktywność (172Thr)AMPK, zwiększając z kolei aktywność ACC, umożliwiając tworzenie kwasów tłuszczowych. Wzrastała również fosforylacja Rb, czyniąc białko nieaktywnym. Autorzy uzyskane w kolejnych testach różnice tłumaczą odmiennym modelem badawczym (mysie komórki linii 3T3-L1 vs ludzkie pierwotne preadipocyty). W opisanych testach, regulacyjne działanie LF wiązało się z hamowaniem wytwarzania czynników zapalnych (IL-6, IL-8 oraz MCP-1) przez komórki tłuszczowe oraz uwrażliwieniem ich na insulinę, autorzy zatem całościowy efekt działania LF uznają za korzystny.
W kolejnym badaniu [53] BLF hamowała tworzenie komórek tłuszczowych z preadipocytów uzyskanych z krezkowej i podskórnej tkanki tłuszczowej szczurów. Podobnie jak w poprzednich testach, obserwowano obniżenie ekspresji: C/EBPα, β i δ oraz PPARγ. LF trawiona trypsyną zachowała swoją aktywność, podczas gdy białko trawione pepsyną było nieaktywne. Uzyskane wyniki niosą również praktyczną informację: podawane doustnie preparaty LF powinny być chronione przed działaniem soków żołądkowych. Na preadipocytach stwierdzono ekspresję receptorów LRP1 [53], które mogą być zaangażowane w działanie LF.
Receptory LRP1 (low density lipoprotein receptor-related protein 1, dawniej określane jako LRP), należą do rodziny receptorów LDL. Występują na większości komórek organizmu (w tym na hepatocytach, fibroblastach, komórkach śródbłonka, makrofagach i adipocytach) i zapewniają główną drogę wejścia cholesterolu egzogennego do komórki. LRP1 zawierają pojedynczą domenę przebijającą błonę komórkową, i podobnie do receptora LDL, skupione są na powierzchni błony komórkowej w dołkach opłaszczonych klatryną. Po związaniu z receptorem, cząsteczki liganda w stanie nienaruszonym wchodzą do komórki w wyniku endocytozy. Receptory po odłączeniu liganda są ponownie transportowane na powierzchnię komórki (recyklizacja receptora). LRP1 wiążą i warunkują internalizację ligandów związanych z metabolizmem lipidów: apolipoproteiny E (apo-E), resztkowych chylomikronów („remnantów”), lipoprotein o bardzo małej gęstości (VLDL), HDL, lipoproteiny a (Lpa) i lipazy lipoproteinowej. Wiążą i internalizują również wiele innych ligandów, m.in.: α2-makroglobulinę, aktywatory plazminogenu, kompleks aktywator-inhibitor plazminogenu, metaloproteinazy, egzotoksynę A Pseudomonas, rhinowirus, fibronektynę, trombospondynę, składnik C3 dopełniacza oraz LF [37]. Łącznie LRP1 „rozpoznają” około 30 różnych ligandów. Poza funkcją w endocytozie ligandów, są również receptorami sygnałowymi aktywującymi szlaki sygnałowe komórki [19,20,43]. Wewnątrzkomórkowe białko o masie 39 kDa nazwane RAP (receptor-associated protein) może hamować wiązanie wszystkich tych ligandów do LRP1 i jest używane w testach badających aktywność tego receptora.
Cząsteczka LF wiążąc się z LRP1 konkuruje z wiązaniem lipoprotein zawierających w swoim składzie apo-E, głównie resztek chylomikronów oraz VLDL (ryc. 3). Jak się okazało, bogata w reszty Arg sekwencja N-końca LF przypomina strukturę w cząsteczce apo-E rozpoznawaną przez LRP1 [24,78]. Wiązanie LF do LRP1 na różnych komórkach wykazano w wielu testach in vitro oraz in vivo [19,24,28,40,43,74,78]. Meilinger i wsp. w testach na szczurach wykazali, że wiązanie LF do LRP1 na hepatocytach warunkowało internalizację białka i jego usuwanie z krążenia po podaniu dożylnym [40]. LF podana i.v. królikom znacznie ograniczała ponadto pobór „remnantów” przez wątrobę, co potwierdzono w testach na świeżo wyizolowanych hepatocytach szczurzych i hepatocytach ludzkich linii HepG2 [24]. Podana dożylnie szczurom LF pozbawiona enzymatycznie 14 aa z N-końca wiązała się do LRP1 na hepatocytach i redukowała wątrobowy pobór VLDL o 50%. Natywna LF wchodziła w interakcje głównie z innymi receptorami na komórkach wątroby i w mniejszym stopniu konkurowała o wiązanie z VLDL [78]. Gdy BLF podano dożylnie myszom 1 min przed podaniem zawiesiny chylomikronów, obserwowano zahamowane usuwanie z krążenia przez wątrobę chylomikronów resztkowych, choć autorzy uwzględniają możliwy udział innego niż LRP1 receptora [8]. BLF dodana do hodowli komórkowych znacznie hamowała pobór VLDL przez hepatocyty linii HepG2, komórki jajnika chomika CHO oraz wyizolowane od ludzi fibroblasty [28]. Willnow i wsp. donieśli natomiast, że BLF związana do LRP1 na ludzkich fibroblastach blokowała zależną od LRP1 syntezę estrów cholesterolu, obserwowaną po dostarczeniu do komórek VLDL i kompleksów lipaza lipoproteinowa-VLDL [74].
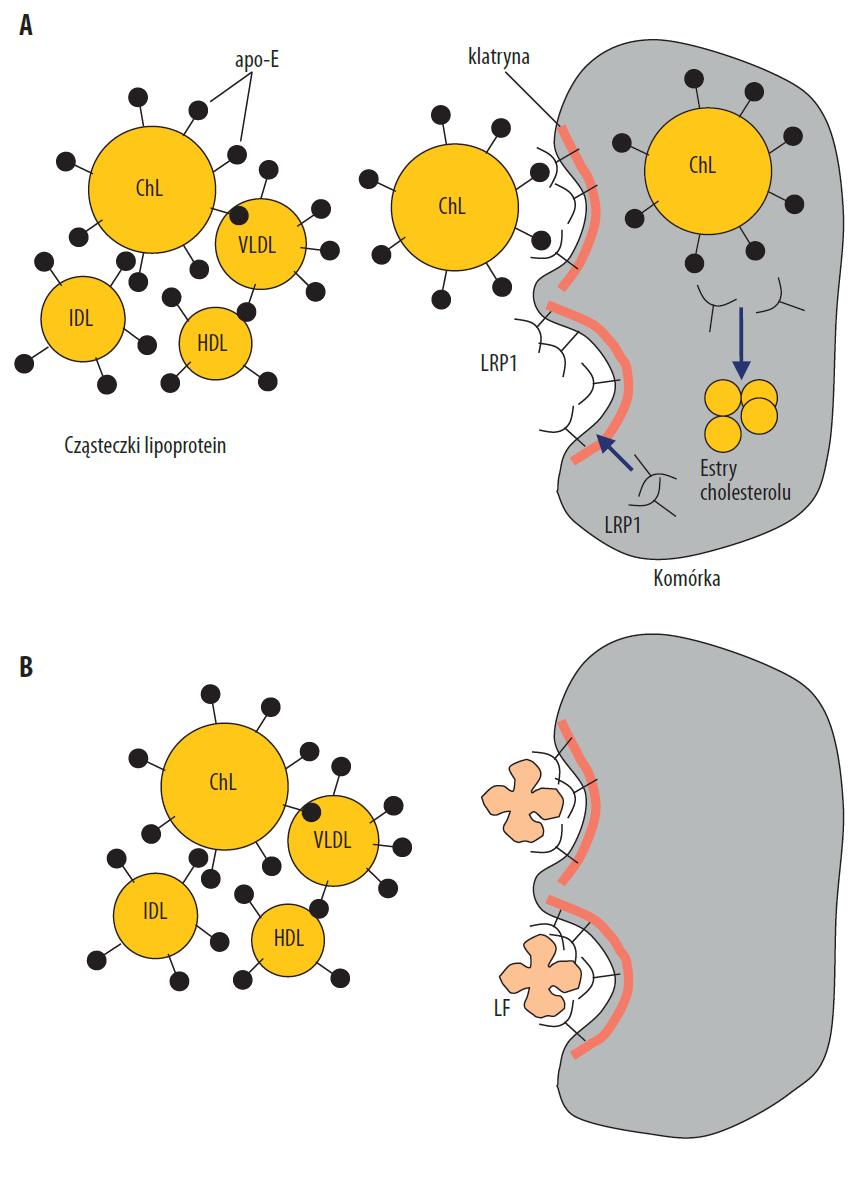
Ryc. 3. Cząsteczki lipidów: chylomikrony (ChL), VLDL, lipoproteiny o pośredniej gęstości (IDL), HDL, zawierające w swoim składzie apo-E, wiążą się do receptorów LRP1 na powierzchni komórek (m.in. wątroby, śródbłonka, fibroblastów, adipocytów), po czym następuje ich internalizacja, tj. wciągnięcie do wnętrza komórki w wyniku endocytozy. W komórce cholesterol uwolniony z lipoprotein ulega dalszym przemianom, a receptor LRP1 wraca na powierzchnię komórki (recyklizacja) (A). Cząsteczki LF wiążą się z receptorami LRP1 na powierzchni komórki i uniemożliwiają związanie lipoprotein i ich internalizację (B). Receptory LRP1 grupują się w zagłębieniach komórki, pokrytych białkiem klatryną, która wyściela od wewnątrz błonę komórkową
Jak się wydaje, zasadnicze znaczenie dla widocznych skutków działania LF ma komórkowa lokalizacja receptorów LRP1, do których białko się wiąże. LF wiążąc się z LRP1 na hepatocytach blokuje wiązanie resztkowych chylomikronów oraz VLDL, a tym samym ogranicza ich sprawne usuwanie z krążenia [78]. O ile pojawiające się w krążeniu po posiłku chylomikrony nie są aterogenne, o tyle powstałe w wyniku ich przemian w tkankach resztkowe chylomikrony mają takie właściwości. Stąd diety bogate w tłuszcze (zwłaszcza nasycone) i powodujące przez to silniejszą lipemię poposiłkową mogą być powodem miażdżycy właśnie przez podwyższenie stężenia „remnantów” [63]. Chylomikrony resztkowe to kompleksy wzbogacone w estry cholesterolu, które podobnie jak VLDL, IDL oraz LDL są w stanie przenikać z osoczem do ścian tętnic, gdzie podlegają przemianom metabolicznym, których skutkiem jest rozwój ognisk miażdżycy. „Remnanty” są transportowane z krwią do wątroby, gdzie wiążą się z receptorami LRP1 na powierzchni hepatocytów, a następnie są przyswajane i metabolizowane. Retencja „remnantów” w osoczu może zatem wynikać z ich nie dość sprawnego usuwania przez wątrobę. W komórkach wątroby składniki chylomikronów resztkowych są hydrolizowane uwalniając: aminokwasy, cholesterol i kwasy tłuszczowe, które mogą być wykorzystane do odpowiednich syntez. Co ważne, cholesterol uwolniony z „remnantów” zmniejsza syntezę cholesterolu endogennego przez hepatocyty. W wątrobie część cholesterolu jest przekształcana w kwasy żółciowe przechodzące do żółci, z którą trafiają do przewodu pokarmowego i dalej do krążenia wątrobowo-jelitowego żółci, bądź są wydalane z kałem. Wątroba zatem odgrywa zasadniczą rolę w utrzymaniu równowagi lipidowej ustroju, a pobór resztkowych chylomikronów musi być efektywny tak, by sprawnie usunąć krążący cholesterol i wydalić do żółci, co chroni przed jego gromadzeniem w tkankach pozawątrobowych.
O ile hamowanie wiązania lipidów przez wątrobę należy więc raczej uznać za niekorzystny aspekt działania LF, to wiązanie białka z receptorami LRP1 na innych komórkach (np. fibroblastach i adipocytach) może ograniczać pobór lipidów (w postaci cząsteczek chylomikronów, VLDL oraz IDL) przez te komórki i ich gromadzenie w postaci tkanki tłuszczowej. Jak już wspomniano, ekspresję LRP1 obserwowano na preadipocytach szczurzych [53]. Ich obecność stwierdzono również na komórkach trzewnej tkanki tłuszczowej u myszy, gdzie wykazano, że LRP1 jest głównym regulatorem homeostazy energetycznej adipocytów, modulującym osoczowy transport lipidów po posiłku i homeostazę glukozy [23]. Myszy z nokautem genu LRP1 w adipocytach miały zredukowaną masę ciała, mniejsze zapasy tłuszczu, lepszą tolerancję glukozy oraz zwiększone zużycie energii z powodu nasilonej termogenezy w mięśniach. Nie rozwijała się u nich otyłość i insulinooporność indukowana bogatotłuszczową dietą [23].
Rozpoznawana przez receptor LRP1 apo-E, poza cząsteczkami chylomikronów, VLDL oraz IDL, wchodzi również w skład lipidów HDL [41,63]. Zatem również pobór tych cząsteczek może być hamowany przez LF, co potwierdziły testy in vitro. Bydlęca LF hamowała o 35-50% pobór estrów cholesterolu HDL przez ludzkie pierwotne adipocyty oraz komórki tłuszczakomięsaka, a działanie to zależało od selektywnego wiązania LF do LRP1 [70]. Cząsteczki HDL pochodzą z różnych źródeł, pierwotnie są wydzielane z wątroby i jelita, ale powstają również we krwi, głównie z produktów rozpadu innych lipoprotein (chylomikronów i VLDL). Lipoproteiny HDL nie na darmo są określane jako „dobry cholesterol”, mogą bowiem łatwo przechodzić z osocza do płynu międzykomórkowego, gdzie wiążą cholesterol nagromadzony w komórkach (w tym ścian tętnic) i przenoszą go do wątroby, gdzie podlega dalszym przemianom lub jest usuwany z żółcią. Dyskutowane są również inne możliwe mechanizmy działania tych lipoprotein. Obecnie jednak właściwości antyaterogenne HDL nie budzą wątpliwości: duża ich zawartość hamuje rozwój miażdżycy, a mała stanowi czynnik zagrożenia. Lepszym wskaźnikiem ryzyka jest stosunek poziomu cholesterolu całkowitego do cholesterolu HDL niż sam poziom HDL. Pożądana wartość tego stosunku to 5,0 lub mniej [63]. Stężenie HDL jest podnoszone przez: wysiłek fizyczny, niewielkie ilości alkoholu, dietę z przewagą mięsa rybiego, a obniżane przez: otyłość, cukrzycę, palenie papierosów, brak ruchu. Cytowane badania wskazują, że również LF może działać korzystnie na poziom HDL.
Jak wykazano wyżej, związanie się LF do receptorów LRP1 na powierzchni komórek może je blokować i korzystnie wpływać na homeostazę lipidów i glukozy. Wydaje się to tym bardziej prawdopodobne, że jak już wspomniano, receptory te nie tylko uczestniczą w endocytozie ligandów, ale też pełnią funkcję sygnałową przekazując sygnał aktywujący do komórki. W ten sposób LF działa na osteoblasty, fibroblasty [20] i chondrocyty [5], aktywując, po związaniu do LRP1, szlak sygnałowy MAPK Erk 1/2 i stymulując proliferację tych komórek.
Wychwytywanie LF przez tkankę tłuszczową jest tym łatwiejsze, że LF po podaniu doustnym jest transportowana do krążenia przez limfatyczny przewód piersiowy [65], podobnie jak triglicerydy (w postaci chylomikronów) wchłonięte z jelita po posiłku tłuszczowym. Cząsteczki chylomikronów trafiają następnie do tkanek obwodowych, w tym do tkanki tłuszczowej, gdzie pozbywają się dużej części triglicerydów i ulegają przemianom do chylomikronów resztkowych. Do tkanki tłuszczowej wraz z lipidami może też trafiać podana doustnie LF, co potwierdzono w testach na szczurach. BLF w dużej dawce (1000 mg w postaci roztworu) zaaplikowana szczurom przez sondę żołądkową była wykryta immunoenzymatycznie w krezkowej tkance tłuszczowej w ilości 14,4 µg/g tkanki po 15 min od podania [53].
Kolejny mechanizm działania antyaterogennego LF może polegać na hamowaniu powstawania tzw. komórek piankowatych, uczestniczących w tworzeniu blaszek miażdżycowych w ścianie naczyń krwionośnych. Komórki piankowate powstają z makrofagów i komórek mięśni gładkich, które zostają naładowane lipidami, pobieranymi nieprawidłową drogą receptorową, która podlega ścisłej kontroli, ale poprzez słabiej kontrolowany tzw. szlak zmiataczowy z udziałem receptora zmiataczowego (scavenger receptor) [3]. Jak się obecnie uważa, za uruchomienie tego nieprawidłowego szlaku poboru lipidów są odpowiedzialne modyfikacje cząsteczek lipoprotein LDL, a konkretnie ich części białkowej, czyli apo-B, w wyniku czego powstają ox-LDL. Cząsteczka LDL zawiera w swoim składzie m.in. reszty wielonienasyconych kwasów tłuszczowych, podatne na peroksydację. Przed utlenianiem chronią je jednak obecne w LDL antyoksydanty: α-tokoferol, β-karoten i ubihydrochinon 10 (zredukowany koenzym Q-10). Procesy peroksydacji jednak w pewnym zakresie zachodzą, być może katalizowane enzymatycznie, czemu mogą sprzyjać jony żelaza i miedzi obecne w złogach miażdżycowych [3]. Powstające ox-LDL oddziałują z receptorami zmiataczowymi, głównie makrofagów, i przekazują im duże ilości lipidów, przekształcając je w komórki piankowate, które stają się podstawowymi składnikami blaszek miażdżycowych. Badania sprzed prawie 20 lat wykazały, że LF może blokować wiązanie cząsteczek ox-LDL do makrofagów [29]. LF (ludzka i bydlęca) zmniejszała o ponad 80% akumulację estrów cholesterolu w szczurzych makrofagach inkubowanych z acetylowanymi lub ox-LDL. Modyfikacja reszt Arg w N-końcowej części cząsteczki znosiła działanie LF. Jaki zatem jest mechanizm działania białka? Jak wynika z nieco późniejszych badań tego samego zespołu, kationowe zgrupowania aminokwasów zasadowych w cząsteczce LF mogą wchodzić w interakcje z ujemnie naładowanymi cząsteczkami ox-LDL, blokując zatem ich rozpoznanie i związanie przez receptory resztkowe na komórkach [60] (ryc. 4).
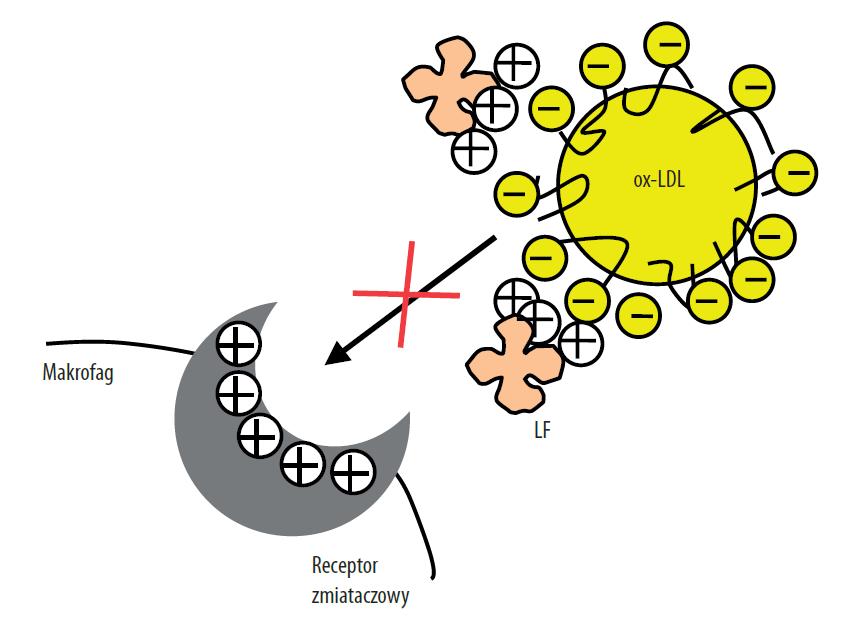
Ryc. 4. Mechanizm blokowania przez LF wiązania ox-LDL do receptora zmiataczowego na powierzchni makrofagów. Kationowe zgrupowania aminokwasów w cząsteczce LF wchodzą w interakcje z ujemnie naładowaną powierzchnią cząsteczek ox-LDL, hamując ich rozpoznanie przez receptor zmiataczowy (wg [60], zmodyfikowano)
Innym możliwym mechanizmem działania LF jest, wspomniane już, przerwanie krążenia wątrobowo-jelitowego kwasów żółciowych [59,66]. Kwasy żółciowe powstające w wątrobie w wyniku skomplikowanych przemian cholesterolu są wydzielane do żółci (jako sole kwasów żółciowych), z którą trafiają do jelita cienkiego. Tutaj są przekształcane przez bakterie jelitowe i jako kwasy żółciowe wtórne absorbowane do krążenia wrotnego, z którym ponownie trafiają do wątroby. Wątroba przetwarza zarówno pierwotne, jak i wtórne kwasy żółciowe i znów wydziela je do żółci. Takie stałe krążenie kwasów żółciowych między wątrobą a jelitem jest nazywane wątrobowo-jelitowym krążeniem żółci i pozwala na wielokrotne wykorzystanie kwasów żółciowych podczas wchłaniania substancji tłuszczowych w jelicie. W ciągu doby wątroba wydziela do jelita ok. 15-30 g soli kwasów żółciowych, z czego jedynie 0,5 g jest wydalane i w przybliżeniu tyle samo powstaje w wątrobie, aby wyrównać ubytek. Mechanizm działania niektórych leków przeciwmiażdżycowych polega na hamowaniu krążenia wątrobowo-jelitowego żółci (związki te wiążą i wydalają kwasy żółciowe z kałem, przerywając ich wątrobowo-jelitowe krążenie, co powoduje kompensacyjny wzrost gęstości receptorów na hepatocytach oraz większy wychwyt miażdżycorodnych LDL przez wątrobę).
Inne mechanizmy korzystnego działania LF w normalizacji procesów metabolicznych mogą obejmować działanie przeciwzapalne, związane z neutralizacją LPS oraz modulacją wydzielania i/lub aktywności cytokin, chemokin, czynników wzrostowych, cząsteczek kostymulujących, adhezyjnych i innych, które uczestniczą w rozwoju zapalenia [35]. Stwierdzono ponadto wiązanie LF do produktów glikacji białek (AGE) [68]. Związki te powstają na skutek nieenzymatycznej glikozylacji białek i występują niemal we wszystkich ludzkich tkankach. Ich ilość wzrasta z procesami starzenia się i rozwoju niektórych schorzeń (miażdżycy, cukrzycy, arteriosklerozy, nefropatii, chorób neurodegeneracyjnych). Receptory AGE występują na wielu komórkach, m.in. monocytach, makrofagach, komórkach śródbłonka, mikrogleju, astrocytach. Interakcja AGE z komórkami prowadzi do ich aktywacji: uwolnienia cytokin prozapalnych, czynników wzrostu i wywołania stresu oksydacyjnego. AGE są jednym z czynników kojarzonych z rozwojem zapalenia podczas zespołu metabolicznego. Można sądzić, że związanie AGE przez LF będzie utrudniało ich wiązanie do komórek, a zatem osłabiało aktywność prozapalną. Przerwanie kaskady procesów zapalnych przez LF może ograniczyć rozwój zaburzeń metabolicznych charakterystycznych dla zespołu metabolicznego.
Jak wykazały badania sprzed kilku lat, otyłość i insulinooporność mogą się wiązać z deficytem IL-18 w ustroju [52]. U myszy z nokautem genu IL-18 lub jej receptora stwierdzono hiperfagię, otyłość, hiperglikemię oraz obniżoną wrażliwość na insulinę. Molekularny mechanizm zaburzeń obejmował zwiększoną ekspresję genów związanych z glukoneogenezą wskutek defektywnej fosforylacji czynnika STAT3. Korzystne działanie LF w otyłości i insulinooporności może zatem wynikać z indukcji IL-18. Wpływ LF na wydzielanie tej cytokiny potwierdzono w układach in vitro oraz testach na zwierzętach i ludziach. Na przykład, znacznie zwiększone osoczowe poziomy IL-18 obserwowano u pacjentów z zapaleniem wątroby typu C leczonych bydlęcą LF [26]. Przez niektórych badaczy IL-18 uważana jest nawet za farmakodynamiczny wskaźnik aktywności LF podanej doustnie [22].
Jak już wspomniano, endogenna LF była wyraźnie skorelowana ze wskaźnikami metabolizmu glukozy zarówno u osób zdrowych, jak i otyłych oraz z nietolerancją glukozy [15,45,46]. To sugeruje udział białka w przemianach glukozy, co potwierdzono w cytowanych już testach in vitro. LF mianowicie stymulowała fosforylację indukowanej przez insulinę kinazy (473Ser)Akt, której fosforylacja i aktywność są obniżone w stanie oporności komórek na insulinę. Białko działało w podobny sposób zarówno na mysie komórki linii 3T3-L1 oraz ludzkie linii HepG2 [47], jak i preadipocyty ludzkie z tkanki podskórnej i brzusznej [48]. LF była aktywna również gdy wrażliwość komórek na insulinę jest mniejsza (obecność cytokin prozapalnych lub deksametazonu w hodowli) [47]. Jednocześnie z aktywacją (473Ser)Akt obserwowano lepsze wykorzystanie glukozy przez komórki inkubowane z LF. W cytowanym badaniu LF działała w zbliżony sposób na komórki tkanki tłuszczowej podskórnej i wisceralnej.
LF ponadto działała regulacyjnie na układ insulinopodobnych czynników wzrostu (insulin-like growth factor, IGF) [4]. Obejmuje on zarówno białkowe czynniki wzrostu komórek, jak i ich receptory oraz białka regulacyjne: insulinę, IGF-1, IGF-2; 3 receptory dużego powinowactwa: IGF-1R, IGF-2R i receptor insulinowy (insulin-R); 6 białek wiążących IGF (IGF binding protein, IGFBP), które wiążą IGF z dużym powinowactwem oraz 9 białek związanych z IGFBP (IGFBP-related protein, IGFBP-rP), które wiążą IGF z małym powinowactwem. Składniki systemu IGF są powszechne w organizmie, działając jako mediatory wzrostu, rozwoju i różnicowania oraz czynniki endokrynne zapobiegające apoptozie. Badania nad bydlęcą LF wykazały, że białko to może swoiście wiązać IGFBP-3 w mleku krowim oraz hodowli komórek nabłonkowych wymienia [4]. Związanie IGFBP-3 przez cząsteczkę LF usuwało z kompleksu IGF, co może zmieniać aktywność komórek przez umożliwienie wiązania IGF do receptora IGF-1R. Autorzy uważają, że zjawisko to może mieć szczególne znaczenie w tkance wymienia podczas zapalenia oraz przed porodem, kiedy ilości LF wydzielanej przez komórki są większe.
Badania z 2010 r. wykazały ponadto, że bydlęca LF wiąże cząsteczki cukrów [42]. Uzyskany poprzez ograniczoną proteolizę enzymatyczną natywnej LF płat C wiązał cząsteczki różnych cukrów spożywczych (m.in. glukozę, galaktozę, mannozę, maltozę, laktozę, sacharozę) ze stałymi wiązania 10-4-10-5 M. To sugeruje, że płat C podany doustnie mógłby sekwestrować cząsteczki cukrów przyjęte z pożywieniem, ograniczając/opóźniając ich wchłanianie do krążenia. W ten sposób (tzn. przez spowolnienie absorpcji węglowodanów w jelicie) działają niektóre leki przeciwcukrzycowe. W dodatku, w testach in vitro płat C cząsteczki LF dodany (1 mg/ml) do osocza krwi zdrowych osób obniżał stężenie glukozy średnio ze 100 mg/ml do 85 mg/ml (zakres redukcji to 10-23% względem kontroli) [42]. Płat C cząsteczki LF jest oporny na proteolizę enzymatyczną [58], co umożliwia jego użycie w postaci doustnej. Zaprezentowane wyniki sugerują możliwość zastosowania tego fragmentu cząsteczki LF w leczeniu cukrzycy typu 2.
Zastosowanie LF u diabetyków może przynieść dodatkową korzyść, wpływając na gojenie ran i owrzodzeń, które są częstym powikłaniem cukrzycy. Trudno gojące się rany, przechodzące w owrzodzenia, to skutek obwodowej neuropatii, zmniejszonego dopływu krwi, ischemii i zakażeń. Cechą ran cukrzycowych jest chroniczny proces zapalny, nasilany przez RFT i nadkażenia bakteryjne, często przekształcające się w trudny do wyleczenia biofilm bakteryjny. Większa podatność na infekcje bakteryjne u cukrzyków może być skutkiem m.in. podniesionych poziomów AGE w tkankach i surowicy, które wiążąc się z białkami przeciwbakteryjnymi (w tym LF i lizozymem) ograniczają ich aktywność [36]. Mechanizm ochronnego działania białka podczas gojenia ran obejmuje m.in. regulację procesów zapalnych, wpływ na aktywność fibroblastów i keratynocytów oraz procesy angiogenezy [64]. Jak dotąd, przeprowadzono próby kliniczne I i II fazy (n=55) z użyciem rHLF w leczeniu ran cukrzycowych. Wykazano brak niepożądanych działań oraz skuteczność białka: w grupie stosującej LF miejscowo w postaci żelu dwukrotnie częściej dochodziło do zagojenia rany niż w grupie placebo [38]. Aktywność białka potwierdzono na modelu myszy diabetycznych db/db, gdzie rHLF użyta miejscowo promowała gojenie ran [14].
Wyniki testów przedklinicznych i klinicznych oraz obserwacji klinicznych wykazujące udział LF w metabolizmie glukozy i lipidów oraz regulacji ciśnienia krwi z uwzględnieniem możliwych mechanizmów działania białka zebrano w tabeli 1.
Tabela 1. Zestawienie testów przedklinicznych i klinicznych oraz obserwacji klinicznych wykazujących udział LF w metabolizmie glukozy i lipidów oraz regulacji ciśnienia krwi z uwzględnieniem możliwych mechanizmów działania białka

Podsumowanie
LF wydaje się obiecującym środkiem dietetycznym pomocnym w profilaktyce i leczeniu chorób metabolicznych: otyłości, nietolerancji glukozy/cukrzycy typu 2, dyslipidemii oraz nadciśnienia. Tym bardziej że jak dotąd brak związków o tak szerokim zakresie działania, które jednocześnie regulowałyby homeostazę lipidów i glukozy oraz ciśnienie krwi. Co należy podkreślić, w żadnym z dotychczas przeprowadzonych testów na zwierzętach i ludziach LF nie była toksyczna i nawet jej długotrwałe przyjmowanie w dużych dawkach nie powodowało żadnych działań niepożądanych [39,67]. Amerykańska Agencja ds. Żywności i Leków (Food and Drug Administration, FDA) nadała LF bydlęcej status GRAS (generally recognized as safe), czyli produktu uznawanego za bezpieczny [7] i na tej podstawie BLF jest używana jako suplement diety i składnik różnych produktów spożywczych, w tym mieszanek mlecznych dla niemowląt. Niedawno również Europejski Urząd ds. Bezpieczeństwa Żywności (European Food Safety Authority, EFSA) uznał BLF za bezpieczny składnik żywności [11]. Całkowite bezpieczeństwo użycia LF jest tym ważniejsze, że wiele z leków używanych w otyłości, cukrzycy czy dyslipidemii odznacza się toksycznością i ma mniej lub bardziej poważne niekorzystne działania. I tak np. niektóre z leków hipolipemizujących (kwas nikotynowy i pochodne) obniżają wprawdzie cholesterol i TG, ale jednocześnie mogą pogarszać tolerancję glukozy, a ich działanie diabetogenne często staje się przeciwwskazaniem do ich użycia.
Bydlęca LF może być dodawana do produktów spożywczych, np. mleka lub jogurtu, których spożycie jest pożądane ze względu na obecność również innych, poza LF, aktywnych składników [79]. Jak już wspomniano, przyjmowanie LF w takiej postaci było skuteczne w leczeniu łojotokowych chorób skóry [31]. Jak dotąd, na rynku polskim nie są jednak dostępne produkty mleczne wzbogacone w LF, choć w niektórych państwach (np. Japonii) już od wielu lat można kupić m.in. mleko, sery i jogurty z dodatkiem bydlęcej LF [49]. BLF można również przyjmować w postaci preparatów – suplementów diety, których obecnie jest duży wybór, również w Polsce. Korzystne wydaje się łączenie LF z innymi składnikami, np. witaminami A, E i C o właściwościach przeciwutleniających oraz bakteriami probiotycznymi i prebiotykami (np. błonnikiem), które korzystnie wpływają na homeostazę organizmu, m.in. poprzez regulację składu mikroflory jelitowej. Niedawna próba kliniczna na otyłych osobach wykazała dużą skuteczność preparatu zawierającego aktywne peptydy serwatki mleka, leucynę oraz wapń (Prolibra®) [17]. Jego przyjmowanie w ciągu 12 tygodni znacznie przyspieszyło utratę tkanki tłuszczowej, chroniąc jednocześnie masę mięśniową.
Należy podkreślić, że przyjmowanie LF może stanowić tylko jeden z elementów w walce z otyłością i zaburzeniami metabolizmu energii i lipidów. Należy pamiętać przede wszystkim o ważnych zasadach dietetycznych, nie tylko o składzie i kaloryczności posiłków (mniej węglowodanów prostych, tłuszczów nasyconych i mniej kalorii), ale też sposobie ich spożywania: korzystniej często, ale w mniejszych ilościach. Już od dawna bowiem wiadomo, że dowóz całodziennej porcji energii w postaci jednorazowego obfitego posiłku prowadzi do zwiększenia stężenia cholesterolu oraz upośledzonej tolerancji glukozy i rozwoju otyłości [41]. Nie mniej ważny dla zachowania równowagi metabolizmu jest wpływ wysiłku fizycznego (czyli odpowiednie wydatkowanie dostarczonej energii). Co istotne, a często pomijane, to wpływ stresu psychicznego na metabolizm. Stwierdzono np., że u księgowych w okresie sporządzania bilansów, a u studentów w czasie sesji egzaminacyjnej, stężenie cholesterolu we krwi zwiększa się. Wyższe poziomy lipidów obserwowano też u osób o określonej konstrukcji psychicznej, tj. ambitnych, dążących do sukcesu, ze skłonnością do współzawodnictwa. Długotrwały stres, jak już wspomniano, wpływa m.in. na rozwój niebezpiecznej brzusznej postaci otyłości. W większości przypadków połączenie trzech wymienionych elementów: ustabilizowanie trybu życia, połączone ze zmianą diety oraz zwiększeniem aktywności fizycznej, prowadzi do skutecznego zmniejszenia masy ciała i normalizacji metabolizmu ustroju.
PIŚMIENNICTWO
[1] Artym J.: Udział laktoferryny w gospodarce żelazem w organizmie. Część I. Wpływ laktoferryny na wchłanianie, transport i magazynowanie żelaza. Postępy Hig. Med. Dośw., 2008; 62: 599-612
[PubMed] [Full Text HTML] [Full Text PDF]
[2] Artym J., Zimecki M.: The effects of lactoferrin on myelopoiesis: can we resolve the controversy? Postępy Hig. Med. Dośw., 2007; 61: 129-150
[PubMed] [Full Text HTML] [Full Text PDF]
[3] Bartosz G.: Druga twarz tlenu. Wolne rodniki w przyrodzie. Wydawnictwo Naukowe PWN, Warszawa, 2003
[4] Baumrucker C.R., Erondu N.E.: Insulin-like growth factor (IGF) system in the bovine mammary gland and milk. J. Mammary Gland Biol. Neoplasia, 2000; 5: 53-64
[PubMed]
[5] Brandl N., Zemann A., Kaupe I., Marlovits S., Huettinger P., Goldenberg H., Huettinger M.: Signal transduction and metabolism in chondrocytes is modulated by lactoferrin. Osteoarthritis Cartilage, 2010; 18: 117-125
[PubMed]
[6] Cani P.D., Amar J., Iglesias M.A., Poggi M., Knauf C., Bastelica D., Neyrinck A.M., Fava F., Tuohy K.M., Chabo C., Waget A., Delmée E., Cousin B., Sulpice T., Chamontin B., Ferrieres J., Tanti J.F., Gibson G.R., Casteilla L., Delzenne N.M., Alessi M.C., Burcelin R.: Metabolic endotoxemia initiates obesity and insulin resistance. Diabetes, 2007; 56: 1761-1772
[PubMed]
[7] CFSAN/Office of Food Additive Safety. Agency response letter: GRAS notice No. GRN 000077. U.S. Food and Drug Administration Web site (02.09.2012)
http://www.fda.gov/Food/FoodingredientsPackaging/GenerallyRecognizedasSafeGRAS/GRASListings/ucm154188.htm
[8] Crawford S.E., Borensztajn J.: Plasma clearance and liver uptake of chylomicron remnants generated by hepatic lipase lipolysis: evidence for a lactoferrin-sensitive and apolipoprotein E-independent pathway. J. Lipid Res., 1999; 40: 797-805
[PubMed] [Full Text HTML] [Full Text PDF]
[9] Dandona P., Aljada A., Bandyopadhyay A.: Inflammation: the link between insulin resistance, obesity and diabetes. Trends Immunol., 2004; 25: 4-7
[PubMed]
[10] Eckel R.H., Grundy S.M., Zimmet P.Z.: The metabolic syndrome. Lancet, 2005; 365: 1415-1428
[PubMed]
[11] EFSA Panel on Dietetic Products, Nutrition and Allergies (NDA): Scientific opinion on bovine lactoferrin. EFSA J., 2012; 10: 2701
[Abstract] [Full Text PDF]
[12] Egan B.M.: Insulin resistance and the sympathetic nervous system. Curr. Hypertens. Rep., 2003; 5: 247-254
[PubMed]
[13] Elgazar-Carmon V., Rudich A., Hadad N., Levy R.: Neutrophils transiently infiltrate intra-abdominal fat early in the course of high-fat feeding. J. Lipid Res., 2008; 49: 1894-1903
[PubMed] [Full Text HTML] [Full Text PDF]
[14] Engelmayer J., Blezinger P., Varadhachary A.: Talactoferrin stimulates wound healing with modulation of inflammation. J. Surg. Res., 2008; 149: 278-286
[PubMed]
[15] Fernández-Real J.M., Garcia-Fuentes E., Moreno-Navarrete J.M., Murri-Pierri M., Garrido-Sánchez L., Ricart W., Tinahones F.: Fat overload induces changes in circulating lactoferrin that are associated with postprandial lipemia and oxidative stress in severely obese subjects. Obesity, 2010; 18: 482-488
[PubMed]
[16] Ferrannini E., Buzzigoli G., Bonadonna R., Giorico M.A., Oleggini M., Graziadei L., Pedrinelli R., Brandi L., Bevilacqua S.: Insulin resistance in essential hypertension. N. Engl. J. Med., 1987; 317: 350-357
[PubMed]
[17] Frestedt J.L., Zenk J.L., Kuskowski M.A., Ward L.S., Bastian E.D.: A whey-protein supplement increases fat loss and spares lean muscle in obese subjects: a randomized human clinical study. Nutr. Metab., 2008; 5: 8
[PubMed] [Full Text HTML] [Full Text PDF]
[18] Gibbons J.A., Kanwar R.K., Kanwar J.R.: Lactoferrin and cancer in different cancer models. Front. Biosci., 2011; 3: 1080-1088
[PubMed]
[19] Goretzki L., Mueller B.M.: Low-density-lipoprotein-receptor-related protein (LRP) interacts with a GTP-binding protein. Biochem. J., 1998; 336: 381-386
[PubMed] [Full Text HTML] [Full Text PDF]
[20] Grey A., Banovic T., Zhu Q., Watson M., Callon K., Palmano K., Ross J., Naot D., Reid I.R., Cornish J.: The low density lipoprotein receptor-related protein 1 is a mitogenic receptor for lactoferrin in osteoblastic cells. Mol. Endocrinol., 2004; 18: 2268-2278
[PubMed] [Full Text HTML] [Full Text PDF]
[21] Hayashida K., Takeuchi T., Ozaki T., Shimizu H., Ando K., Miyamoto A., Harada E.: Bovine lactoferrin has a nitric oxide-dependent hypotensive effect in rats. Am. J. Physiol. Regul. Integr. Comp. Physiol., 2004; 286: R359-R365
[PubMed] [Full Text HTML] [Full Text PDF]
[22] Hayes T.G., Falchook G.F., Varadhachary G.R., Smith D.P., Davis L.D., Dhingra H.M., Hayes B.P., Varadhachary A.: Phase I trial of oral talactoferrin alfa in refractory solid tumors. Invest. New Drugs, 2006; 24: 233-240
[PubMed]
[23] Hofmann S.M., Zhou L., Perez-Tilve D., Greer T., Grant E., Wancata L., Thomas A., Pfluger P.T., Basford J.E., Gilham D., Herz J., Tschöp M.H., Hui D.Y.: Adipocyte LDL receptor-related protein-1 expression modulates postprandial lipid transport and glucose homeostasis in mice. J. Clin. Invest., 2007; 117: 3271-3282
[PubMed] [Full Text HTML] [Full Text PDF]
[24] Huettinger M., Retzek H., Hermann M., Goldenberg H.: Lactoferrin specifically inhibits endocytosis of chylomicron remnants but not α-macroglobulin. J. Biol. Chem., 1992; 267: 18551-18557
[PubMed] [Full Text PDF]
[25] Ibrahim M.M.: Subcutaneous and visceral adipose tissue: structural and functional differences. Obes. Rev., 2010; 11: 11-18
[PubMed]
[26] Ishii K., Takamura N., Shinohara M., Wakui N., Shin H., Sumino Y., Ohmoto Y., Teraguchi S., Yamauchi K: Long-term follow-up of chronic hepatitis C patients treated with oral lactoferrin for 12 months. Hepatol. Res., 2003; 25: 226-233
[PubMed]
[27] Jenssen H., Hancock R.E.: Antimicrobial properties of lactoferrin. Biochimie, 2009; 91: 19-29
[PubMed]
[28] Ji Z.S., Mahley R.W.: Lactoferrin binding to heparan sulfate proteoglycans and the LDL receptor-related protein. Further evidence supporting the importance of direct binding of remnant lipoproteins to HSPG. Arterioscler. Thromb., 1994; 14: 2025-2031
[PubMed] [Full Text PDF]
[29] Kajikawa M., Ohta T., Takase M., Kawase K., Shimamura S., Matsuda I.: Lactoferrin inhibits cholesterol accumulation in macrophages mediated by acetylated or oxidized low-density lipoproteins. Biochim. Biophys. Acta, 1994; 1213: 82-90
[PubMed]
[30] Karbowska A., Boratyńska M., Klinger M.: Rezystyna – czynnik patogenetyczny czy biomarker zaburzeń metabolicznych i zapalenia? Postępy Hig. Med. Dośw., 2009; 63: 485-491
[PubMed] [Full Text HTML] [Full Text PDF]
[31] Kim J., Ko Y., Park Y.K., Kim N.I., Ha W.K., Cho Y.: Dietary effect of lactoferrin-enriched fermented milk on skin surface lipid and clinical improvement of acne vulgaris. Nutrition, 2010; 26: 902-909
[PubMed]
[32] Kim J.Y., van de Wall E., Laplante M., Azzara A., Trujillo M.E., Hofmann S.M., Schraw T., Durand J.L., Li H., Li G., Jelicks L.A., Mehler M.F., Hui D.Y., Deshaies Y., Shulman G.I., Schwartz G.J., Scherer P.E.: Obesity-associated improvements in metabolic profile through expansion of adipose tissue. J. Clin. Invest., 2007; 117: 2621-2637
[PubMed] [Full Text HTML] [Full Text PDF]
[33] Kimoto H.: Case report of 4 patients on the improvement of serum lipids by the enteric-coated LF tablets. Progr. Medicine, 2003; 23: 1519-1523
[34] LaForce F.M., Boose D.S.: Release of lactoferrin by polymorphonuclear leukocytes after aerosol challenge with Escherichia coli. Infect. Immun., 1987; 55: 2293-2295
[PubMed] [Full Text PDF]
[35] Latorre D., Berlutti F., Valenti P., Gessani S., Puddu P.: LF immunomodulatory strategies: mastering bacterial endotoxin. Biochem. Cell Biol., 2012; 90: 269-278
[PubMed]
[36] Li Y.M., Tan A.X., Vlassara H.: Antibacterial activity of lysozyme and lactoferrin is inhibited by binding of advanced glycation-modified proteins to a conserved motif. Nat. Med., 1995; 1: 1057-1061
[PubMed]
[37] Lillis A.P., Van Duyn L.B., Murphy-Ullrich J.E., Strickland D.K.: LDL receptor-related protein 1: unique tissue-specific functions revealed by selective gene knockout studies. Physiol. Rev., 2008; 88: 887-918
[PubMed] [Full Text HTML] [Full Text PDF]
[38] Lyons T.E., Miller M.S., Serena T., Sheehan P., Lavery L., Kirsner R.S., Armstrong D.G., Reese A., Yankee E.W., Veves A.: Talactoferrin alfa, a recombinant human lactoferrin promotes healing of diabetic neuropathic ulcers: a phase 1/2 clinical study. Am. J. Surg., 2007; 193: 49-54
[PubMed]
[39] Manzoni P., Stolfi I., Messner H., Cattani S., Laforgia N., Romeo M.G., Bollani L., Rinaldi M., Gallo E., Quercia M., Maule M., Mostert M., Decembrino L., Magaldi R., Mosca F., Vagnarelli F., Memo L., Betta P.M., Stronati M., Farina D., Italian Task Force for the Study and Prevention of Neonatal Fungal Infections – the Italian Society of Neonatology: Bovine lactoferrin prevents invasive fungal infections in very low birth weight infants: a randomized controlled trial. Pediatrics, 2012; 129: 116-123
[PubMed]
[40] Meilinger M., Haumer M., Szakmary K.A., Steinbock F., Scheiber B., Goldenberg H., Huettinger M.: Removal of lactoferrin from plasma is mediated by binding to low density lipoprotein receptor-related protein/α2-macroglobulin receptor and transport to endosomes. FEBS Lett., 1995; 360: 70-74
[PubMed] [Full Text HTML] [Full Text PDF]
[41] Michajlik A., Sznajderman M.: Lipidy i lipoproteiny osocza. Państwowy Zakład Wydawnictw Lekarskich, Warszawa 1986
[42] Mir R., Kumar R.P., Singh N., Vikram G.P., Sinha M., Bhushan A., Kaur P., Srinivasan A., Sharma S., Singh T.P.: Specific interactions of C-terminal half (C-lobe) of lactoferrin protein with edible sugars: binding and structural studies with implications on diabetes. Int. J. Biol. Macromol., 2010; 47: 50-59
[PubMed]
[43] Misra U.K., Chu C.T., Gawdi G., Pizzo S.V.: The relationship between low density lipoprotein-related protein/α2-macroglobulin (α2M) receptors and the newly described α2M signaling receptor. J. Biol. Chem., 1994; 269: 18303-18306
[PubMed] [Full Text PDF]
[44] Moreno-Navarrete J.M., Fernández-Real J.M.: Antimicrobial-sensing proteins in obesity and type 2 diabetes: the buffering efficiency hypothesis. Diabetes Care, 2011; 34 (Suppl. 2): S335-S341
[PubMed] [Full Text HTML] [Full Text PDF]
[45] Moreno-Navarrete J.M., Ortega F.J., Bassols J., Castro A., Ricart W., Fernandez-Real J.M.: Association of circulating lactoferrin concentration and 2 nonsynonymous LTF gene polymorphisms with dyslipidemia in men depends on glucose-tolerance status. Clin. Chem., 2008; 54: 301-309
[PubMed] [Full Text HTML] [Full Text PDF]
[46] Moreno-Navarrete J.M., Ortega F.J., Bassols J., Ricart W., Fernández-Real J.M.: Decreased circulating lactoferrin in insulin resistance and altered glucose tolerance as a possible marker of neutrophil dysfunction in type 2 diabetes. J. Clin. Endocrinol. Metab., 2009; 94: 4036-4044
[PubMed]
[47] Moreno-Navarrete J.M., Ortega F.J., Ricart W., Fernandez-Real J.M.: Lactoferrin increases (172Thr)AMPK phosphorylation and insulin-induced (p473Ser)AKT while impairing adipocyte differentiation. Int. J. Obes., 2009; 33: 991-1000
[PubMed]
[48] Moreno-Navarrete J.M., Ortega F., Sabater M., Ricart W., Fernández-Real J.M.: Proadipogenic effects of lactoferrin in human subcutaneous and visceral preadipocytes. J. Nutr. Biochem., 2011; 22: 1143-1149
[PubMed]
[49] Morinaga Milk Industry Co. Ltd. (12.09.2012)
http://www.morinagamilk.co.jp/english/
[50] Mueller E.A., Trapp S., Frentzel A., Kirch W., Brantl V.: Efficacy and tolerability of oral lactoferrin supplementation in mild to moderate acne vulgaris: an exploratory study. Curr. Med. Res. Opin., 2011; 27: 793-797
[PubMed]
[51] Nam M.S., Cho M.C., Bae H.C., Yoon D.Y.: Effects of lactoferrin on adipogenesis in 3t3-l1 cells and obesity mice. Biochem. Cell Biol., 2006; 84: 399 (Abstracts of the 7th International Conference on Lactoferrin)
http://researchcommons.waikato.ac.nz/bitstream/handle/10289/1349/Bio%20and%20cell%20bio.pdf?sequence=1
[52] Netea M.G., Joosten L.A., Lewis E., Jensen D.R., Voshol P.J., Kullberg B.J., Tack C.J., van Krieken H., Kim S.H., Stalenhoef A.F., van de Loo F.A., Verschueren I., Pulawa L., Akira S., Eckel R.H., Dinarello C.A., van den Berg W., van der Meer J.W.: Deficiency of interleukin-18 in mice leads to hyperphagia, obesity and insulin resistance. Nat. Med., 2006; 12: 650-656
[PubMed]
[53] Ono T., Morishita S., Fujisaki C., Ohdera M., Murakoshi M., Iida N., Kato H., Miyashita K., Iigo M., Yoshida T., Sugiyama K., Nishino H.: Effects of pepsin and trypsin on the anti-adipogenic action of lactoferrin against pre-adipocytes derived from rat mesenteric fat. Br. J. Nutr., 2011; 105: 200-211
[PubMed] [Full Text HTML] [Full Text PDF]
[54] Ono T., Murakoshi M., Suzuki N., Iida N., Ohdera M., Iigo M., Yoshida T., Sugiyama K., Nishino H.: Potent anti-obesity effect of enteric-coated latoferrin: decrease in visceral fat accumulation in Japanese men and women with abdominal obesity after 8-week administration of enteric-coated lactoferrin tablets. Br. J. Nutr., 2010; 104: 1688-1695
[PubMed] [Full Text HTML] [Full Text PDF]
[55] Pilvi T.K., Harala S., Korpela R., Mervaala E.M.: Effects of high-calcium diets with different whey proteins on weight loss and weight regain in high-fat-fed C57BL/6J mice. Br. J. Nutr., 2009; 102: 337-341
[PubMed] [Full Text HTML] [Full Text PDF]
[56] Program prewencji i leczenia cukrzycy w Polsce na lata 2010-2011. Informacja prasowa Ministerstwa Zdrowia, Biuro Prasy i Promocji, Warszawa, 28 października 2011 (02.09.2012)
http://www.mz.gov.pl/wwwfiles/ma_struktura/docs/info_prasowa_cukrzyca_28102011.pdf
[57] Ruiz-Giménez P., Ibánez A., Salom J.B., Marcos J.F., López-Diez J.J., Vallés S., Torregrosa G., Alborch E., Manzanares P.: Antihypertensive properties of lactoferricin B-derived peptides. J. Agric. Food Chem., 2010; 58: 6721-6727
[PubMed]
[58] Sharma S., Singh T.P., Bhatia K.L.: Preparation and characterization of the N and C monoferric lobes of buffalo lactoferrin produced by proteolysis using proteinase K. J. Dairy Res., 1999; 66: 81-90
[PubMed]
[59] Shimizu H.: Development of an enteric-coated lactoferrin tablet and its application. Biometals, 2004; 17: 343-347
[PubMed]
[60] Suginohara Y., Miyazaki A., Hakamata H., Sakamoto Y., Ohta T., Matsuda I., Horiuchi S.: The heparin-bound fraction of human lipoprotein-deficient serum inhibits endocytic uptake of oxidized low density lipoprotein by macrophages. Atherosclerosis, 1996; 120: 167-179
[PubMed]
[61] Szopa M., Sieradzki J.: Zespół metaboliczny: kliniczno-teoretyczna dyskusja nad rozpoznaniem. Diabetologia Praktyczna, 2005; 6: 312-318
[Abstract] [Full Text PDF]
[62] Szopa M., Skupień J.: Omentyna – nowa adipokina w kontekście założeń nowej definicji zespołu metabolicznego (IDF 2005). Diabetologia Praktyczna, 2007; 8: 35-39
[Abstract] [Full Text PDF]
[63] Szostak W.B., Cybulska B.: Żywienie w hiperlipidemii. W: Żywienie człowieka zdrowego i chorego. Red. J. Hasik i J. Gawęcki. Wydawnictwo Naukowe PWN, Warszawa 2003; 136-153
[64] Takayama Y., Aoki R.: Roles of lactoferrin on skin wound healing. Biochem. Cell Biol., 2012; 90: 497-503
[PubMed]
[65] Takeuchi T., Kitagawa H., Harada E.: Evidence of lactoferrin transportation into blood circulation from intestine via lymphatic pathway in adult rats. Exp. Physiol., 2004; 89: 263-270
[PubMed] [Full Text HTML] [Full Text PDF]
[66] Takeuchi T., Shimizu H., Ando K., Harada E.: Bovine lactoferrin reduces plasma triacylglycerol and NEFA accompanied by decreased hepatic cholesterol and triacylglycerol contents in rodents. Br. J. Nutr., 2004; 91: 533-538
[PubMed] [Full Text HTML] [Full Text PDF]
[67] Tamano S., Sekine K., Takase M., Yamauchi K., Iigo M., Tsuda H.: Lack of chronic oral toxicity of chemopreventive bovine lactoferrin in F344/DuCrj rats. Asian Pac. J. Cancer Prev., 2008; 9: 313-316
[PubMed] [Full Text PDF]
[68] Thornalley P.J.: Cell activation by glycated proteins. AGE receptors, receptor recognition factors and functional classification of AGEs. Cell. Mol. Biol., 1998; 44: 1013-1023
[PubMed]
[69] Tripathy D., Mohanty P., Dhindsa S., Syed T., Ghanim H., Aljada A., Dandona P.: Elevation of free fatty acids induces inflammation and impairs vascular reactivity in healthy subjects. Diabetes, 2003; 52: 2882-2887
[PubMed] [Full Text HTML] [Full Text PDF]
[70] Vassiliou G., Benoist F., Lau P., Kavaslar G.N., McPherson R.: The low density lipoprotein receptor-related protein contributes to selective uptake of high density lipoprotein cholesteryl esters by SW872 liposarcoma cells and primary human adipocytes. J. Biol. Chem., 2001; 276: 48823-48830
[PubMed] [Full Text HTML] [Full Text PDF]
[71] Velliyagounder K., Kaplan J.B., Furgang D., Legarda D., Diamond G., Parkin R.E., Fine D.H.: One of two human lactoferrin variants exhibits increased antibacterial and transcriptional activation activities and is associated with localized juvenile periodontitis. Infect. Immun., 2003; 71: 6141-6147
[PubMed] [Full Text HTML] [Full Text PDF]
[72] Vogel H.J.: Lactoferrin, a bird’s eye view. Biochem. Cell Biol., 2012; 90: 233-244
[PubMed] [Full Text HTML] [Full Text PDF]
[73] Walrand S., Guillet C., Boirie Y., Vasson M.P.: In vivo evidences that insulin regulates human polymorphonuclear neutrophil functions. J. Leukoc. Biol., 2004; 76: 1104-1110
[PubMed] [Full Text HTML] [Full Text PDF]
[74] Willnow T.E., Goldstein J.L., Orth K., Brown M.S., Herz J.: Low density lipoprotein receptor-related protein and gp330 bind similar ligands, including plasminogen activator-inhibitor complexes and lactoferrin, an inhibitor of chylomicron remnant clearance. J. Biol. Chem., 1992; 267: 26172-26180
[PubMed] [Full Text PDF]
[75] Wu Y.M., Juo S.H., Ho Y.P., Ho K.Y., Yang Y.H., Tsai C.C.: Association between lactoferrin gene polymorphisms and aggressive periodontitis among Taiwanese patients. J. Periodontal Res., 2009; 44: 418-424
[PubMed]
[76] Yagi M., Suzuki N., Takayama T., Arisue M., Kodama T., Yoda Y., Numasaki H., Otsuka K., Ito K.: Lactoferrin suppress the adipogenic differentiation of MC3T3-G2/PA6 cells. J. Oral Sci., 2008; 50: 419-425
[PubMed]
[77] Zhang X., Beynen A.C.: Lowering effect of dietary milk-whey protein v. casein on plasma and liver cholesterol concentrations in rats. Br. J. Nutr., 1993; 70: 139-146
[PubMed] [Full Text PDF]
[78] Ziere G.J., Bijsterbosch M.K., van Berkel T.J.: Removal of 14 N-terminal amino acids of lactoferrin enhances its affinity for parenchymal liver cells and potentiates the inhibition of β-very low density lipoprotein binding. J. Biol. Chem., 1993; 268: 27069-27075
[PubMed] [Full Text PDF]
[79] Zimecki M., Kruzel M.L.: Milk-derived proteins and peptides of potential therapeutic and nutritive value. J. Exp. Ther. Oncol., 2007; 6: 89-106
[PubMed]
Autorka deklaruje brak potencjalnych konfliktów interesów.